小麦×玉米杂交双单倍体后代的HMW-GS组成和易位分析
2016-09-23庞建周王雪征茜晓哲王晨阳陈淑萍
庞建周,王雪征,茜晓哲,王晨阳,陈淑萍
(河北省农林科学院 旱作农业研究所,河北省农作物抗旱研究重点实验室,河北 衡水 053000)
小麦×玉米杂交双单倍体后代的HMW-GS组成和易位分析
庞建周,王雪征,茜晓哲,王晨阳,陈淑萍
(河北省农林科学院 旱作农业研究所,河北省农作物抗旱研究重点实验室,河北 衡水053000)
为了提高优质小麦的育种效率,在早代明确高分子量谷蛋白亚基和醇溶蛋白的组成,增加育种的针对性,以1个小麦杂交组合的296个DH株系群体为试材,采用半粒SDS-PAGE法检测麦谷蛋白,且用A-PAGE检测了醇溶蛋白。结果表明:母本衡09-6324的HMW-GS构成为null、7+9、2+12,是有Sec-1特征带的1BL/1RS易位系品种,父本师栾02-1亚基组成1、7+9、5+10,是非1BL/1RS易位系品种。296个DH株系共鉴定出6种不同的HMW-GS类型,均为父母本表达亚基类型的再分配,无变异类型。与面包加工品质呈正相关的5+10亚基出现频率为35.13%。DH株系材料中1BL/1RS易位系类型的占54.05%,1、5+10优质亚基同时出现,且没有Sec-1表达的占15.20%,这些株系可作为新的面包小麦种质资源使用。采用半粒SDS-PAGE和A-PAGE法对育种早代的优异单株进行有效鉴定和选择,可以提高优质小麦育种效率。
小麦;玉米;双单倍体;半粒电泳法;HMW-GS;黑麦碱
小麦和玉米杂交,利用玉米染色体消失诱导小麦双单倍体(DH,Double haploid)材料是一种高效率的小麦育种途径,缩短了育种年限,加快了育种进程[1]。由小麦×玉米杂交产生的是纯合的DH株系,因此DH群体可以稳定繁殖、长期使用,是一种永久性群体。DH材料的遗传结构能直接反映F1配子中基因的分离和重组,能在较短时间内构建一个理想的遗传作图群体,因而能有效地进行小麦遗传图谱的构建、QTL的定位以及多基因聚合等研究[2]。
20世纪70年代国外Barclay就利用“远缘杂交法”使得父本染色体消失从而诱导小麦单倍体的相关报道[3]。80年代中后期以英国、德国、法国、加拿大、CIMMYT为代表的学者在利用小麦×玉米杂交法诱导小麦单倍体领域的进展较快[4-6],应用此项技术在法国、加拿大等已经育成一批高产优质小麦品种[7]。我国单倍体育种起步较晚,中国农科院从20世纪90年代初就开展了利用小麦和玉米(或摩擦禾)杂交诱导小麦产生单倍体的研究,逐步完善了该技术过程,直到2011年才首次出现了应用此项技术育成小麦品种中麦533的报道[8]。
常规育种中品质性状不能够在田间通过性状表达和产量筛选,应用双单倍体育种与常规育种相结合的方法以及生化及分子标记辅助选择技术,实现优质育种从传统的经验到高效的精准育种的转化,是快速培育优质高产面包麦的有效途径。目前,关于地方常规品种的HMW-GS和1BL/1RS易位的遗传规律研究居多[9],但对于小麦×玉米杂交DH群体的研究鲜有报道。本试验将本地综合性状好的高产母本衡09-6324与强筋优质父本师栾02-1杂交,在F1授玉米粉诱导生成双单倍体植株,通过生化标记筛选含优质基因,但不表达黑麦碱(sec-1)的材料,作为小麦改良的新种质资源。利用染色体消失法获得的DH系在品种培育、材料创造、遗传研究等方面都有着重要意义。
1 材料和方法
1.1供试材料
所用小麦母本为衡09-6324,是本地综合性状好的高产优良品系,父本为师栾02-1,含1、5+10优质亚基,中国春为对照,均来自本单位小麦育种课题,玉米品种农大958系市售品种。杂种F1授玉米粉诱导DH小麦单倍胚,经0.75‰秋水仙素加倍处理后得到296个株系。双单倍体生产试验在河北省农林科学院旱作农业研究所旱作节水农业试验站温室进行,生化检测试验在河北省农作物抗旱研究重点实验室进行。
1.2试验方法
麦谷蛋白的提取和SDS-PAGE电泳参考封德顺[10]和赵庆玮等[11]的方法。采用SDS不连续缓冲系统,用北京六一公司的小型电泳槽进行凝胶电泳分析,分离胶浓度为10.5%(pH值8.8),交联度2.0%,浓缩胶浓度为3.7%(pH值6.8),交联度2.7%,稳压电泳,直至溴酚兰离开凝胶。
醇溶蛋白(Gliadin)的提取参考刘丽等[12]的方法,略有改动。采用酸性聚丙烯酰胺凝胶电泳法(A-PAGE)分析Gliadin,参考ISTA于1986年颁布的A-PAGE(pH值3.2,T=10%,C=3.5%)标准程序进行[13],检测Sec-1特征谱带,确定后代材料是否属于1BL/1RS易位系。
2 结果与分析
2.1亲本材料HMW-GS组成和1BL/1RS易位分析
衡09-6324是本单位小麦育种课题选育的抗旱、高产、早熟、抗病好的优良品系,SDS-PAGE分析表明,HMW-GS构成为null、7+9、2+12,A-PAGE分析表明有明显的Sec-1特征带,是典型的1BL/1RS易位系品种。师栾02-1是2007年国审的优质强筋小麦,产量中等,亚基组成1、7+9、5+10,是非1BL/1RS易位系品种,其电泳谱带的1、5+10属优质亚基,尤其是5+10亚基是当前世界公认的最优亚基[14],对小麦面粉烘烤品质有着重要的影响。本试验是通过双单倍体快速育种技术把优质的1和5+10亚基引入本地高产亲本衡09-6324,同时在早代进行生化标记Sec-1的鉴定,淘汰表达Sec-1的易位株系,选择的后代材料既继承父本的优质强筋特性,又具有母本的产量优势。中国春(null、7+8、2+12,非1BL/1RS易位)为对照材料。2.2DH后代群体中HMW-GS的类型和频率分布
以中国春和师栾02-1为对照,以衡09-6324/师栾02-1杂交F1种子为亲本,通过小麦×玉米杂交的双单倍体技术,诱导得到的296个DH纯合株系为试验材料,对籽粒的HMW-GS和Gliadin进行鉴定分析,亚基命名和分类参照魏益民等[15]方法。籽粒麦谷蛋白的SDS-PAGE分析表明,对应于小麦HMW-GS区域,所有DH籽粒都能检测到3~6条带,统计结果分别列于表1。图1为部分株系的HMW-GS组成类型。
由表1可以看出,296个株系在3个位点上共鉴定出6种不同的HMW-GS类型,均为父母本表达亚基类型的再分配,无变异类型。Glu-D1位点上与面包加工品质呈正相关的5+10亚基出现频率为35.13%(15.87+19.26,表1加下划线部分),与2+12亚基出现频率的数据进行统计学分析比较,χ2分析P值为0.033 5,分离达到显著水平。分析原因:1.可能是DH群体偏小;2.没有进行重复试验;3.胚诱导、加倍以及移栽过程中的死亡都可能导致结果出现偏差,有待进一步的试验验证。
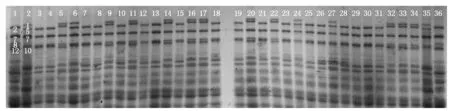
1、36.中国春;2、35.师栾02-1;3~34.F1(衡09-6324/02-1)诱导的部分DH株系。
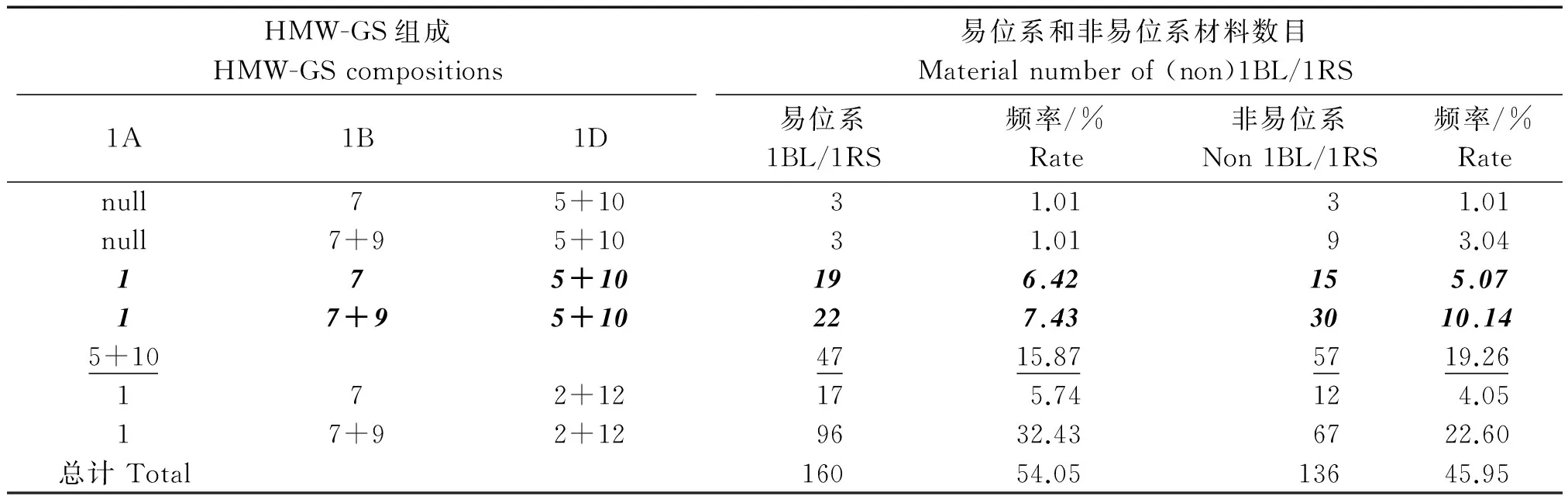
表1 部分DH株系的HMW-GS及1BL/1RS易位的组成类型
2.3DH后代群体中1BL/1RS易位的频率分布
以师栾02-1为对照,对DH株系的Gliadin进行A-PAGE分析,图2为部分株系Gliadin的类型,ω区Sec-1特征带被看作是黑麦1RS染色体臂的标记,是1BL/1RS易位的标志[16]。表1的数据进行统计,表达1和5+10亚基、且没有Sec-1的类型占15.20%(5.07+10.14,表1斜体加粗部分),这是我们育种需要的。这些材料可作为优质面包小麦新品种或育种材料使用,是栽培小麦改良的新种质资源。由图2可以看出,ω区Sec-1是4条特征谱带,而师栾02-1在此区域是3条不同的谱带,而由衡09-6324/师栾02-1诱导的DH材料中却出现了Sec-1特征谱带减弱和消失的情况,经重复验证,排除了试验错误。分析可能是玉米染色体作用的结果,此结果能否在其他育种试验中也能出现,以及玉米染色体在DH中是否完全消失,有待进一步试验研究。
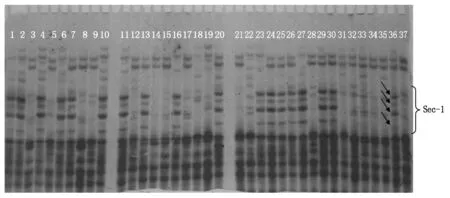
1~35.衡09-6324/02-1诱导的部分DH材料;36.衡09-6324;37.师栾02-1。
3 讨论
有关常规育种后代HMW-GS组成与1BL/1RS易位系的分离及组合规律研究较多[12,17-18],但利用染色体消失法选育DH过程中未见报道,本试验中尽管出现了5+10亚基的偏分离现象,在SDS-PAGE和A-PAGE电泳技术的协助下,也可选育出表达1和5+10亚基,不表达Sec-1的DH株系,这些可作为面包小麦新的种质资源或育种材料使用。
Zhou等[16]认为在选育强筋小麦品种时,不宜采用1BL/1RS易位品种(系)作亲本,基于本试验结果笔者认为采用小麦×玉米杂交技术可以选用单一亲本是1BL/1RS易位的材料,研究表明蛋白质含量与产量呈负相关[12,14],一般优质强筋小麦品种不高产。在DH生产过程中利用玉米染色体对Sec-1的抑制作用,通过生化标记在早代淘汰表达Sec-1的材料,若有幸能保留易位系增产抗病的优越性,使后代既继承亲本的强筋特点,又具有产量优势,这将进一步加快我国优质小麦育种的进程。在欧洲已经产生了Disponent、Branka等大量具有较好烘烤品质的易位系变种[7,19]。
国内外大量研究及实际育种实践表明,HMW-GS及1BL/1RS易位系是影响小麦烘烤品质的2个重要因素[15,20-21],本试验对小麦×玉米杂交诱导的296个DH株系的HMW-GS和1BL/1RS易位情况进行研究,出现一个值得关注的现象,母本衡09-6324是标准的易位系,有较好的高、低分子量谷蛋白亚基的遗传背景,父本师栾02-1是非易位系,其ω区也有醇溶蛋白的表达,但其杂交后代的DH株系却出现了ω区无蛋白表达的情况,类似结果未见报道。笔者认为此现象是DH诱导过程中玉米染色体作用的结果,虽然DH材料没有出现玉米染色体[1,22],但并不能否认杂交过程中玉米基因诱导DH的作用。本试验只是初步筛选了一个杂交组合,结果是否有普遍性值得进一步研究证实。假如DH株系能有效抑制Sec-1的表达,同时又能保留易位系的增产、抗病特性,那将为小麦品质改良开辟一条新的途径。
[1]陈新民,陈孝.小麦×玉米产生单倍体及双单倍体研究进展[J].麦类作物学报,1998,18(3):4-7.
[2]李春莲,薛芳,靳凤,等.等普通小麦DH群体条锈病抗性鉴定及抗病基因的分子检测[J].麦类作物学报,2013,33(6):1300-1304.
[3]Barclay I R.High frequencies of haploid production in wheat (TriticumaestivumL.) by chromosome elimination[J].Nature,1975,256:410-411.
[4]Zenkteler M,Nitzsche W.Wide hybridization experiments in cereals[J].Theor Appl Genet,1984,68:311-315.
[5]Laurie D A,Bennett M D.Wheat maize hybridization[J].Theor Appl Genet,1986,28:313-316.
[6]Suenaga K,Nakajima K.Efficient production of haploid wheat (Triticumaestivum) through crosses between Japanese wheat and maize (Zeamays)[J].Plant Cell Reports,1989,8(5):263-266.
[7]Depauw R M,Knox R E,Humphreys D G,et al.New breeding tools impact Canadian commercial farmer fields[J].Czech Journal of Genetics and Plant Breeding,2011,47(SI):S28-S34.
[8]陈新民,何中虎,刘春来,等.利用小麦×玉米诱导单倍体技术育成小麦新品种中麦533[J].麦类作物学报,2011,31(3):427-429.
[9]Chen X M,Wang F J,Li S M,et al.Stable production of wheat haploid and doubled haploid by wheat×maize cross[J].Acta Agronomica Siniac,2013,39(12):1-6.
[10]封德顺.体细胞杂种小麦与供体高冰草的高分子量麦谷蛋白亚基及其基因分析[D].济南:山东大学,2004:23-26.
[11]赵庆玮,王洪刚.国内外部分小麦种质资源高分子量麦谷蛋白亚基组成分析[J].山东农业科学,2007(6):25-28.
[12]刘丽,周阳,何中虎,等.高、低分子量麦谷蛋白亚基等位变异对小麦加工品质性状的影响[J].中国农业科学,2004,37(1):8-14.
[13]Cooke R J.The classification of wheat cultivars using a standard reference electrophoresis method[J].National Association of Colleges and Teachers of Agriculture,1987,17:273-281.
[14]Jazaeri S,Bock J E,Bagagli M P,et al.Structural modifications of gluten proteins in strong and weak wheat dough during mixing[J].Cereal Chemistry,2015,92(1): 105-113.
[15]魏益民,张波,关二旗,等.中国冬小麦品质改良研究进展[J].中国农业科学2013,46(20):4189-4196.
[16]Zhou Y,He Z H,Zhang G S,et al.Utilization of 1BL/1RS translocation in wheat breeding in China[J].Acta Agronomica Sinica,2004,30(6):531-535.
[17]张影全,张晓科,魏益民,等.高分子量麦谷蛋白亚基对小麦蛋白质品质特性的影响[J].西北农业学报,2013,22(1):48-53.
[18]何中虎,夏先春,罗晶,等.国际小麦育种研究趋势分析[J].麦类作物学报,2006,26(2):154-156.
[19]Ravi m,Chan S W.Haploid plants produced by centromere-mediated genome elimination[J].Nature,2010,464(7288):615-618.
[20]赵德辉,阎俊,黄玉莲,等.1BL/1RS易位对小麦贮藏蛋白组分含量和面团流变学特性的影响[J].作物学报,2015,41(11):1648-1656.
[21]魏学军,张娜,胡亚亚,等.53个小麦品种中1BL/1RS易位系的分子检测[J].分子植物育种,2013,11,(4):503-508.
[22]闫贵云,左静静,冯瑞云,等.玉米花粉诱导小麦单倍体技术研究进展[J].山西农业科学,2015,43(9):1204-1206.
Analysis of HMW-GS and Translocation Line in Progenies of Doubled Haploid from Wheat×Maize Cross
PANG Jianzhou,WANG Xuezheng,QIAN Xiaozhe,WANG Chenyang,CHEN Shuping
(Dry Land Farming Institute,Hebei Academy of Agriculture and Foresty Sciences,Key Lab of Crop Drought Tolerance Research of Hebei Province,Hengshui053000,China)
To improve the efficiency of high quality wheat breeding program,clarify the composition of HMW-GS and Gliadin in the early generation,296 lines from the wheat maize cross were analyzed useing SDS-PAGE method in this experiment,along with this Gliadin were analyzed useing A-PAGE method.A statistics showed that female parent Heng 09-6324 carried null,7+9,2+12 subunits,and were 1BL/1RS translocation lines with Sec-1 characteristic band.Male parent Shiluan 02-1 carried 1,7+9,5+10 subunits,and were non 1BL/1RS translocation.Six types of HMW-GS all originated from parents were identified from 296 DH lines.The frequency of subunits 5+10 which positively related to bread baking quality was 35.13%.A statistics showed that in the A-PAGE the type of translocation of the DH lines accounted for 54.05%,and the percent of lines with subunits 1 and 5+10,but non Sec-1 translocation were 15.20%,these lines could be used as new germplasm for quality breeding.The paper indicated that that the method of half-seed SDS-PAGE and A-PAGE which could be used to identify and select excellent plants in early generation breeding effectively.
Wheat;Maize;Doubled haploid;Half-seed electrophoresis;HMW-GS;Secalin
2016-03-26
河北省农林科学院前期预备项目(A2015040301);河北省农林科学院旱作农业研究所青年基金项目
庞建周(1983-),男,河北邯郸人,助理研究员,硕士,主要从事小麦×玉米杂交双单倍体育种研究。
S512.03;S513.03
A
1000-7091(2016)04-0126-04
10.7668/hbnxb.2016.04.021
