拟南芥CPK10/CPK30双突变体的构建及表型分析
2016-09-23魏凤菊
谢 鑫,孙 宁,魏凤菊
(河北农业大学 生命科学学院,河北省植物生理与分子病理学重点实验室,河北 保定 071001)
拟南芥CPK10/CPK30双突变体的构建及表型分析
谢鑫,孙宁,魏凤菊
(河北农业大学 生命科学学院,河北省植物生理与分子病理学重点实验室,河北 保定071001)
CDPKs是植物细胞内一类重要的钙感受器,拟南芥CPK10属于CDPK家族成员。为研究CPK10及与其同源性较高的CPK30是否共同参与逆境响应。首先构建了cpk10×cpk30双突变体,然后进行多种逆境下的生理表现检测,并利用RT-PCR方法分析2个基因的表达情况。结果显示,模拟干旱、盐、ABA处理拟南芥幼苗后,双突变体与野生型无差异;成苗期双突变体与单突变体对干旱敏感程度类似。转录水平检测到,在干旱胁迫时双突变体中RD29A的表达与野生型及单突变体趋势相反,呈下降趋势;响应ABA的OST1在双突变体中受干旱诱导后0.5 h明显表达上调。成功获得了cpk10×cpk30双突变体,由生理表型和表达分析结果推测CPK10与CPK30可能共同参与了依赖ABA的干旱逆境信号转导过程,且二者之间存在功能冗余。
钙依赖的蛋白激酶;双突变体;逆境胁迫;表型分析
钙依赖的蛋白激酶(Calcium-dependent protein kinases,CDPKs)作为生物体内重要的钙信号感受器,在胞内钙信号的级联传递过程中起着重要的作用。植物CDPK属于多基因家族,家族中许多成员在应答非生物逆境过程发挥重要功能。尤其在干旱、高盐胁迫方面最为明显,如拟南芥AtCPK10和AtCPK11可以受干旱和高盐诱导表达[1-2];AtCPK27[3]参与植株对盐胁迫的应答;过量表达AtCPK8[4]和AtCPK10[5]可以使植株耐旱力明显增强;其他植物中如玉米ZmCPK4[6]、葡萄VaCPK20[7]、姜ZoCDPK1[8]过量表达后,植株对干旱胁迫的耐受性明显增强;OsCPK12[9]正调控水稻应答高盐胁迫。有些CDPK成员表现对逆境的负调控,如AtCPK21[10]的突变体表现出对逆境胁迫耐受力增强。由于多基因家族中结构域的保守性,会存在多个基因的功能冗余现象,如AtCPK4和AtCPK11共同介导了ABA信号转导过程[11];AtCPK2和AtCPK20共同调控花粉管阴离子通道SLAH3[12]。烟草的NaCDPK4和NaCDPK5共同参与茉莉酸的早期合成过程,二者同时被沉默后,会导致茉莉酸的积累[13]。
前期研究发现,拟南芥CPK10参与对干旱胁迫的响应,通过ABA及钙离子介导了对气孔运动的调节[5]。在干旱处理条件下,Atcpk10突变体比野生型叶片失水萎蔫明显。拟南芥CDPK家族中,AtCPK30与AtCPK10属于同一亚家族,且序列相似性为家族最高,有研究显示AtCPK30参与了拟南芥幼苗期ABA等激素信号途径[14],而在应答逆境方面的研究未见报道。是否高度同源性的AtCPK30与AtCPK10基因存在相似或冗余的功能,这2个基因在同时缺失的情况下,对于逆境会发生怎样的表型变化,它们在参与逆境过程中的作用有什么相关性,这些问题都尚不清楚。本研究分别以CPK10基因及CPK30基因单突变体为父母本,通过单突变体杂交获得双突变体,结合生理表型及逆境相关基因检测,为深入探讨AtCPK10及AtCPK30在非生物逆境中的功能提供理论依据。
1 材料和方法
1.1试验材料
拟南芥cpk10(SALK_082441)及cpk30(SALK_087240)T-DNA插入突变体自SALK库购得。植物总RNA提取试剂、反转录酶、Taq聚合酶、dNTP等购自大连宝生物公司(TaKaRa);胶回收试剂盒等购自天根生物科技公司(Tiangen);质粒提取试剂盒购自北京全式金生物技术(TransGen Biotech)有限公司。植物激素ABA、MOPS等购自Sigma公司。其他化学药品为进口或国产分析纯试剂。
1.2植物材料培养方法
1.2.1植物材料培养首先用0.5%次氯酸钠消毒液(0.5%(V/V)NaClO+0.01%(V/V)Triton X-100)对拟南芥(Arabidopsisthaliana)种子进行消毒处理,4 ℃黑暗处理2~3 d,之后播种于MS培养基上生长约7 d。将幼苗移栽到土壤(蛭石∶营养土1∶1~1∶2)中培养(用透明塑料膜覆盖保湿4~5 d)。温室培养条件为:光/暗周期为16 h/8 h,温度22 ℃,湿度60%,光照强度80~100 μmol/(m2·s)。
1.2.2杂交试验采用常规的人工杂交方法获得双突变体。选取纯合单突变体作为亲本,杂交前去除母本已长角果和已开花以及未开花的花瓣和雄蕊,将父本的花粉涂布在母本未沾花粉的柱头上。授粉10 h后柱头毛萎蔫,3 d后角果形成并伸长,即为杂交成功。对杂交子代F1进行PCR检测,获得2个目的基因均杂合的个体,然后进行自交,分别从DNA和RNA水平对F2进行纯合鉴定,最终获得纯合体。
1.2.3逆境生理表型试验种子萌发阶段表型筛选:将各种基因型的种子播种在MS筛选培养基上。每种筛选浓度均进行3次重复试验。播种好的培养皿竖直放于温室中培养,定期交换摆放位置,排除位置效应可能对试验结果产生影响。培养14 d观察并照相。
幼苗阶段表型筛选:播种于MS培养基上的种子,竖直培养4 d。选取不同基因型长势相同的幼苗移栽到MS筛选培养基上。每种筛选浓度均进行3次重复试验。培养条件同上。生长14 d,观察照相。
成苗阶段表型筛选:温室培养钵中生长21 d左右的幼苗,进行干旱、高盐处理。定期交换培养钵位置,避免位置效应对试验结果造成影响,观察植株长势并照相。
1.3分子试验相关方法
1.3.1T-DNA插入检测参照CTAB法提取成熟叶片基因组DNA,T-DNA插入突变体的鉴定引物依据Signal网站(http://siganl.salk.edu/tdnaprimers.html)的方法设计。以基因组DNA为模板,不同引物组合进行PCR扩增,检测其T-DNA插入情况。
1.3.2总RNA的提取及RT-PCR利用TRIzol方法提取成熟叶片RNA,以反转录所得cDNA为模板扩增拟南芥组成型表达的持家基因EF1a,通过调整模板的加入量使得EF1a基因电泳检测时条带亮度基本一致。扩增目的基因,观察分析目的基因的亮度变化。
1.4引物设计与合成
PCR引物由上海生工通过PAGE纯化方式合成,各引物名称及序列如表1所示。
2 结果与分析
2.1F1cpk10×cpk30双突变杂合体的获得拟南芥CDPK家族中,CPK30与CPK10的序列相似性为86%。是否这二者共同参与了幼苗期植株响应逆境的过程,通过纯合单突变体杂交,构建cpk10×cpk30的双突变体,以cpk10×cpk30杂交F1各株基因组DNA为模板检测和筛选两基因的杂合体,利用CPK30TF/LBa1进行PCR扩增,只有cpk10×cpk30双突变杂合体能够扩增出条带。本试验随机选取约40棵F1植株进行检测,其中1,6,36,38号植株为cpk10×cpk30双突变杂合体,如图1所示。

表1 引物序列
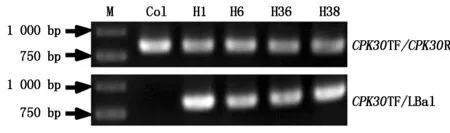
图1 双突变F 1杂合体鉴定
2.2F2双突变纯合体筛选
F1杂合体自交后,获得4个F2株系分别标记为:H1、H6、H36、H38。对这4个株系进行DNA水平鉴定以获得纯合的双突变体。首先检测CPK10基因的纯合情况。如果以CPK10TF和CPK10TR引物对进行PCR扩增,杂合体与野生型植株一样可以扩增出CPK10基因的片段;利用LBa1与CPK10TF组成引物对扩增,杂合体与纯合体植株均能扩增出条带。利用上述PCR方法对H1(24个植株样品)、H6(24个样品)、H36(6个样品)以及H38(18个样品)4个株系的材料分别进行了CPK10基因鉴定(图2-A)。结果显示,H1株系的10棵植株;H6株系的3棵植株;H36株系的4号植株;H38株系的5棵植株为cpk10纯合突变体(图2-A)。
经鉴定为CPK10基因插入纯合的植株再进行CPK30基因的插入突变鉴定。分别利用CPK30TF和CPK30TR、CPK30TF和LBa1 2对引物搭配进行PCR扩增。确定H1株系中1,3,4号植株;H38株系中5,7,13,15,17号植株;H36的4号植株为DNA水平上cpk10×cpk30双突变纯合体(图2-B)。
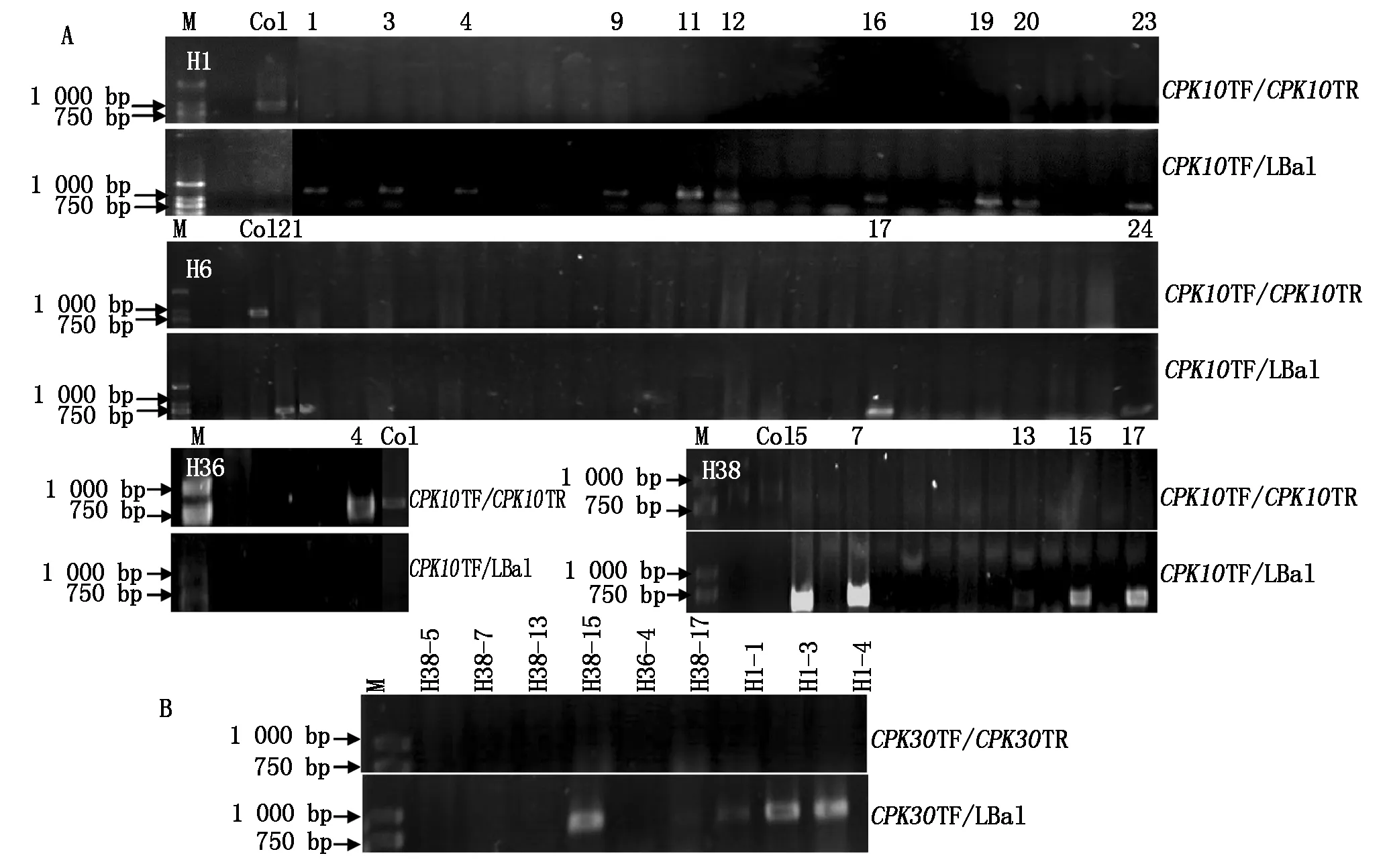
A.F2双突变纯合体CPK10基因纯合鉴定;B.F2双突变纯合体CPK30基因纯合鉴定。
2.3F3纯合双突变体的筛选
DNA水平上确定为CPK10和CPK30基因双突变的纯合体,接下来通过RT-PCR技术,在RNA水平检查2个基因的转录情况。野生型材料中能检查到特异扩增出的CPK10与CPK30基因片段(图3);cpk10单突变体中能检测到CPK30基因的片段,而没有扩增出CPK10基因的特异性条带;cpk30突变体材料中能够扩增出CPK10特异片段,而不能扩增出CPK30基因片段;双突变体材料中CPK10与CPK30 2个基因的目的片段都不能扩增出来。由以上结果得出,H1株系的4号植株为cpk10×cpk30的双突变纯合体。
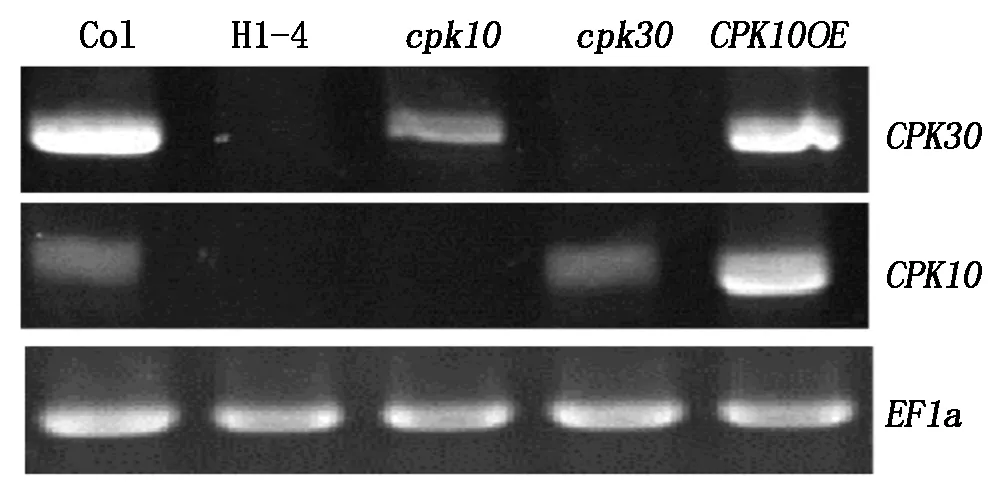
图3 RT-PCR鉴定双突变纯合体
2.4双突变的表型筛选
2.4.1萌发期表型观察在含有ABA的MS筛选培养基上,双突变体与单突变体均未见与野生型明显差异的表型。在MS培养基中加入Mannitol,观察各基因型在渗透物质处理条件下种子萌发情况,同样没有发现cpk10×cpk30双突变体与cpk10、cpk30植株及野生型有明显差异(图4)。将双突变体种子播种在含有100 mmol/L NaCl的MS筛选培养基上(图4),与正常培养基相比,双突变体、单突变体及野生型都表现为冠小发黄、根短;几种材料与野生型相比无明显差异。表型检测结果表明,在种子萌发阶段CPK10与CPK30可能不是拟南芥响应逆境胁迫过程中的关键基因。
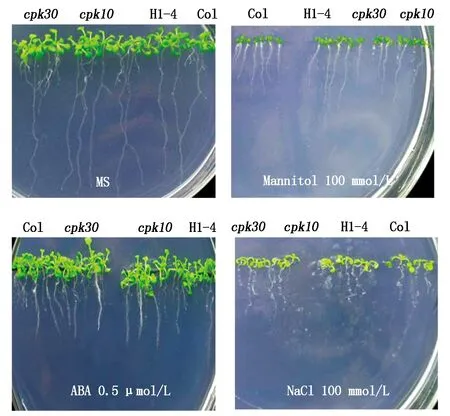
图4 双突变在处理培养基上种子萌发表型观察
2.4.2萌发后表型观察把MS培养基上长势相同的幼苗移到含有不同浓度的渗透物质(Mannitol)、ABA、NaCl的处理培养基上,双突变体、单突变体与野生型之间长势基本相同,无明显差异(图5)。这表明CPK10、CPK30基因的敲除,并没有使植株对以上逆境的应答发生改变,证明这2个基因不是拟南芥幼苗生长期响应上述逆境的关键因子。

图5 各基因型材料在处理培养基上表型观察
2.4.3成苗期表型观察正常生长14 d后的幼苗经干旱处理后,双突变体表现出与单突变体相似的干旱敏感表型:叶片萎蔫,茎秆倒伏(图6)。CPK10基因的缺失会导致植株耐旱能力减弱[11],双突变体也变现同样的表型,说明CPK30基因的缺失并没有使双突变体比cpk10对干旱更敏感。

图6 双突变体成苗期干旱胁迫表型观察
2.5干旱相关基因转录水平分析
为了探讨CPK10/CPK30是否共同参与了响应干旱胁迫的信号转导过程,本研究通过双突变体、单突变体及野生型植株进行干旱处理,利用RT-PCR技术对干旱逆境信号通路的关键基因表达情况进行了分析。对成苗莲座叶片进行0,0.5,1.0,3.0 h的干旱处理,RT-PCR结果显示,干旱胁迫响应的关键基因RD29A,随着时间的变化在野生型及单突变体中呈现明显的上升趋势,而在双突变体中呈现明显的下降趋势(图7),表明CPK10/CPK30基因与水分胁迫响应相关。OST1是依赖于ABA的干旱胁迫信号途径的关键基因。通过检测发现,双突变体中OST1基因在干旱处理0.5 h时表达量增加,而后又逐渐恢复到原来的水平;而在野生型与单突变体中OST1的表达量并没有明显变化(图7)。这预示着CPK10与CPK30基因可能共同参与了ABA依赖的干旱胁迫信号转导过程。那么,在该信号途径中这2个基因是否参与了对ABA合成的调控。接下来,选择了ABA合成关键基因ABA3和转录因子MYB2作为标记基因,结果显示这2个基因在干旱处理下的表达趋势在4种不同材料中并没有明显差异(图7),推测CPK10与CPK30可能在植株响应干旱胁迫过程中只参与ABA信号的转导,并未参与ABA的合成。
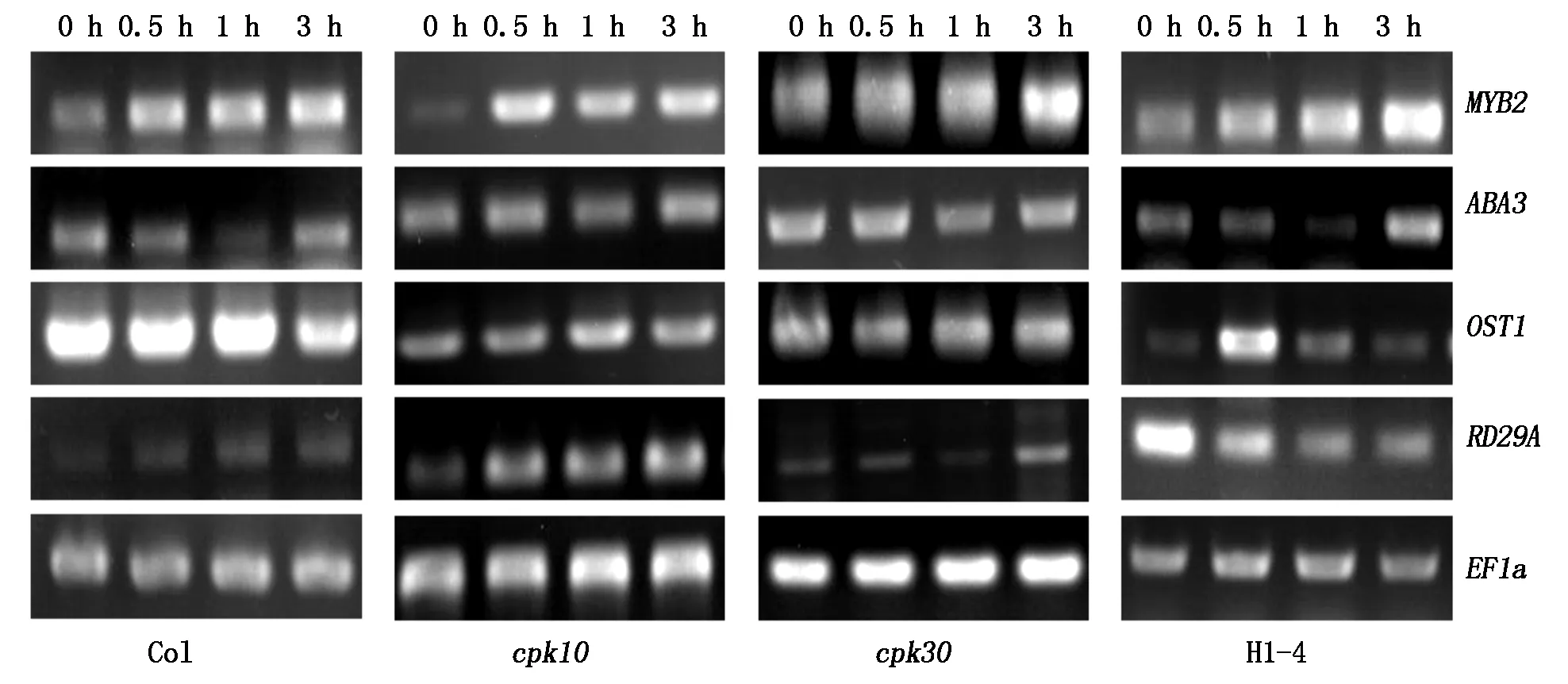
图7 干旱逆境标记基因的RT-PCR分析
3 讨论
对于多基因家族,一些成员表现为功能冗余,可使得植物对不良环境做出适应性调整。CDPKs家族成员在植物响应非生物逆境过程中发挥着重要作用,有些核心CDPKs对植物生存至关重要,序列同源性较高的CDPK基因,功能相似或冗余的可能性较大,如拟南芥CPK4、CPK5、CPK6和CPK11几个成员共同参与调控早期MAMP信号途径中基因的表达,4个成员同时缺失的突变体对flg22的抗性明显减弱[15]。马铃薯StCDPK4和StCDPK5共同参与对NADPH氧化酶的磷酸化,进而促使活性氧爆发[16]。AtCPK4、AtCPK11[11]、AtCPK10、AtCPK30和AtCPK32[17]都能通过转录因子ABF4调控植物对ABA的响应。AtCPK10与AtCPK30具有高度的序列相似性,本研究通过构建二者的T-DNA插入双突变体,进而分析探讨二者响应非生物逆境方面的功能。结果表明,幼苗期各种处理条件下双突变体与野生型无差异;成苗时期干旱处理条件下,双突变体与cpk10单突变体没有表型差异。由表型检测结果推测,在植株成苗期对干旱逆境的应答过程中,CPK10和CPK30二者并不存在功能冗余。接下来的转录水平检测显示,CPK30参与了对干旱逆境的应答,表型检测和转录水平检测结果的不一致性,暗示着在应答干旱逆境过程中,可能还有该家族的其他成员发挥功能。
CDPK属于钙受体激酶,逆境胁迫引发的钙信号经CDPK传递给下游哪些蛋白,这是探讨CDPK作用机制的关键所在。AtCPK10和AtCPK30是CDPK家族中最早报道的正向调控ABA信号的蛋白因子[18-19]。本研究结果显示,CPK10/CPK30在植株响应干旱胁迫过程中参与ABA信号的转导过程,这与前人研究结果一致。体外试验证明,AtCPK10、AtCPK30都能通过转录因子ABF4调控植物对ABA的响应,表明CDPK介导的ABA信号的转录调控可能是通过转录因子ABFs起作用。体内试验也证实了这一点:AtCPK32可以体内激活ABF4并启动下游基因的表达[17]。AtCPK4、AtCPK11可以调控ABF1和ABF4而影响逆境基因的表达,cpk4/cpk11双突变体表现出耐旱力减弱,双突变体中ABA响应的基因表达受到影响[11]。同样水稻的OsCPK21[20]过表达后,ABA响应基因会因ABFs活性变化而改变。CPK10[5]与CPK30[14]二者均定位于细胞膜,它们在体内是否会直接影响到细胞核中ABFs的活性,它们如何通过磷酸化作用将干旱逆境信号向下传递,这是我们将来进一步探讨的方向。
通过杂交获得cpk10×cpk30双突变体;表型检测得出,在种子萌发过程和幼苗时期CPK10和CPK30可能不是参与非生物逆境响应的关键蛋白;在植株成苗期二者参与干旱胁迫响应,并在依赖ABA的逆境响应信号途径中发挥功能。
[1]Qin L X,Nie X Y,Hu R,et al.Phosphorylation of serine residue modulates cotton Di19-1 and Di19-2 activities for responding to high salinity stress and abscisic acid signaling[J].Scientific Reports,2016,6:20371.
[2]Urao T,Katagiri T,Mizoguchi T,et al.Two genes that encode Ca2+dependent protein kinases are induced by drought and high-salt stresses inArabidopsisthaliana[J].Molecular & General Genetics,1994,244(4):331-340.
[3]Zhao R,Sun H,Zhao N,et al.The arabidopsis Ca2+dependent protein kinase CPK27 is required for plant response to salt-stress[J].Gene,2015,563(2):203-214.
[4]Zou J J,Li X D,Ratnasekera D,et al.Arabidopsiscalcium-dependent protein KINASE8 and CATALASE3 function in abscisic Acid-Mediated signaling and H2O2homeostasis in stomatal guard cells under drought stress[J].The Plant Cell,2015,27(5):1445-1460.
[5]Zou J J,Wei F J,Wang C,et al.Arabidopsis calcium-dependent protein kinase CPK10 functions in abscisic acid-and Ca2+-mediated stomatal regulation in response to drought stress[J].Plant Physiology,2010,154(3):1232-1243.
[6]Jiang S,Zhang D,Wang L,et al.A maize calcium-dependent protein kinase gene,ZmCPK4,positively regulated abscisic acid signaling and enhanced drought stress tolerance in transgenicArabidopsis[J].Plant Physiology and Biochemistry,2013,71:112-120.
[7]Dubrovina A S,Kiselev K V,Khristenko V S,et al.VaCPK20,a calcium-dependent protein kinase gene of wild grapevineVitisamurensisRupr.,mediates cold and drought stress tolerance[J].Journal of Plant Physiology,2015,185:1-12.
[8]Vivek P J,Tuteja N,Soniya E V.CDPK1 from ginger promotes salinity and drought stress tolerance without yield penalty by improving growth and photosynthesis inNicotianatabacum[J].PLoS One,2013,8(10):e76392.
[9]Asano T,Hayashi N,Kobayashi M,et al.A rice calcium-dependent protein kinase OsCPK12 oppositely modulates salt-stress tolerance and blast disease resistance[J].The Plant Journal:for Cell and Molecular Biology,2012,69(1):26-36.
[10]Franz S,Ehlert B,Liese A,et al.Calcium-dependent protein kinase CPK21 functions in abiotic stress response inArabidopsisthaliana[J].Molecular Plant,2011,4(1):83-96.
[11]Zhu S Y,Yu X C,Wang X J,et al.Two calcium-dependent protein kinases,CPK4 and CPK11,regulate abscisic acid signal transduction inArabidopsis[J].The Plant Cell,2007,19(10):3019-3036.
[12]Gutermuth T,Lassig R,Portes M T,et al.Pollen tube growth regulation by free anions depends on the interaction between the anion Channel SLAH3 and calcium-dependent protein kinases CPK2 and CPK20[J].The Plant Cell,2013,25(11):4525-4543.
[13]Hettenhausen C,Yang D H,Baldwin I T,et al.Calcium-dependent protein kinases,CDPK4 and CDPK5,affect early steps of jasmonic acid biosynthesis inNicotianatabacum[J].Plant Signaling & Behavior,2013,8(1):e22784.
[14]Yuan X,Deng K Q,Zhao X Y,et al.A calcium-dependent protein kinase is involved in plant hormone signal transduction inArabidopsis[J].Journal of Plant Physiology and Molecular Biology,2007,33(3):227-234.
[15]Boudsocq M,Willmann M R,Mccormack M,et al.Differential innate immune signalling via Ca2+sensor protein kinases[J].Nature,2010,464(7287):418-422.
[16]Witte C P,Keinath N,Dubiella U,et al.Tobacco calcium-dependent protein kinases are differentially phosphorylatedinvivoas part of a kinase cascade that regulates stress response[J].The Journal of Biological Chemistry,2010,285(13):9740-9748.
[17]Choi H I,Park H J,Park J H,et al.Arabidopsiscalcium-dependent protein kinase AtCPK32 interacts with ABF4,a transcriptional regulator of abscisic acid-responsive gene expression,and modulates its activity[J].Plant Physiology,2005,139(4):1750-1761.
[18]Simeunovic A,Mair A,Wurzinger B,et al.Know where your clients are:subcellular localization and targets of calcium-dependent protein kinases[J].Journal of Experimental Botany,2016,67(13):3855-3872.
[19]Sheen J.Ca2+-dependent protein kinases and stress signal transduction in plants[J].Science,1996,274(5294):1900-1902.
[20]Asano T,Hakata M,Nakamura H,et al.Functional characterisation of OsCPK21,a calcium-dependent protein kinase that confers salt tolerance in rice[J].Plant Molecular Biology,2011,75(1/2):179-191.
Construction and Phenotype Analysis of CPK10/CPK30 Double MutantinArabidopsis
XIE Xin,SUN Ning,WEI Fengju
(College of Life Sciences,Agricultural University of Hebei,Hebei Province Key Laboratory of Plant Physiology and Molecular Pathology,Baoding071001,China)
Calcium-dependent protein kinases (CDPKs) function as Ca2+sensor in various plant physiological processes.CPK10 was one member of CDPK family.In order to investigate the molecular mechanisms ofCPK10 and it′s high homologyCPK30 in response to stress,cpk10×cpk30 double mutant was firstly constructed.The phenotypes in response to various stress were detected,and the RT-PCR method was used to analyzed the two genes expression.The results showed that there was no difference in the seed germination stage between double mutant and wild-type plants in the treatment of drought,salt and ABA.In the mature stage,double mutant exhibited no difference to drought stress compared with wild-type and single mutant.Nevertheless,the expression levels ofRD29Awas decreased in the double mutant and showed opposite trend in wild-type and single mutant during drought stress.The expression of ABA-responsive geneOST1 was enhanced obviously in double mutant after drought stress 0.5 h.Thecpk10×cpk30 double mutant was successfully obtained.Our data indicated thatCPK10 andCPK30 might be involved in ABA-dependent signal transduction pathway,and showed redundant functions.
CDPK;Double mutant;Adversity stress;Phenotype analysis
2016-05-18
国家自然科学基金专项(31040052);国家自然科学基金青年基金项目(31101022);植物生理学与生物化学国家重点实验室开放课题项目(SKLPPBKF1504)
谢鑫(1990-),女,河北邯郸人,在读硕士,主要从事植物逆境研究。谢鑫、孙宁为同等贡献作者。
魏凤菊(1979-),女,河北衡水人,讲师,博士,主要从事植物抵抗逆境分子机制方面的研究。
Q78
A
1000-7091(2016)04-0094-06
10.7668/hbnxb.2016.04.016
