广西2013年-2015年鸡传染性支气管炎病毒S1基因分子流行病学分析
2016-09-21于冬玲谭春萍张艳雯
于冬玲,谭春萍,张艳雯
(南宁学院,广西南宁 530200)
广西2013年-2015年鸡传染性支气管炎病毒S1基因分子流行病学分析
于冬玲,谭春萍,张艳雯
(南宁学院,广西南宁 530200)
为了解近年来广西地区鸡传染性支气管炎病毒(IBV)的分子流行病学特点和遗传变异规律,本研究对广西地区2013年-2015年分离的26株IBV的S1基因进行了克隆、序列测定和分析。结果发现,26株IBV S1基因核苷酸序列长度在1 599 bp~1 632 bp之间,编码533个~544个氨基酸,说明分离毒株核苷酸存在较多的插入和缺失;分离毒株之间核苷酸序列同源性在76.5%~100%之间,其推导氨基酸序列同源性在74.4%~100%之间,分离株与参考株核苷酸及推导氨基酸的同源性分别介于75.9%~97.3%和74.9%~96.3%之间;分离毒株可以分为4个主要分支,南宁地区2013年-2014年分离毒株主要在第2分支,而在2015年分离毒株主要分布在第1.1分支,这是一个新的流行趋势,桂林地区的分离毒株主要分布在1.3、4.2分支,2015年以来的3个分离毒株分别存在于3和4.2分支,没有形成比较明显的优势流行毒株,玉林地区的毒株分布较散,也没有形成明显的优势流行毒株;分离毒株中大部分毒株与疫苗毒株同源性较低,部分毒株向着各自的方向进化,呈现出一定的地域性。
鸡传染性支气管炎病毒;S1基因;遗传进化
鸡传染性支气管炎(Avian infectious bronchitis,IB)是由鸡传染性支气管炎病毒(Avian infectious bronchitis virus,IBV)引起的危害养鸡业的重要传染病,主要侵害鸡的呼吸系统和泌尿生殖系统,从而引起死淘率上升、产蛋下降、继发感染等。近年来,IB的防控虽取得了一定的成效,但是部分地区仍有发生,特别是肾型IB尤为严重,给我国养鸡业造成了重大损失[1-4]。本研究对从广西地区检测的26份IBV进行了S1基因的克隆、测序及分子流行病学分析,及时了解IBV变异情况,为IB的流行病学研究、疫苗筛选和综合防控奠定基础。
1 材料与方法
1.1材料
1.1.1毒株本研究所选取得毒株均分离自广西南宁、桂林、玉林等地区的散养鸡场,具体信息如表1所示。选用某商品H120疫苗作为阳性对照,选用健康鸡样品作为阴性对照。
1.1.2主要试剂DH5α感受态细胞,天根生化科技有限公司产品;AxyPrep体液病毒DNA/RNA小量试剂盒、DNA片段快速纯化/回收试剂盒,爱思进生物技术(杭州)有限公司产品;一步法RT-PCR试剂盒(PrimeScript One Step RT-PCR Kit Ver.2)、ExTaqDNA聚合酶(5 U/μL)、DNA Marker DL 2 000、pMD18-T Simple载体,宝生物工程(大连)有限公司产品。
1.2方法
1.2.1引物设计与合成应用Primer 5.0 和DNA Star Lasergene 7.1基因分析软件分析了GenBank中注册的IBV S1基因序列,设计扩增S1全长片段的引物:
P1 (Forward primer):5′-AAGACTGAACAAAAGACCGACT-3′(22 bp);
P2 (Reverse primer ):5′-CAAAACCTGCCATAACTAACATA-3′(23 bp);
引物由上海生工生物工程技术服务有限公司合成。P1/P2引物对扩增的片段长度为1600 bp左右[5]。
1.2.2S1基因的克隆测序IBV RNA的提取按照AxyPrep体液病毒DNA/RNA小量试剂盒说明书进行。参照PrimeScript One Step RT-PCR Kit Ver.2 一步法反转录试剂盒说明进行RT-PCR,反应采用25 μL体系,反应条件为:50 ℃反转录30 min;94 ℃ 5 min;94 ℃ 40 s,53 ℃ 40 s,72 ℃ 1 min,共30个循环;72 ℃ 10 min,4 ℃ 终止反应。扩增产物使用10 g/L琼脂糖凝胶电泳检测,并用成像系统照相。S1基因纯化使用爱思进DNA片段快速纯化/回收试剂盒进行。纯化后产物与pMD18-T Simple载体进行连接反应,按说明书进行。将连接产物转化至DH5α感受态细胞,涂布于含氨苄青霉素的LB固体培养基上,37 ℃培养过夜,挑选单菌落扩大培养,提取质粒,鉴定克隆成功样品,送上海生工生物工程技术服务有限公司测序。
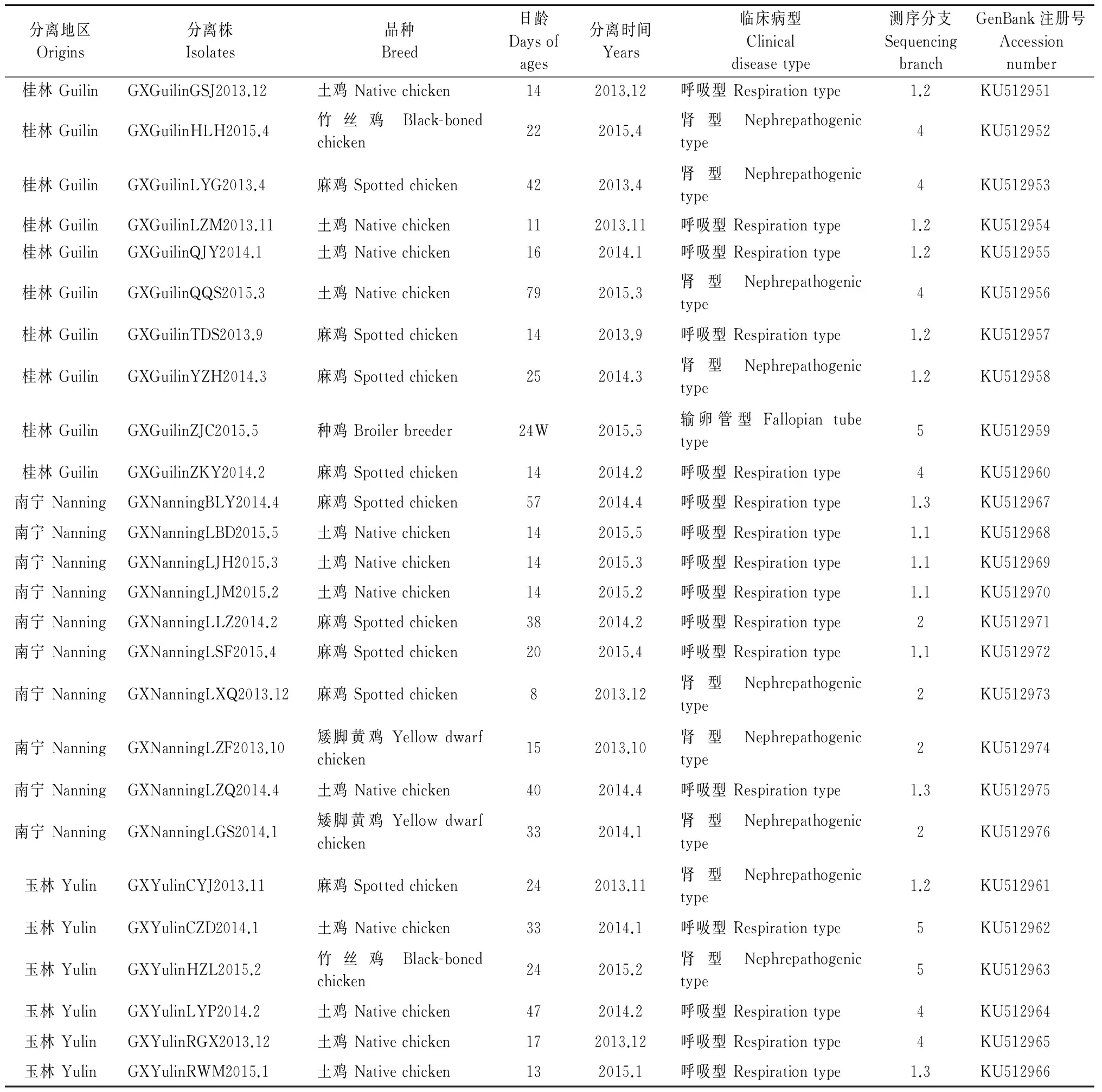
表1 IBV广西分离株的来源及背景
1.2.3序列分析与进化树的构建用DNA Star对分离株与GenBank中登录的12株参考毒株S1基因核苷酸序列及其推导氨基酸序列进行同源性比较分析。应用ClustalX计算方法进行序列比对,使用MEGA5.0的临位相接法(Neighbor-Joining法)进行进化树的构建,并用Bootstrap对进化树进行统计验证。参考毒株及其GenBank登录号如下:M41(DQ834384)、Ark99(M99482)、4/91(AF093794)、H120(M21970)、Ma5(AY561713)、W93(AY427818)、A2(AY043312)、QXIBV (AF193423)、LX4 (AY189157)、PSH050513(DQ160004)、LDT3(AY702975)和28/86(AY846750)。
2 结果
2.1RT-PCR扩增结果
采用RT-PCR扩增IBV S1基因片段,结果分别扩增出约1 600 bp的目的条带,与预期的片段大小相符。如图1所示为部分样品S1基因RT-PCR扩增片段产物的电泳结果。
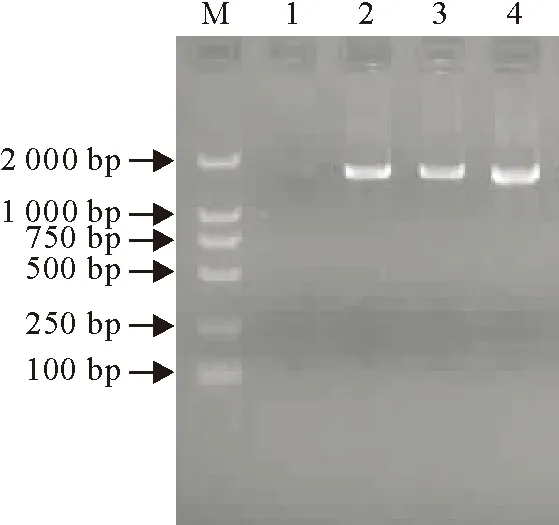
M.DNA标准DL 2 000;1.阴性对照;2 .阳性对照;3~4.样品
M.DNA Marker DL 2 000; 1 Negative control; 2 Positive control; 3-4.Samples
图1IBV S1基因RT-PCR扩增片段产物的电泳结果
Fig.1The electrophoresis results of RT-PCR products of IBV S1 gene
2.2测序结果
将测序所得序列提交GenBank,获得分离株的注册号,如表1所示。
2.3同源性分析
测序结果表明,26株IBV S1基因核苷酸序列长度在1 599 bp~1 632 bp之间,编码533-544氨基酸,说明分离毒株存在较多的碱基插入和缺失。26个分离毒株与12个参考毒株S1基因序列通过MegAlign软件Clustal W 方法进行序列比较[5-6]。分析表明,各分离毒株之间核苷酸序列同源性在76.5%~100%之间,其推导氨基酸序列同源性在74.4%~100%之间。分离株与参考株核苷酸及推导氨基酸的同源性分别介于75.9%~97.3%和74.9%~96.3%之间。可见分离毒株之间、分离毒株与参考毒株之间同源性差异较大,参差不齐,十分复杂。推测S1基因的多样性差异是导致目前免疫失败的一个重要因素。
2.4遗传进化分析
将26株IBV分离株与参考毒株的S1基因核苷酸序列用MEGA5.0分析软件构建进化树,进行遗传分析,如图2所示。结果表明,全部分离毒株可以分为4个大分支,每个分支里面又可以分为若干小的分支。第1分支包括13个分离毒株(56%)和3个参考毒株(LX4、QXIBV、A2);第2分支包括4个分离毒株(15%)和1个参考毒株(PSH050513)1个疫苗毒株(LDT3)。第3分支包括2个参考毒株(W93、M41)3个疫苗毒株(28/86、H120、Ma5);第4分支包括6个分离毒株(23%),未见参考毒株,由于本研究选用的参考毒株有限,说明这个分支是一个新的进化方向,与选用的经典毒株均不同;第5分支包括3个分离毒株(12%)和1个参考毒株(Ark99)1个疫苗毒株(4/91)。结果说明,分离毒株与疫苗毒株均不在同一分支上,1、2分支毒株与我国经典的分离毒株在同一分支,未见3分支分离毒株,5分支与国外Ark99在同一分支,说明国外毒株也存在于广西地区,并产生了进化,4分支与选用的参考毒株均不在一起,属不同的分支,提示为一个新的进化方向。
从分布规律来看,2015年以来南宁地区4个分离株均在1.1分支;2013年10月~2014年4月南宁6个分离株中有2个分离毒株在1.3分支、4个分离毒株在2分支;2013年4月~2015年5月桂林地区10个分离株中有5个分离毒株在1.2分支、4个分离毒株在第4分支、1个分离毒株在5分支。桂林地区的毒株分布较散,提示桂林地区的IBV感染更为复杂。2013年11月~2015年2月玉林地区6个分离株中有2个分离毒株在1分支、2个分离毒株在第4分支、2个分离毒株在5分支,分布也较分散。
综合来看南宁地区2013年-2014年分离毒株多在第2分支,而在2015年分离毒株多分布在第1.1分支,这是一个新的流行趋势。桂林地区的分离毒株多分布在1.3、4.2分支,2015年以来的3个分离毒株分别存在于3、4.2分支,没有形成比较明显的优势流行毒株。玉林地区的毒株分布较分散,也没有形成明显的优势流行毒株,这可能与其地理位置有关。
3 讨论
广西地区是我国重要的黄羽肉鸡养殖区域,其中又以桂林、玉林、南宁3个地区存栏量最多。鸡传染性支气管炎是危害养殖生产的重要疾病之一,给养殖场造成了重大损失[7-8]。检测结果表明,养殖生产的各日龄段均有分离到IBV,其中10日龄~30日龄为高发阶段,有16群,占21%,最早8日龄就有感染发病的。这可能是因为雏鸡在这个生长阶段,IBV母源抗体已经逐步消退,但疫苗免疫后保护力尚未完全建起,IBV野毒感染率比较高,所以10 d~ 30 d发病较多。呼吸型IB多发生于20日龄以内,有16群,占62%。另外从4群无发病症状的鸡群也分离到了IBV,证明IB是存在潜伏感染或者耐过后持续排毒的。发生大肠埃希菌、沙门菌、支原体等感染的鸡群也能够检出IBV,说明存在混合感染。IBV分离毒株中,大部分毒株与疫苗毒株同源性较低,部分毒株向着各自的方向进化,呈现出一定的地域性。这与国内其他学者的研究结果相近[9-10]。
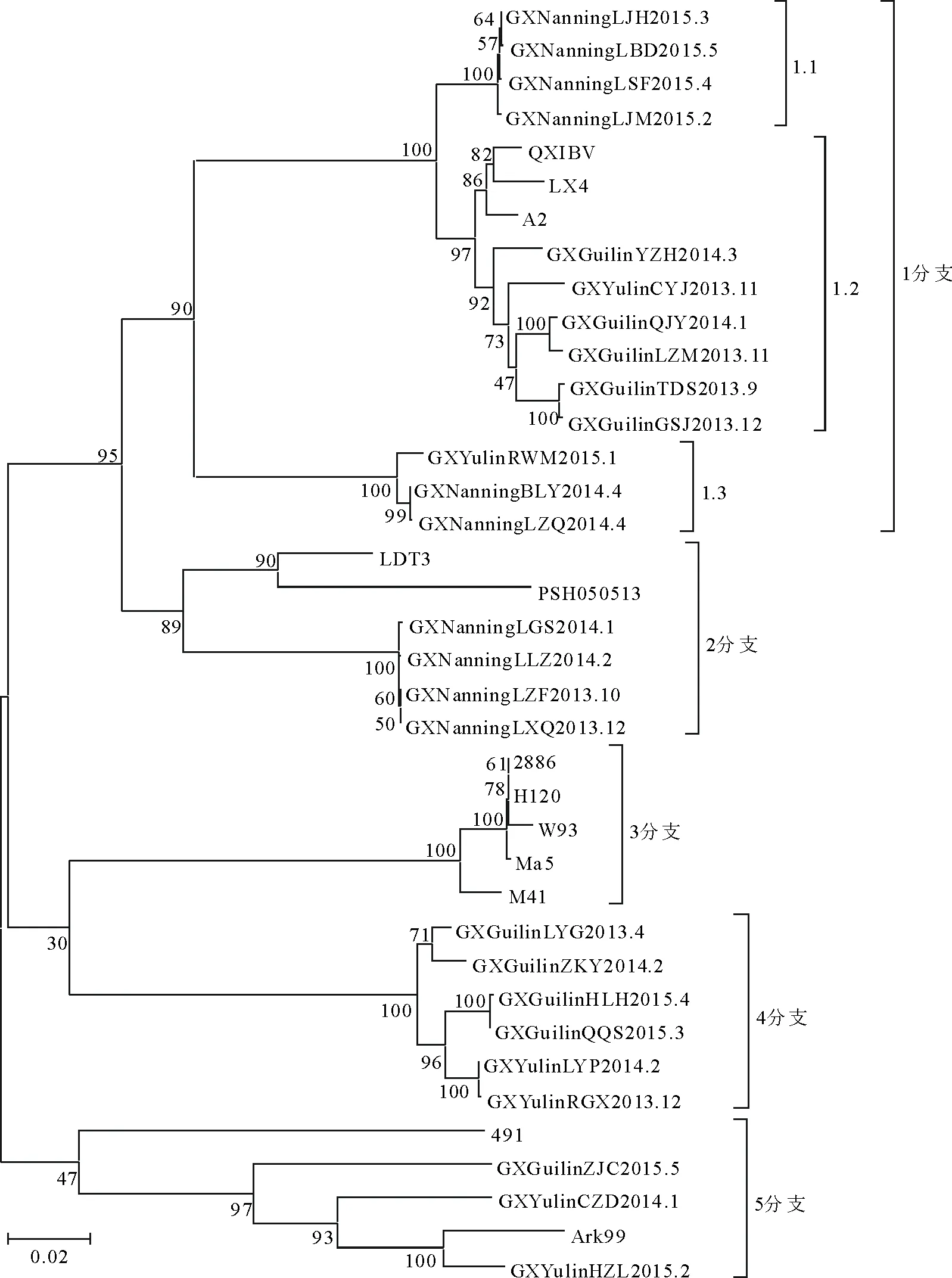
图2 S1基因进化树
IB的早期免疫主要是黏膜免疫,雏鸡母源抗体虽高,但始终不能产生有效的保护。针对广西地区IB的复杂情况,在免疫预防方面笔者认为,提早接种免疫,使雏鸡产生黏膜免疫力,是非常重要的,可以选择1日龄喷雾(或者3天龄点眼)免疫多价弱毒疫苗。从测序结果来看,可以选用H120、4/91、LDT3等弱毒苗来免疫。也可以根据目前流行趋势使用油苗免疫,所使用毒株最好在第1、2、4分支当中选择2个~3个毒株,制成多价苗,以期能够取得较好的免疫效果。
IBV毒株变异较快,一方面是病毒本身不断变化,另一方面是免疫压力促使病毒免疫逃避,产生新的流行毒株,所以需要我们不断的持续跟踪IBV的变化,适时掌握其流行规律,制定合理的防疫计划,对于养殖生产具有重要的指导意义。
[1]Cavanagh D,Naqi S A.Infectious bronchitis in diseases of poultry,11th[M].2003:108-130.
[2]Fabricant J.The early history of infectious bronchitis [J].Avian Dis,2000(42):648-650.
[3]Cavanagh D,Casais R,Armesto M,et al.Manipulation of the infectious bronchitis coronavirus genome for vaccine development and analysis of the accessory proteins [J].Vaccine, 2007, 25(30):5558-5562.
[4]谢景伟,陈峰,谢青梅,等.鸡传染性支气管炎病毒分子流行病学研究进展[J].动物医学进展,2008,29(8):75-78.
[5]Li L,Xue C,Chen F,et al.Isolation and genetic analysis revealed no predominant new strains of avian infectious bronchitis virus circulating in South China during 2004-2008 [J].Vet Microbiol,2010,143(2-4):145-154.
[6]Ji Jun,Xie Jingwei,Chen Feng,et al.Phylogenetic distribution and predominant genotype of the avian infectious bronchitis virus in China during 2008-2009 [J].Virol J,2011(8):184.
[7]磨美兰,李孟,何坤,等.近期广西鸡传染性支气管炎流行概况及防控措施[J].广西畜牧兽医,2013,29(2):67-70.
[8]孙裴,施雷,魏建忠,等.鸡传染性支气管炎病毒感染的分子机制[J].动物医学进展,2013,34(12):199-203.
[9]丁梦,黄磊,单虎.IBV分离株部S1蛋白基因的序列分析[J].动物医学进展.2008,29(5):36-39.
[10]磨美兰,李孟,韦正吉,等.广西1985年-2008年IBV分离株S1基因高变区Ⅰ和N基因的序列和系统进化分析[J].中国预防兽医学报,2009,31(10):761-765.
Molecular Epidemiological Analysis of S1 Gene of Avian Infectious Bronchitis Virus Isolated in Guangxi Province during 2013-2015
YU Dong-ling,TAN Chun-ping,ZHANG Yan-wen
(Nanning University,Nanning,Guangxi,530200,China)
In order to find out the characteristics of molecular epidemiology and molecular genetic variation of avian infectious bronchitis virus (IBV) in Guangxi province of China in recent years,S1 genes were cloned,sequenced and analysed in 26 IBV strains isolated between 2013 and 2015 in the study.The results showed that: 26 IBV strains of S1 gene nucleotide sequence lengths were 1 599 bp-1 632 bp,encoding 533-544 amino acids,indicated that there were many insertions and deletions.The nucleotide sequence homology of strains isolated was 76.5%-100%,the amino acid sequence homology was 74.4%-100%,the nucleotide and amino acid homology between the isolated and referenced strains were 75.9%-97.3% and 97.3%-74.9% respectively.Isolated strains can be divided into four main branches.In Nanning area,some strains were in the No 1 branch from 2013 to 2014,but the other strains were mainly distributed in the No 1.1 branch in 2015,it is a new popular trend.In Guilin area,strains were mainly distributed in No 1.3 and 4.2 branches,but some strains were found in No 3 and 4.2 branches in 2015,there was no obvious predominant pandemic strain.The strains isolated were very fragmented distribution in Yulin area,also does not have the obvious predominant pandemic strain.The homology of most isolates and vaccine strains was low,and some of the strains showed some regional characteristics toward the respective directions in the evolution.
Avian infectious bronchitis virus; S1 gene; genetic evolution
2015-11-06
南宁学院校级项目(2014JSGC04,2014XJ09)
于冬玲(1985-),女,山东威海人,讲师,硕士研究生,主要从事动物疾病研究。
S852.659.6;S858.31
A
1007-5038(2016)08-0019-05
