秦岭南坡中段主要森林群落类型划分及环境梯度解释
2016-09-19王宇超周亚福王得祥陕西省西安植物园陕西西安7006西北农林科技大学林学院陕西杨凌7200
王宇超,周亚福,王得祥. 陕西省西安植物园,陕西 西安 7006;2. 西北农林科技大学林学院,陕西 杨凌 7200
秦岭南坡中段主要森林群落类型划分及环境梯度解释
王宇超1,周亚福1,王得祥2*
1. 陕西省西安植物园,陕西 西安 710061;2. 西北农林科技大学林学院,陕西 杨凌 712100
该研究旨在揭示秦岭大熊猫主要栖息地植物群落分布规律及其与环境的对应关系。在研究区设置2条样带,每条样带各设置40~50个样地,共计93个。通过样地调查,利用双向聚类法(Two-way Clustering Method)、DCA、DCCA对秦岭大熊猫(Ailuropoda melanoleuca)栖息地森林群落进行数量分类和排序。(1)TWINSPAN将研究区93个样地分成太白红杉(Larix chinensis)、巴山冷杉(Abies fargesii)、铁杉(Tsuga chinensis)、华山松(Pinus armandii)、油松(Pinus tabulaeformi)、锐齿槲栎(Quercus aliena var. acuteserrata)等34个群落类型,并通过去趋势对应分析(DCA)对93个样地进行排序,结果表明不同类型样地呈现聚集分布,充分验证了TWINSPAN对群落类型划分的科学性。(2)通过对93个样地除趋势典范对应分析(DCCA)和Monte Carlo显著性检验表明,在分析12个环境因子中,制约秦岭南坡大熊猫栖息地群落类型、植物种分布格局的主要因素是海拔、坡度、速效磷与全磷。海拔、坡度、速效磷和全磷能够很好地解释群落样地及优势种在环境梯度上的分布,同时表现出样方间在种类和环境因子组成上的相似性。
秦岭;南坡;双向聚类法;去趋势对应分析;除趋势典范对应分析
引用格式:王宇超, 周亚福, 王得祥. 秦岭南坡中段主要森林群落类型划分及环境梯度解释[J]. 生态环境学报, 2016, 25(6): 965-972.
WANG Yuchao, ZHOU Yafu, WANG Dexiang. The Quantitative Classification and Environmental Interpretation of Forest Communities in the Middle Area of South Slope of Qinling Mountains [J]. Ecology and Environmental Sciences, 2016, 25(6): 965-972.
目前,国内针对山地植被进行的研究,无论是从宏观尺度还是微观研究,都开展得比较深入和透彻。其中,山地植物群落格局与环境关系研究是当前植被研究的热点问题之一。许多研究认为植被分布地形环境(徐广平等,2005;张先平等,2006)、群落的演替阶段(王世雄等,2010)、外界干扰程度(李晋鹏等,2007)、地表和植被状况(王国宏等,2001;张玲等,2007)、土壤状况(龚志莲等,2015)等因素都可能对植物群落分布格局产生影响。任何植物群落的存在,都与其所在的环境条件有着密切的相关性。因此,清楚认识植物群落分布格局和生态环境之间的关系是植被生态研究的基础与前提。然而,数量分析为客观、准确地群落类型划分及其与环境之间的生态关系提供了科学、合理、有效的途径,己成为植被生态学研究的重要内容之一(张新时,1991;江洪等,1994)。其中,数量分类和排序是最主要和应用最广泛的多元统计方法,是全面认识研究区域内植被分布格局特征的重要手段(张峰等,2003)。目前,数量分类和排序已被较为广泛地应用在森林(Veazquez,1994;王晶等,2016)、湿地(李思阳等,2016)、草甸(宋爱云等,2006)、灌丛(孙菊等,2009)等群落类型研究之中。数量分类和排序已经在植被生态学中发展成为研究群落生态关系的重要数量方法(Russelt,1991;Pamer,1993)。
佛坪和长青国家级自然保护区位于秦岭中段南坡,是秦岭重要核心区域,同时也是大熊猫Ailuropoda melanoleuca等珍稀动物的主要栖息地。其植被空间格局直接影响到大熊猫等野生动物生存空间,所以对该研究区植物群落进行研究显得尤为重要。然而,近些年关于该研究区植被分布格局及其与环境关系的研究还较少,对植被分布格局及其与环境关系的认识还不是很清楚。本研究结合我国山地植物群落数量分类和排序的相关研究,利用TWINSPAN数量分类和DCA、DCCA排序,全面深入探讨秦岭大熊猫栖息地地形、土壤等环境因子与山地植被类型及其分布的关系,并对植被类型作出环境解释,以期为保护栖息地的森林植被,充分发挥其强大的生态系统服务功能,实现保护区森林生态系统可持续发展和管理提供理论指导,为制定保护措施提供科学依据。
1 研究区概况与研究方法
1.1研究区概况
佛坪、长青国家级自然保护区位于秦岭中段南坡,地理位置为东经107°19′~107°55′、北纬33°17′~33°46′,总面积59146 hm2,森林覆盖率达90%以上。研究区为亚热带向暖温带的过渡区,海拔在900~3071 m之间,年均温为11.4~15 ℃,无霜期为225~239 d,年降水量813~1129.6 mm。研究区主要以保护大熊猫、朱鹮 Nipponia nippon、羚牛Budorcas taxicolor、金丝猴 Rhinopithecus roxellana为主。该区植被类型丰富,主要有针叶林、阔叶林、竹林、草甸等植被类型。主要分布植物群落优势种有太白红杉Larix chinensis、巴山冷杉Abies fargesii、华山松Pinus armandii、油松Pinus tabulaeformis、锐齿槲栎 Quercus aliena var. acutiserrata、巴山木竹Bashania fargesii、秦岭箭竹Fargesia qinlingensis等。
1.2研究方法
1.2.1样地设置
在查阅相关资料和踏查的基础上,采用典型取样法对研究区内海拔900~3071 m之间的山地乔木植物群落进行野外调查。在每个保护区内各设置 1条样带,在每条样带上按海拔每升高 100 m设置1~4个典型样地。2条样带共设置93个样地。
乔木样地:面积500 m2(20 m×25 m),对样地内乔木层(胸径大于4 cm)植株进行每木检尺,在每个样地内设置5个5 m×5 m的灌木样方和1 m×1 m的草本样方,并在每个样地沿对角线设置25个1 m×1 m灌草频度调查样方。所有样地调查记录内容包括植物种类、高度、盖度、胸径、频度、生活力等指标,同时记录各群落的综合特征和生境特征,主要包括:经纬度、海拔、群落盖度、层间植物种类、坡位、坡度、坡向和腐殖质层厚度等。
1.2.2土壤采集及测定
在每个样地四角及中央设置5个土壤剖面,每个剖面分3层取土。分别是0~20、20~40、40~60 cm,带回室内分析其养分含量。测试指标有:全氮、全磷、全钾、有效氮、有效磷、有效钾、有机质、pH值。土样处理以及测试方法参照鲍士旦等(1999)实验方法进行。
1.2.3数据分析
(1)物种重要值计算(王宇超等,2012)
乔木重要值=(相对高度+相对密度+相对优势度)/300;
灌木及草本重要值=(相对密度+相对频度+相对盖度)/300;
(2)植物群落数量分类与排序
本文采用双向聚类法TWINSPAN(Two-Way Indicators Species Analysis)对群落进行分类,去趋势对应分析 DCA(Detrended Correspondence Analysis)、除趋势典范对应分析(Detrended Canonical Correspondence Analysis,DCCA)进行排序。
1.2.4坡度、坡向的标准化


野外测量时,坡度以度为单位记录(0°~90°)。坡向的数据用方位角表示(0°~360°)。为了便于分析,将坡度和坡向通过如下公式进行标准化:式中,SP为百分比坡度,S为坡度,SP的取值范围为(0, 200);AT为标准化坡向,A为坡向,AT的取值范围为(0, 2),最大值2代表湿冷的东北坡,最小值0则代表干热的西南坡。
1.2.5数据统计与分析
数据分析利用TWINSPAN 2.3、Canoco 4.5、Excel 2007等软件。
2 结果分析
2.1TWINSPAN数量分类
基于样地调查数据生成的物种重要值矩阵,利用双向聚类法(Two-way Clustering Method)对93个样地进行数量分类,该方法采用TWINSPAN软件包进行。本文根据物种重要值选用的假种分级为7级:0.02~0.05,0.05~0.1,0.1~0.15,0.15~0.2,0.2~0.35,0.35~0.7,>0.7。列表中的最大物种数为120个,用来划分每一组样地个数最小值为 5,最大划分分级水平为 9,每次划分的最多区别中数目为8。样方分类时为了突出优势种的作用,对7级“假种”的权重分别取:1、1、1、1、2、3和4。
93个样地 658种植物进行数量分类结果见图1。其中,图 1的下方和右方分别显示对样方和物种的分化水平和类型。从图1可以看出TWINSPAN 将93个样地划分成39个群落类型。由于本研究只是从植物群系方面进行划分,所以结合野外调查数据,对一些划分过细的样地进行合并。样地46、65、17、18、19、25的建群种均为锐齿槲栎,将其合并成为一个群系;样地12、15、24、69、45、58、51的建群种均为油松,将其合并成一个群系,样地59、10、11、26、28、29、53建群种为板栗,将其合并成一个群系。通过合并将 93个样地划分成太白红杉、巴山冷杉、油松、华山松等 34个群系,具体名录如表1所示。

表1 秦岭南坡中段主要乔木群落类型Table 1 Tree community types in the middle area of south slope of Qinling Mountains

图1 秦岭南坡中段植物群落TWINSPAN分类结果Fig. 1 The TWINSPAN classification for forest communities
2.2植物群落DCA排序
2.2.1森林植物群落DCA排序
利用DCA对秦岭南坡中段93个森林乔木群落样方进行排序,由表2可知,4个排序轴的前两个轴特征值较大且与环境有着较高相关性,具有一定生态意义,而后两个排序轴特征值较小且与环境之间相关性甚微,其生态意义不明确。所以本文根据前两个排序轴作出二维排序图(图2),由图2可知,TWINSPAN分类所得到各个群落类型在二维的排序图上各自有一定的分布范围,基本都呈现出相同植物群落类型聚集在同一个范围内,DCA排序结果基本印证TWINSPAN的分类结果。这也进一步说明了DCA能够较好地反映各个群落之间以及群落与环境之间的关系。
从排序轴来看,第一轴基本上反映各植物群落所在环境的海拔梯度变化,即DCA第一轴从左到右,海拔逐渐增大;第二轴基本上反映各植物群落所在环境的湿度变化,即DCA第二轴从上到下,林下湿度逐步增加。各个群落在排序轴上呈现规律分布,林下环境较为干燥、分布海拔较低的栓皮栎Quercus variabilis植物群落位于排序轴的左上方,林下环境极其湿润,分布在沟谷中山楠群落位于排序轴下方,远离其它群落类型。水杉 Metasequoia glyptostroboides林是人工林,分布海拔较低,常常栽植在河道两侧空旷地上,其林下环境较为干燥,所以水杉林和山核桃Carya cathayensis 群落位于排序轴的上方。巴山冷杉、牛皮桦Betula albo-sinensis var. septentrionalis、太白红杉高海拔分布植物群落位于排序轴右侧。
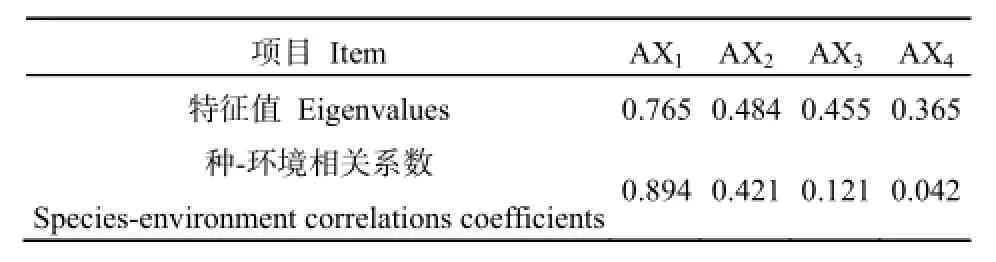
表2 群落DCA排序轴特征值及种-环境相关系数Table 2 Characteristic values and species-environment correlation coefficients on DCA axis forest communities
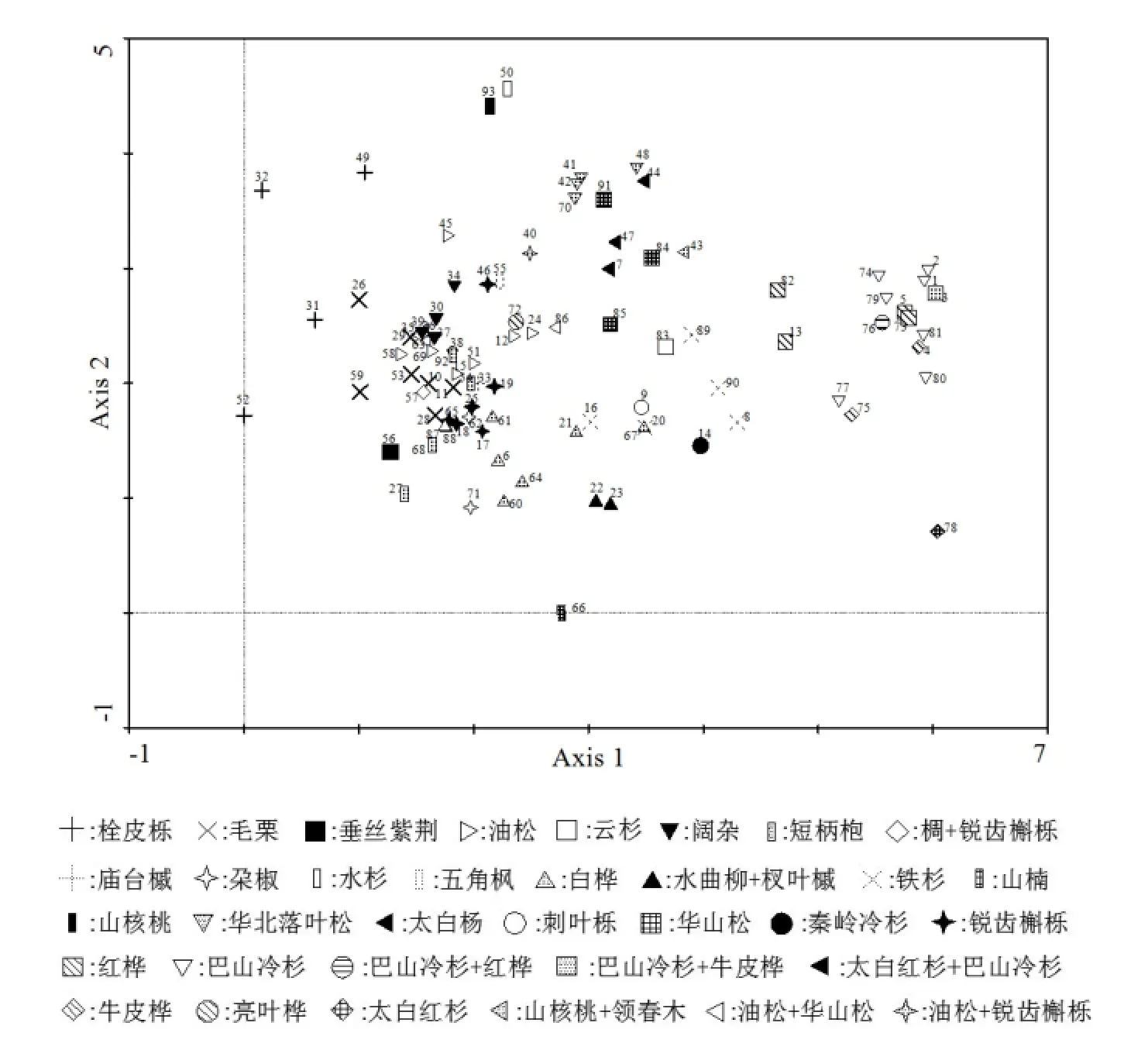
图2 秦岭南坡大熊猫栖息地森林植物群落93个样地DCA二维散布图Fig. 2 A two-dimensional scatter plot of DCA ordination for forest communities
从图2可知阔杂、油松、锐齿槲栎、白桦、板栗、短柄枹栎Quercus glandulifera分布在林下中度湿度、中低海拔区域,位于排序轴中间靠左的位置,各个群落类型分布较为密集,相互之间有一定交错分布。说明这几个群落类型分布生境相近,在物种组成上有一定的相似性,该环境区域是整个研究区植物群落类型分布最多,物种最多的区域。
2.3植物群落分布与环境之间的关系
2.3.1群落样地分布与环境因子的DCCA排序
本研究以秦岭南坡中段主要乔木植物群落物种组成和土壤、地形指标所组成的环境因子矩阵进行DCCA分析,并与DCA排序结果进行分析对比。
从表3可知,第1轴特征值最大(0.701),第2轴次之(0.359),而其它 2个轴的特征值较小,说明前两个轴包含较多信息量,故采用第1轴、第2轴数据作DCCA二维排序图(图3)。图中箭头代表各个环境因子,这是DCCA排序的特点。箭头所处象限,代表着环境因子与排序轴间的正负相关性;箭头连线的长短表示植物群落的分布与该环境因子相关性的大小,箭头连线在排序中的斜率表示环境因子与排序轴相关性的大小,箭头所处的象限表示环境因子与排序轴之间相关性的正负。

表3 DCCA排序轴特征值及种-环境因子相关系数Table 3 Characteristic values and species-environment correlation coefficients on DCCA axis
从图3和表4中可以看出,第一轴基本上反映各植物群落样地所在环境的海拔梯度变化趋势(海拔与DCCA第一轴典范系数0.9648,相关系数为0.9925),即沿DCCA第1轴从左到右,海拔逐步增加;第二轴基本上表现出植物群落所在环境的坡度、全磷、速效磷的变化趋势(全磷与第二轴典范系数为0.6610,相关系数为0.7421;速效磷与第二轴的典范系数为0.5760,相关系数为0.6466;坡度与第二轴的典范系数为-0.4459,相关系数为-0.5006),即沿DCCA第二轴从下到上,坡度渐缓、坡向越向阳,土壤全磷、速效磷含量逐步增加。因此,海拔、全磷、速效磷和坡度是本研究 12个环境因子中影响植物群落分布的几个主导因子。
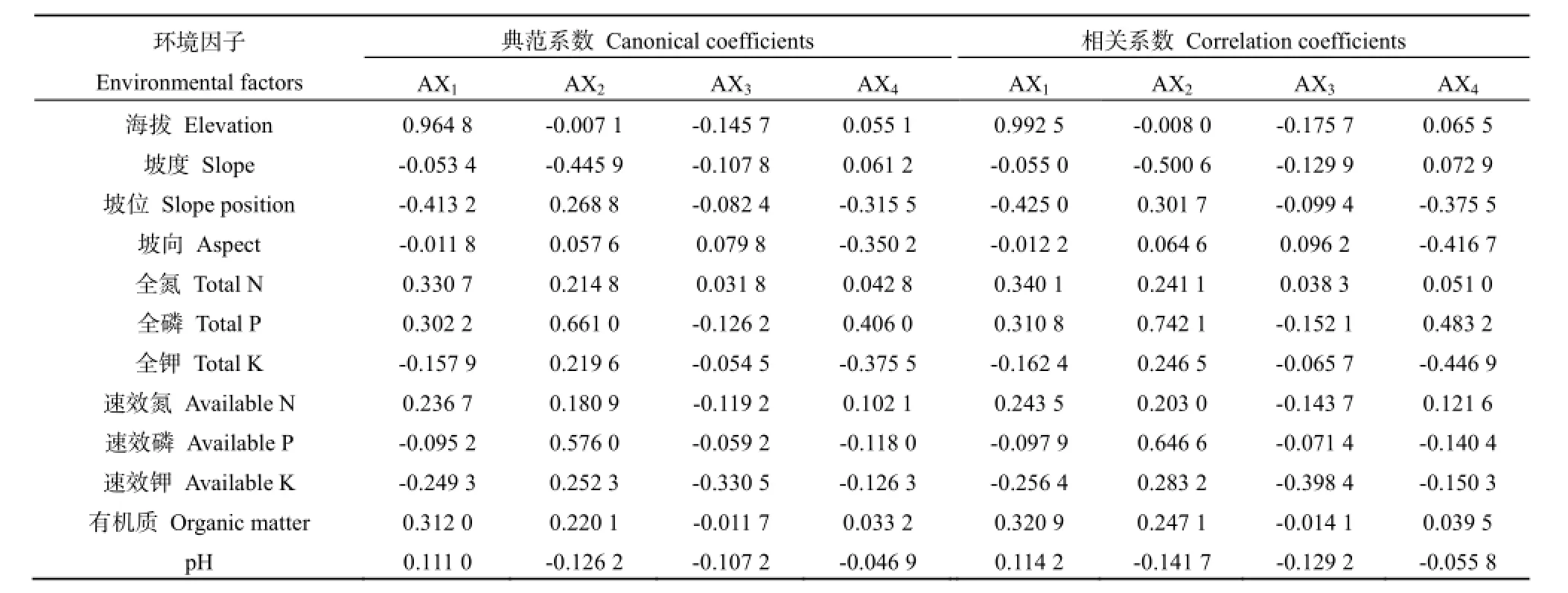
表4 环境因子与DCCA排序轴的典范系数和相关系数Table 4 Canonical coefficients and correlation coefficients of environmental factors with DCA axis

图3 93个样地与12个环境因子的DCCA二维排序图Fig. 3 Two-dimensional DCCA ordination diagram of 93 samples with 12 environmental factors
2.3.2主要优势种分布与环境因子的DCCA排序
通过分析主要优势种的DCCA二维排序图(图4)发现,群落优势种分布格局所揭示的环境梯度与样地群落的分布梯度有着很大相似性。沿着Axis1轴从左至右,乔木层中分布在低海拔的栓皮栎、庙台槭 Acer miaotaiense、枫杨 Pterocarya stenoptera等物种位于排序轴最左侧。随着海拔进一步升高,进入整个研究区中海拔区域,从图4也可以看出这个区域优势种数量明显高于两端,该区域分布乔木有油松、锐齿槲栎、白桦、亮叶桦Betula luminifera等,这些树种分布在各个环境因子中心位置,对环境要求相对较低,所以它们是整个研究区分布最为广泛的几个树种。高海拔太白红杉、巴山冷杉位于排序图的最右侧,与其它乔木层相隔较远,说明其生境极其严酷,以致其它物种无法在此生长。灌木层和草本层也表现出与乔木层相同的变化规律。其中,巴山木竹和秦岭箭竹作为大熊猫的主要食材,它们分别分布在中海拔区域和高海拔区域,这与现实调查结果相吻合。通过上面分析说明,Axis1轴从左至右,海拔逐渐增大,环境温度逐步降低,物种对热量需求逐步减少。从图4也可以看出,Axis2轴与土壤速效磷以及坡度有着密切相关关系,从上至下,坡度逐步增加,土壤速效磷逐步减少。山核桃、白桦、太白杨等位于较平缓山地上,土壤中速效磷的含量较为丰富。秦岭冷杉 Abies chensiensis、铁杉 Tsuga chinensis、杈叶槭 Acer robustum等位于坡度较大山地上,土壤速效磷含量较低。通过分析图4发现,DCCA二维排序图所显示的植物群落优势种分布情况与实际调查记录相一致,说明DCCA二维排序图能较好反应主要优势种与环境梯度之间的关系。因此,海拔、坡度和速效磷是影响优势种分布格局的主要因素,这与群落样地分布和环境梯度关系相似。
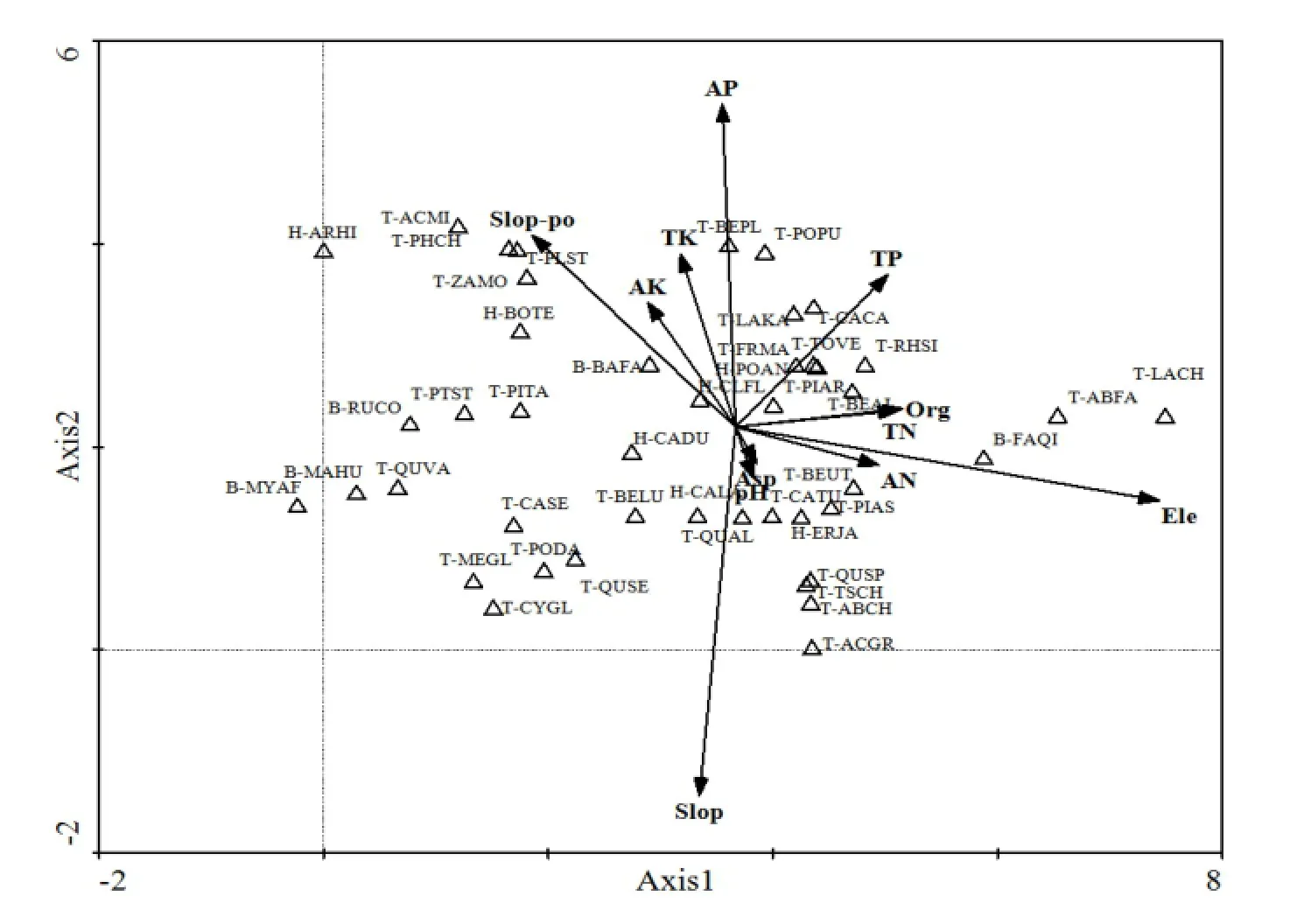
图4 主要优势种与12个环境因子的DCCA二维排序图Fig. 4 Two-dimensional DCCA ordination diagram of main dominant species with 12 environmental factors
2.3.3Monte Carlo检验
从DCCA第一排序轴Monte Carlo检验结果(表5)可以看出,无论是对于群落样地,还是对于优势种分布,海拔、坡度、速效磷和全磷的F值都达到了显著水平(P<0.01),因此,海拔、坡度、速效磷和全磷可以很好地解释植物群落样地和优势物种的分布。该检验结果充分证明前面二维排序图所显示的海拔、坡度、速效磷和全磷是决定植物群落和优势种分布的主要影响因素。
3 结论与讨论
分别采用双向聚类法(TWINSPAN)、去趋势对应分析(DCA)和除趋势典范对应分析(DCCA)对秦岭南坡中段主要乔木群落进行分类与排序,结果表明TWINSPAN双向聚类分析较好地将不同的群落类型进行分类,研究区 93个群落样方被分成34个群落类型。本文利用去趋势对应分析(DCA)对样地进行排序,结果不同类型样地呈现聚集分布,其结果充分验证了TWINSPAN对群落类型划分的科学性。利用DCA对TWINSPAN分类结果进行验证,在以往研究中都得到普遍体现(王得祥等,2009)。

表5 环境因子与DCCA第一排序轴的Monte Carlo显著性检验Table 5 Monte Carlo permutation tests of significance of selected environmental variables with the first canonical axis from DCCA ordinations
从广泛的意义上讲,植物种、植物群落的分布格局是在不同尺度上各种气候、地形、土壤等环境因子综合表现的结果。在区域尺度上讲,气候、植物区系决定特定的植被类型;景观尺度上,如海拔这个因子,使局部环境的水热条件发生改变,导致植被类型发生变化,从而决定植被分布格局;而微生境、土壤养分等决定最终的植物群落类型(Burke,2001)。本文结果表明,在景观和土壤养分这两个尺度上,制约秦岭南坡大熊猫栖息地群落类型、植物种分布格局的主要因素是海拔、坡度、速效磷与全磷。海拔与坡度作为光、热、水条件综合表现,其格局变化也就是光、热、水的空间再次分配,从而使得植物群落组成以及植物种的分布格局发生变化(张峰等,2003;代力民等,2008)。速效磷和全磷作为土壤营养元素重要成分之一,对植物生长有着重要作用,直接影响着植物群落的组成与生理活力,决定着生态系统的结构、功能和生产力水平(Robertson et al.,1981;Vitousek et al.,1989)。综上,海拔、坡度、速效磷和全磷格局变化是光、热、水、土壤营养的格局变化,从而导致植物群落类型及物种分布格局发生变化。
本文通过Monte Carlo显著性检验表明,海拔、坡度、速效磷和全磷能够很好地解释群落样地及优势种在环境梯度上的分布。同时发现DCA、DCCA排序图不仅反映样方间在种类组成上的相似性,而且也反映样方间在环境因子组成上的相似性,而这两种相似性往往相互联系(代力民等,2008)。一般来讲,种类组成接近的植物群落,其环境因子组成也较接近,这是由植物种、植物群落和环境因子之间相互作用的生态关系所决定的。
BURKE A. 2001. Classification and ordination of plant communities of the Naukluft Mountains Namibia [J]. Journal of Vegetation Science, 12(1): 53-60.
PAMER M W. 1993. Putting things in even better order: The advantages of canonical correspondence analysis [J]. Ecology, 74(8): 2215-2230.
ROBERTSON G P, VITOUSEK P M. 1981. Nitrification potentials in primary and secondary Succession [J]. Ecology, 62(2): 376-387.
RUSSELT SMITH J. 1991. Classification, species richness, and environmental relations of monsoon rain forest in northem Australia [J]. Journal of Vegetation Science, 2(2): 259-278.
VEAZQUEZ A. 1994. Multivariate analysis of the vegetation of the volcanoes Taloc and Pelado, Mexico [J]. Journal of Vegetation Science,5(2): 263-270.
VITOUSEK P M, MATSON P A, VANCKVE K. 1989. Nitrogen availability and nitrification during succession, primary, secondary and old field seres [J]. Plant and Soil, 115(2): 229-239.
代力民, 唐立娜, 曹玉明, 等. 2008. 辽东山区生态土地分类中的植物群落数量分析[J]. 林业科学, 44(3): 6-12.
鲍士旦, 江荣风, 杨超光, 等. 1999. 土壤农化分析[M]. 北京: 中国农业出版社: 14-114.
龚志莲, 李勇. 2015. 川西南不同植被木本物种多样性与土壤有机碳积累[J]. 生态环境学报, 24(9): 1447-1452.
江洪, 黄建辉, 陈灵芝, 等. 1994. 东灵山植物群落的排序、数量分类与环境解释[J]. 植物学报, 36(7): 539-551.
李晋鹏, 上官铁梁, 孟东平, 等. 2007. 山西吕梁山南段植物群落的数量分类和排序研究[J]. 应用与环境生物学报, 13(5): 615-619.
李思阳, 张娟, 姚玲爱, 等. 2016. 西枝江流域浮游植物群落结构特征与主要环境因子的关系研究[J]. 环境科学学报, 36(6): 1939-1947.
宋爱云, 刘世荣, 史作, 等. 2006. 卧龙自然保护区亚高山草甸植物群落物种多样性研究[J]. 林业科学研究, 19(6): 767-772.
王得祥, 蔺雨阳, 雷瑞德, 等. 2009. 秦岭山地天然油松林群落结构特征和数量分类研究[J]. 西北植物学报, 29(5): 1016-1025.
王国宏, 杨利民. 2001. 祁连山北坡中段森林植被梯度分析及环境解释[J]. 植物生态学报, 25(6): 733-740.
王晶, 张钦弟, 许强, 等. 2016. 山西庞泉沟银露梅群落物种多样性的海拔格局[J]. 植物学报, 51(3): 335-342.
王世雄, 王孝安, 李国庆, 等. 2010. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释[J]. 生态学报, 30(6): 1638-1647.
王宇超, 王得祥, 胡有宁, 等. 2012. 陕西黑河上游主要天然林类型及物种多样性特征研究[J]. 西北农林科大学学报(自然科学版), 40(7): 106-112.
孙菊, 李秀珍, 胡远满, 等. 2009. 大兴安岭沟谷冻土湿地植物群落分类、物种多样性和物种分布梯度[J]. 应用生态学报, 20(9): 2049-2056.徐广平, 张德罡, 蒲小鹏, 等. 2005. 东祁连山高寒草地植物群落二维极点排序[J]. 草原与草坪, 3: 42-45.
张峰, 张金屯. 2003. 历山自然保护区猪尾沟森林群落植被格局及环境解释[J]. 生态学报, 23(3): 421-427.
张玲, 袁晓颖, 张东来. 2007. 大小兴安岭过渡区木本植物群落数量分类与排序[J]. 东北林业大学学报, 35(9): 49-51.
张先平, 王孟本, 佘波, 等. 2006. 庞泉沟国家自然保护区森林群落的数量分类和排序[J]. 生态学报, 26(3): 754-761.
张新时. 1991. 西藏阿里植物群落的间接梯度分析、数量分类与环境解释[J]. 植物生态学报, 15(2): 101-113.
The Quantitative Classification and Environmental Interpretation of Forest Communities in the Middle Area of South Slope of Qinling Mountains
WANG Yuchao1, ZHOU Yafu1, WANG Dexiang2*
1. Xi'an Botanical Garden of Shaanxi Province, Xi'an 710061, China;2. College of forestry, Northwest A & F Unversity, Yangling 712100, China
In order to revealed the plant community distribution pattern and the corresponding relation with environment. Tow line transects were set between the altitude of 900~3 071 m in the field, and 93 standard plots (25 m×20 m) were investigated on transects. Vegetation community, growth status and distribution environment were recorded and soil samples were collected in the standard plots. Based on analysis of plots and soil data, the main forests in giant panda habitat were classified and ordering by using the two-way indicator species analysis (TWINSPAN), Detrended Corresponding Analysis (DCA) and Detrended Canonical Correspondence Analysis (DCCA). The results showed that: (1) 93 standard plots were classified into 34 plant communities by Two-way Indicators Species Analysis (TWINSPAN). The result of ordering by DCA showed that the standard plots fit to aggregation distribution and verified the scientificity of TWINSPAN. (2) The restrictive factors of community type and species distribution were altitude, slope, rapidly-available phosphorus and total phosphorus at the landscape level by the test of significance between DCCA and Monte Carlo. Factors of altitude, slope, rapidly-available phosphorus and total phosphorus can explain distribution of communities and dominant species at the landscape level, and the species and environmental factors show great similarities between different plots.
Qinling Mountains; south slope; TWINSPAN; DCA; DCCA
10.16258/j.cnki.1674-5906.2016.06.008
Q948; X171.1
A
1674-5906(2016)06-0965-08
陕西省科学院科技计划项目(2014k-25);陕西省科学院科技重点项目(2013K-01);国家科技基础性工作专项重点项目(2007FY110800)
王宇超(1978年生),男,博士,主要从事森林生态以及群落生态学研究。Email: wangyuchao2000@126.com
王得祥(1966年生),男,教授,博士生导师,主要从事森林生态及森林健康可持续经营研究。Email: wangdx66@126.com
2016-06-08
