南亚热带杉木林分改造中不同树种组合模式评价
2016-09-19黄钰辉张卫强甘先华唐洪辉盘李军冼杆标1广东省森林病虫害生物防治重点实验室广东广州510520广东省林业科学研究院广东广州510520广东省佛山市云勇生态林养护中心广东佛山528518
黄钰辉,张卫强*,甘先华,唐洪辉,盘李军,冼杆标1. 广东省森林病虫害生物防治重点实验室,广东 广州 510520;2. 广东省林业科学研究院,广东 广州 510520;. 广东省佛山市云勇生态林养护中心,广东 佛山 528518
南亚热带杉木林分改造中不同树种组合模式评价
黄钰辉1, 2,张卫强1, 2*,甘先华1, 2,唐洪辉1, 2,盘李军3,冼杆标3
1. 广东省森林病虫害生物防治重点实验室,广东 广州 510520;2. 广东省林业科学研究院,广东 广州 510520;3. 广东省佛山市云勇生态林养护中心,广东 佛山 528518
低效杉木(Cunninghamia lanceolata)人工林的改造在我国林业建设中占有重要地位,其目标主要是促进低效杉木人工林转变为结构复杂的针阔混交林,从而提高其生态系统功能。以南亚热带地区杉木林改造后的针阔混交林为研究对象,比较了不同树种组合模式的土壤理化性质、碳储量和乔木层生物多样性的差异,并采用主成分分析对不同树种组合模式进行综合评价。结果表明,土壤理化性质在不同树种组合模式间存在显著差异。生态系统碳储量介于110.01~183.24 t·hm-2之间,主要存储于土壤层(70.1%~85.7%),其次是植被层(11.5%~25.8%)和凋落物层(0.8%~5.2%);植被层中 85%以上的碳集中于乔木层。不同树种组合模式的生物多样性差异明显,其中丰富度指数的差异最大。采用主成分分析的方法,将26个评价指标提取为5个主成分,可反映针阔混交林改造模式92.7%的变异性。综合评价得分表明,在本地区对杉木林进行皆伐改造时,优化的树种组合模式为:(1)杉木+阴香(Cinnamomum burmanni)+米老排(Mytilaria laosensis)+红锥(Castanopsis hystrix)+火力楠(Michelia macclurei);(2)杉木+米老排+阴香+山杜英(Elaeocarpus sylvestris)+枫香(Liquidambar formosana);(3)杉木+红花荷(Rhodoleia championii)+刺桐(Erythrina variegata)+火焰木(Spathodea campanulata)+灰木莲(Mangletia glauca)。研究结果可为南亚热带地区低效针叶林改造、生态林经营及生态系统服务功能评估提供参考。
杉木;林分改造;树种组合;主成分分析;综合评价;南亚热带
引用格式:黄钰辉, 张卫强, 甘先华, 唐洪辉, 盘李军, 冼杆标. 南亚热带杉木林分改造中不同树种组合模式评价[J]. 生态环境学报, 2016, 25(6): 956-964.
HUANG Yuhui, ZHANG Weiqiang, GAN Xianhua, TANG Honghui, PAN Lijun, XIAN Ganbiao. Evaluation of Different Tree Species Composition Patterns in Stand Conversion of Cinnamomum burmanni in Southern Subtropical Region of China [J]. Ecology and Environmental Sciences, 2016, 25(6): 956-964.
杉木(Cunninghamia lanceolata)是我国亚热带地区主要的速生用材树种之一,具有生长快、产量高和材质好的特点,因此在我国南方地区广泛种植。第八次全国森林资源清查结果显示其人工林面积达1.096×107hm2,占南方人工林面积的60%~80%(刘世荣等,2005),在我国森林资源结构中占有重要地位。由于长期采取纯林、连栽、短轮伐期和皆伐等经营方式,杉木林生产力和土壤肥力逐代下降(陈龙池等,2004;田大伦等,2011),同时抗逆性差、自我更新能力弱、生态服务功能低下等问题也逐渐暴露出来(刘庆等,2010),因此亟需进行林分改造。林分改造是当前世界范围内森林经营的一项重要措施(Gärtner et al.,2004;Knoke et al.,2008),其目的是将低效人工针叶纯林改造为结构复杂、更新能力和生态服务功能强的混交林生态系统。在开展林分改造时,树种的选择及其配置模式是决定群落演替的关键因素,这是由于不同的树种适宜的立地条件不同,而种间关系也可随树种的种类、径级、距离以及时间等因素而改变(Kelty,2006;Getzin et al.,2006),进而影响林分生产力、土壤肥力和生物多样性等。因此,在林业生产实践中,为取得更好的经济效益和生态效益,一般需要筛选优化的林分改造模式加以推广。目前对林分改造模式的评价一般是对林木生长(Meng et al.,2014)、物种多样性(罗应华等,2013;何友均等,2013a)、土壤养分(何友均等,2013b)2489-2495等单一目标的评价,而全面反映林分改造效果的多因子综合评价相对较少(吕海龙等,2011)。
本文以杉木林皆伐改造形成的不同树种组合模式的针阔混交林为研究对象,分析不同树种组合模式的针阔混交林土壤理化性质、森林生态系统碳储量和生物多样性的差异,运用主成分分析对不同树种组合模式进行综合评价,旨在为珠三角低效针叶林改造、生态林经营及生态服务功能评估提供参考。
1 研究区域与方法
1.1研究区域概况
研究区位于广东省佛山市高明区云勇林场,北纬22°43′,东经112°40′,属南亚热带湿润季风气候,水热资源丰富。年平均气温、最高气温和最低气温分别为22.0、34.5和3.5 ℃,雨量充沛,年降水量平均达2000 mm,集中在4─8月。地势属丘陵地带,土壤为花岗岩发育的酸性赤红壤,土层深厚。2002年起,为了营造生态公益林,对杉木林进行皆伐改造,保留单株杉木萌芽条,引入不同类型乡土树种造林。造林前整地,造林当年抚育2次,造林后第2~3年各抚育1次,抚育措施包括铲草、松土、扩穴和培土。造林阔叶树种包括刺桐(Erythrina
variegata)、大叶紫薇(Lagerstroemia speciosa)、凤凰木(Delonix regia)、枫香(Liquidambar formosana)、格木(Erythrophleum fordii)、海南蒲桃(Syzygium hainanense)、海南石梓(Gmelina hainanensis)、红花荷(Rhodoleia championii)、红花羊蹄甲(Bauhinia blakeana)、红荷木(Schima wallichii)、红锥(Castanopsis hystrix)、灰木莲(Mangletia glauca)、火力楠(Michelia macclurei)、火焰木(Spathodea campanulata)、假苹婆(Sterculia lanceolata)、黧蒴(Castanopsis fissa)、米老排(Mytilaria laosensis)、木荷(Schima superba)、盆架子(Winchia calophylla)、山杜英(Elaeocarpus sylvestris)、香樟(Cinnamomum camphora)、阴香(Cinnamomum burmanni)。2014年8─10月,选择不同树种组合模式的 12个针阔混交林作为研究对象,林分地形地貌、海拔、母岩、土壤类型等相同或相近,且彼此相连,每个改造模式林分内设置1个研究样地,共12个研究样地(表1)。
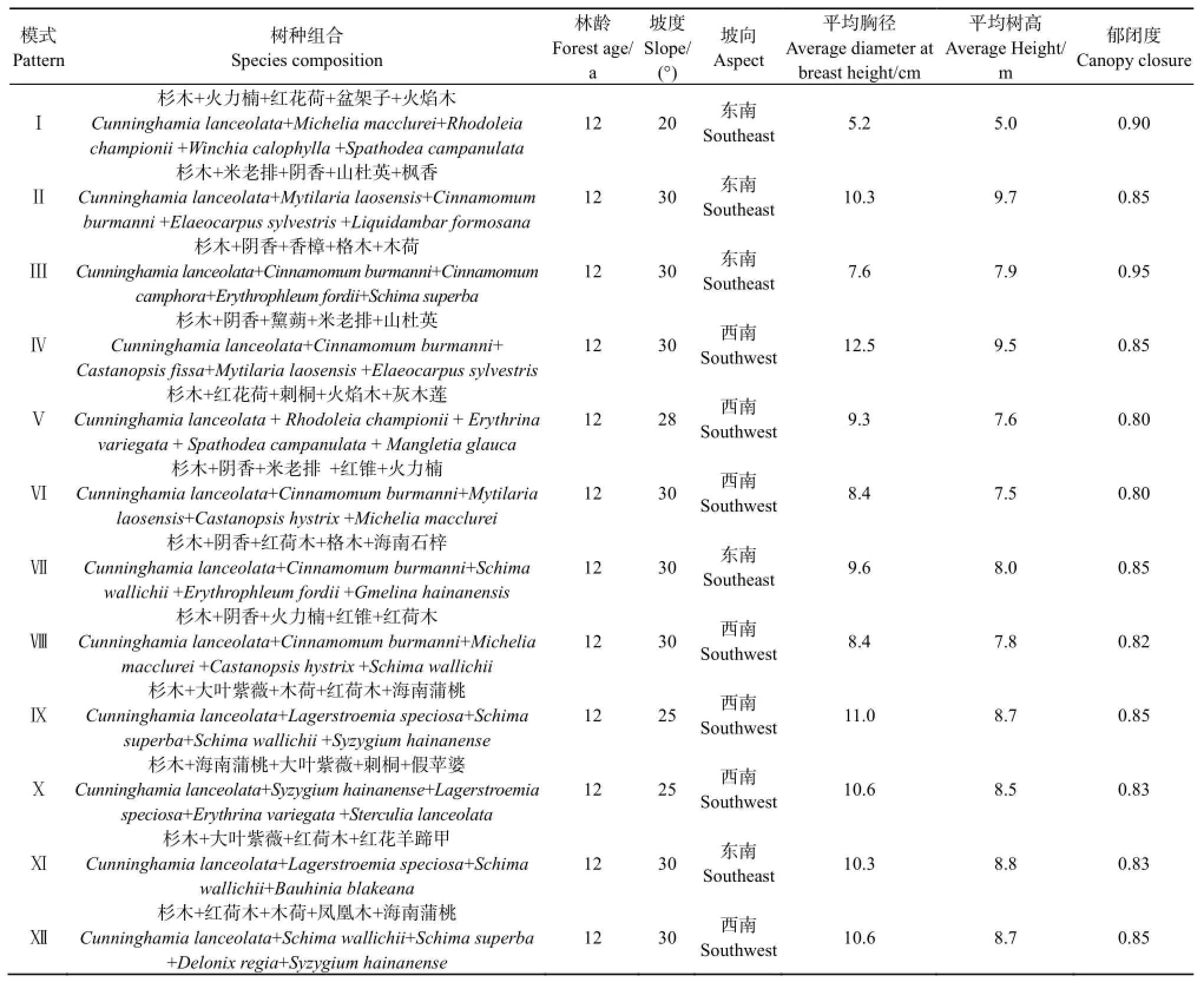
表1 不同树种组合模式林地概况Table 1 Plantations characteristics of different tree species composition patterns
1.2土壤理化性质测定
2014年8月,在每个样地内选择有代表性的部位,分别挖取3个土壤剖面,按照0~25、25~50、50~75和75~100 cm土壤深度用容积100 cm3环刀和密封袋取样,各层重复取样3次。参考《森林土壤水分——物理性质的测定LY/T 1215─1999》测量土壤容重、总孔隙度、毛管孔隙度、非毛管空隙度、土壤通气性、自然含水量、最大持水量、毛管持水量和田间持水量等指标。将密封袋中土壤样品风干、研磨、过筛后,采用pH计测定土壤pH值,重铬酸钾法测定有机质,半微量凯式法测定全氮,碱解扩散法测定速效氮,NaOH熔融-钼锑抗比色法测定全磷,NaHCO3浸提法测定速效磷。结果中土壤理化性质各指标的数值为4个土层的平均值。
1.3森林植被碳储量测定
1.3.1乔木碳储量测定
2014年10月,在每个研究样地内设计1个固定样方,样方规格为20 m×20 m,调查树高在1.5 m(含1.5 m)以上的乔木树高和胸径。各样地乔木的生物量测算采用已有生物量相对生长方程和样地调查数据相结合的方法(王卫霞等,2013)。乔木各器官及森林总生物量与测树因子间普遍存在着相关关系,如米老排(明安刚等,2012)、红锥(刘恩等,2012)、杉木(俞月凤等,2013)、樟树(姚迎九等,2003)、木荷(曾小平等,2008)等,这种相关关系可以用数学模型进行拟合。根据已有生物量预测模型并结合样地实测数据可估算出杉木林改造主要造林树种树干、树枝、树叶和树根生物量,其他常绿阔叶树种生物量参考相关文献计算获得(李静鹏等,2013;杨昆等,2007)。结合树干、树枝、树叶及树根碳含量,利用各器官加权平均法,估算出森林植被碳储量。
1.3.2灌木和草本碳储量测定
2014年10月,在每个研究样地内设置5个2 m×2 m调查小样方,分别收获小样方内的全部灌木和草本并称取鲜重,对灌木和草本进行取样,带回实验室在85 ℃的恒温箱中烘干至恒重,由干重/鲜重比计算含水率,结合灌木和草本碳含量(碳含量采用重铬酸钾-氧化法测定),计算灌木和草本碳储量。
1.4凋落物碳储量测定
2014年10月,在每个研究样地内设置5个2 m×2 m的小样方,收集各小样方内全部凋落物,立即称量鲜重,并对凋落物进行取样,带回实验室在85 ℃的恒温箱中烘干至恒重,由干重/鲜重比计算含水率,结合凋落物碳含量(碳含量采用重铬酸钾-氧化法测定),计算凋落物的碳储量。
1.5土壤碳储量测定
采用重铬酸钾-水合加热法测定土壤有机碳含量。土壤层碳储量的计算采用如下公式:(1)
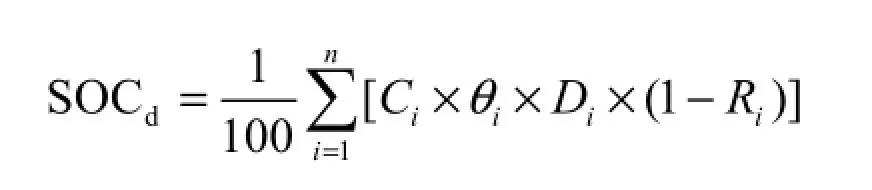
式中,SOCd为土壤有机碳储量(t·hm-2);n为土层数;Ci为第i层土壤的有机碳含量(%);θi为第i层土壤容重(g·cm-3);Di为第i层土壤厚度(cm);Ri为第i层土壤中直径大于2 mm石砾体积含量百分比。
1.6乔木多样性
根据乔木调查的树高、胸径和枝下高计算生物多样性,生物多样性指标选取丰富度指数、Shannon-Wiener指数、Simpson指数和Pielou均匀度指数,计算方法参考相关文献(马克平等,1994)。
1.7数据分析
运用 Excel将数据进行整理和预处理,利用SPSS 16.0进行单因素方差(one-way ANOVA)分析和主成分分析。采用Duncan法比较各指标在各研究样地间的差异,显著性水平设为α=0.05。
在进行综合评价时,选取反映不同树种组合改造模式综合评估值的n个指标为X1,X2,……,Xn,m个改造模式的 n项指标构成原始数据矩阵X=[Xij]n×m,其中Xij为第i个模式的第j项指标数据(i=1,2,…,n;j=1,2,…,m)。
第一步,为消除量纲和数量级的影响,按公式(2)对原始数据进行标准化:

第二步,确定主成分,将标准化的数据用SPSS软件处理,从总方差分析表选取累积贡献率≥85%的前p个主成分,然后建立p个主成分和标准化变量的关系,计算公式为:
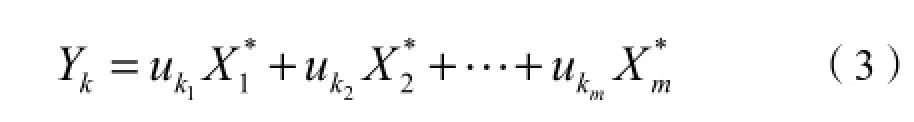
式中,Yk是第k个主成分(k=1,2,…,p),uk1为第k个主成分的因子荷载。
第三步,确定权重,用第k个主成分的贡献率与选取的p个主成分的总贡献率的比值来确定每个主成分的权重,计算公式如下:

式中,ωk为第k个主成分的权重,λk为第k个主成分的贡献率。
第四步,构造综合评价函数,根据前p个主成分及其权重构造综合评价函数,公式如下:

式中,F为不同树种组合改造模式的综合评价得分,分值越高,表明该树种组合模式的效果越好。
2 结果与分析
2.1土壤物理性质
不同树种组合改造模式的土壤物理性质见表2。不同模式的土壤容重介于1.31~1.60 g·cm-3之间;土壤总孔隙度介于38.2%~49.2%之间,其中非毛管孔隙度远低于毛管孔隙度。不同模式的土壤自然含水量介于19.5%~28.9%之间,最大持水量、毛管持水量及田间持水量分别介于 382.2~491.7、353.9~459.4及326.8~431.7 mm之间。方差分析表明,土壤物理性质各指标在不同模式之间的差异显著(P<0.05)。12种模式中,模式Ⅵ的土壤容重最小,总孔隙度、自然含水量、最大持水量和田间持水量最大,其余指标值也相对较大;模式Ⅳ的土壤容重和土壤通气性最大,但自然含水量和田间持水量最低;模式Ⅺ的总孔隙度、毛管孔隙度、自然持水量和毛管持水量均最低,其余指标值也相对较低。

表2 不同树种组合模式的土壤物理性质Table 2 Soil physical properties of plantations of different species composition patterns
2.2土壤化学性质
从表3可知,土壤pH值介于3.89~4.69,呈强酸性。土壤有机质的变化范围较大,介于9.56~18.64 g·kg-1之间。全N和全P的变化范围分别是0.46~0.86 和 0.11~1.10 g·kg-1,速效 N和速效 P分别介于44.02~81.22和0.45~2.28 mg·kg-1之间。12种模式中,模式Ⅷ的土壤有机质、氮、磷含量均为最低;模式Ⅱ的pH最低,全N和速效N含量最高,其余指标值也较大;模式Ⅰ和Ⅲ的全磷含量显著高于其他模式,是其他模式的3~14倍,而速效P含量却偏低。

表3 不同树种组合模式的土壤化学性质Table 3 Soil chemical properties of plantations of different species composition patterns
2.3森林生态系统碳储量及乔木多样性
从表 4可知,针阔混交林总碳储量均值介于110.01~183.24 t·hm-2之间。针阔混交林碳储量分配格局表现为土壤层>植被层(乔木、灌木、草本)>凋落物层。土壤层碳储量(81.87~144.35 t·hm-2)占总储量的70.1%~85.7%;植被层的碳储量(17.68~43.48 t·hm-2)占总碳储量的13.4%~26.9%,其中乔木层碳储量占植被层碳储量的 85.8%~99.8%。凋落物层碳储量介于1.17~4.84 t·hm-2之间,仅占总碳储量的0.8%~5.2%。模式Ⅳ由于土壤和乔木碳储量都较大而具有最高的总碳储量;模式V的土壤碳储量最大;模式Ⅲ的乔木层碳储量最大,但其土壤碳储量偏低;模式Ⅷ的土壤碳储量最小,乔木碳储量也较小,故总碳储量最低。

表4 不同树种组合模式的碳储量及比例Table 4 Carbon Storage and biodiversity indexes of tree layer in different species composition patterns
不同模式乔木层丰富度指数介于8~32之间(图1a),Shannon-Wiener指数、Simpson指数及Pielou指数分别介于1.59~2.82、0.67~0.90和0.59~0.95之间(图1b~d)。模式Ⅰ的丰富度指数、Shannon-Wiener指数和Simpson指数均是12种林分中最大的,其Pielou指数也较大,说明该模式的生物多样性较高,物种丰富且分布均匀;模式Ⅴ和Ⅻ的丰富度指数最小,说明这两种模式与改造初始阶段相比仅有少数物种侵入,两者的区别在于前者的物种分布比后者均匀;模式Ⅲ和Ⅳ的丰富度指数相对较高,但其他多样性指数相对较低,说明这两种模式受到不少树种入侵,但分布不均。
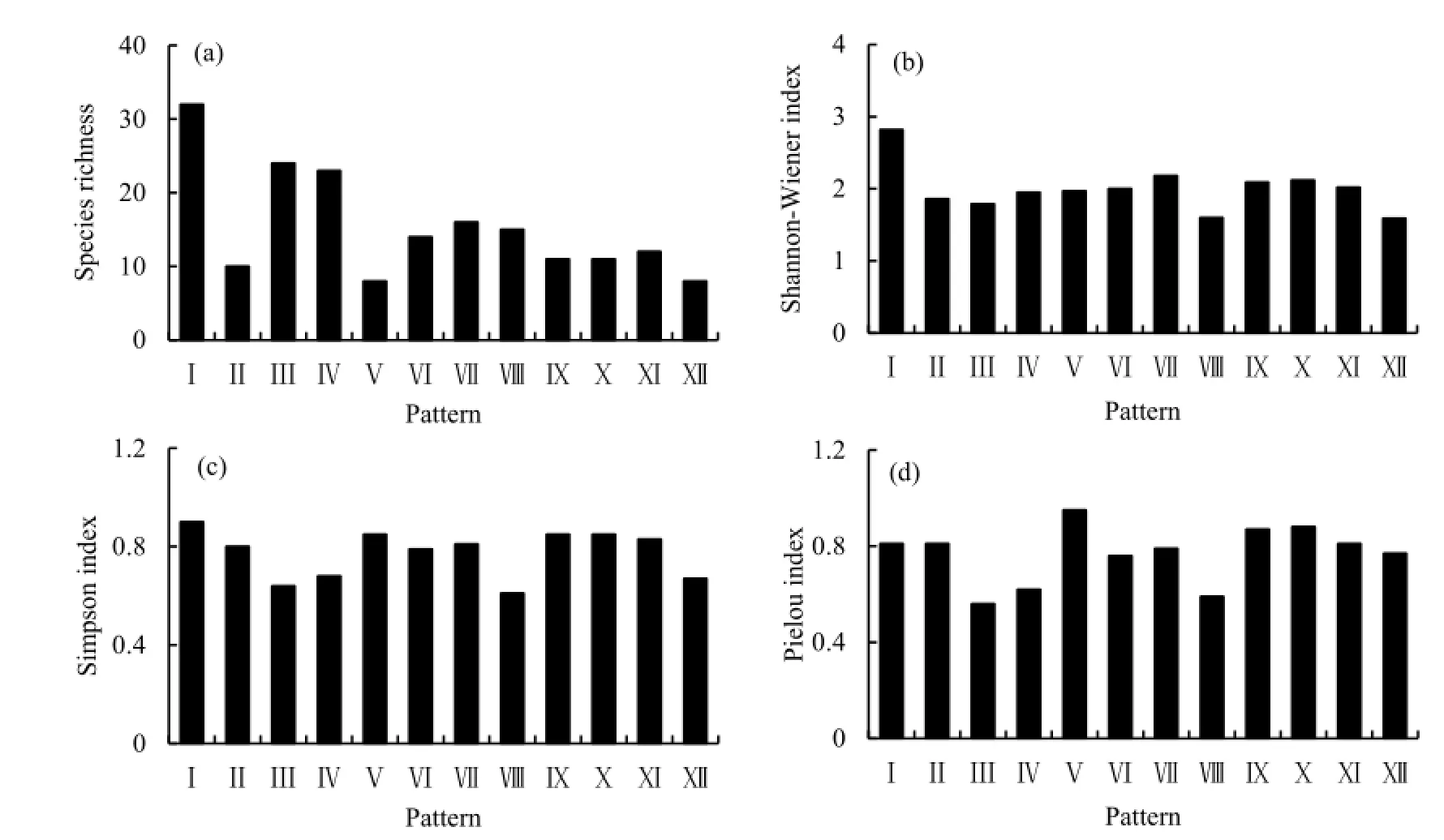
图1 不同树种组合模式的乔木层多样性Fig. 1 Species diversity of different species composition patterns
2.4不同树种组合模式综合评价
以不同树种组合模式主要造林树种生长状况、土壤物化性质指标、碳储量和生物多样性共 26个评价指标进行主成分分析,并根据特征值≥1的原则,提取了5个主成分(表5)。前5个主成分的累计贡献率达92.701%,基本上能反映不同树种组合模式系统内的变异信息。第一主成分在最大持水量、总孔隙度、毛管孔隙度、毛管持水量、自然含水量和田间持水量上有较大的载荷;第二主成分在有机质含量、土壤碳储量、全氮含量、速效N含量上有较大的载荷;第三主成分载荷较大的指标为灌木碳储量和丰富度指数;第四主成分中Simpson指数和Pielou指数的载荷较大;第五主成分中土壤通气性和非毛管孔隙度的载荷较大。
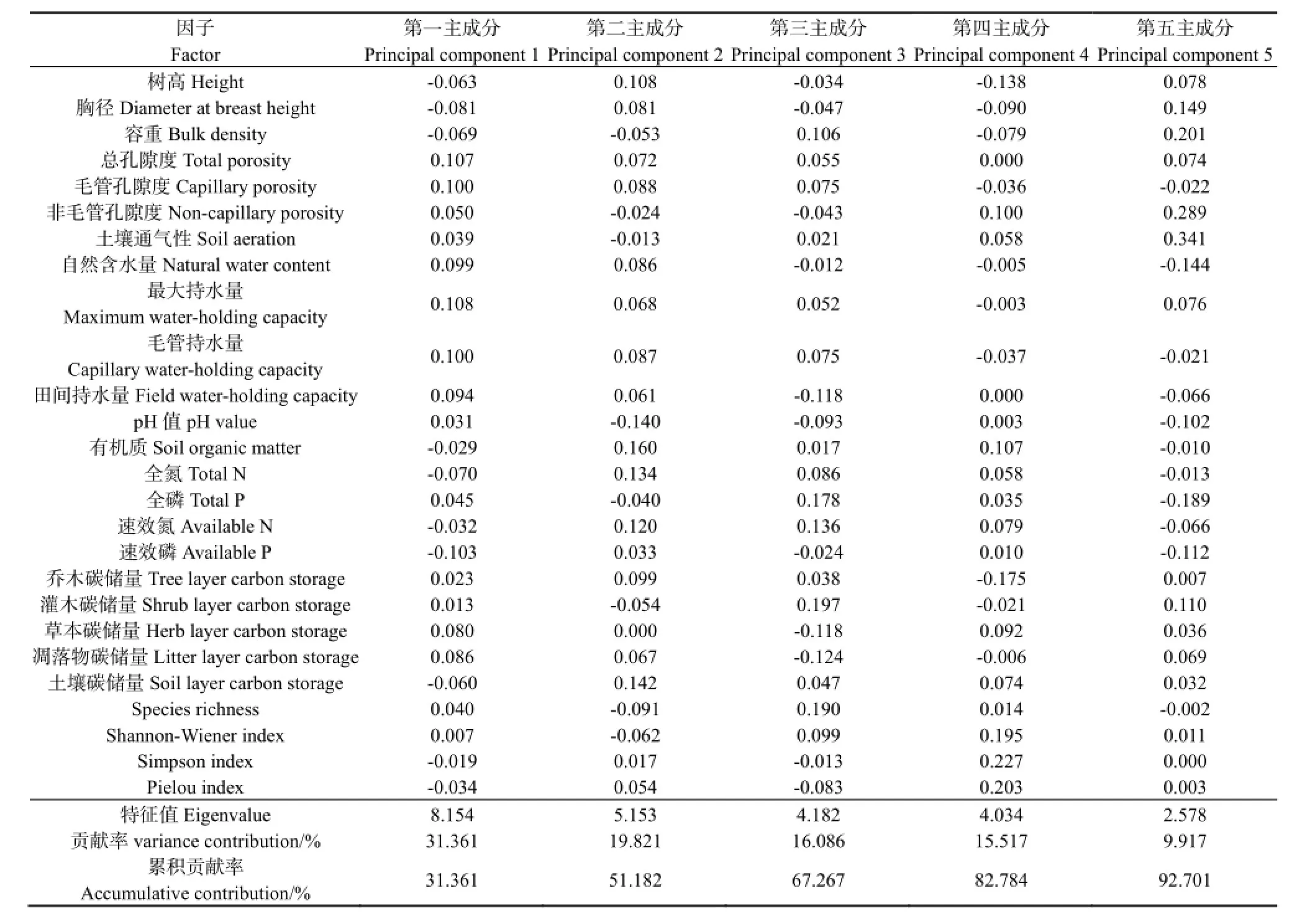
表5 不同树种组合模式林分评价的主成分分析结果Table 5 The results of the principal component analysis for the plantations evaluation of different species composition patterns
根据主成分分析的特征值计算综合评价得分,结果见表6。由表6可知,综合评价得分位列前三的树种组合模式分别为:模式Ⅵ(杉木+阴香+米老排+红锥+火力楠)、模式Ⅱ(杉木+米老排+阴香+山杜英+枫香)、模式Ⅴ(杉木+红花荷+刺桐+火焰木+灰木莲)。
3 讨论
3.1不同树种组合对土壤理化性质的影响
针阔混交林土壤理化性质在不同树种组合模式间存在显著差异,这说明不同树种对土壤质地存在显著影响,这在以前的研究中也有类似的发现(Binkley,19951-33;Augusto et al.,2002;郭琦等,2014)。植被与土壤之间的相互作用是一个复杂的过程,植物可以通过凋落物等影响土壤的化学性质,还可以通过根系活动等影响土壤的物理性质(Binkley,19951-33),而土壤质地、养分状况又反过来影响植物的生长。本研究中相关树种对土壤理化性质的影响已有报道,如康冰等(2010)发现杉木与红锥混交后土壤有机质含量提高,格木作为豆科树种可提高土壤全氮含量,米老排纯林的土壤容重较小;林德喜等(2004)发现采用火力楠对马尾松纯林进行改造,改善了土壤孔隙状况;庄雪影等(2012)的研究表明火力楠和米老排对土壤肥力的改良优于楝叶吴茱萸。然而,对于纯林或简单组合模式,树种的影响较易确定,而多树种混交时可能需要对每一物种的生长状况、空间分布格局等进行深入研究才能了解其影响机制。
在土壤物理性质方面,本研究不同树种组合模式的土壤容重(1.31~1.60 g·cm-3)略高于杉木林皆伐1 a后的早期研究结果(薛立等,2005)1417-1421,多数与鼎湖山的马尾松林和针阔混交林接近,而高于地带性植被季风常绿阔叶林(李德军等,2004)。需要注意的是,本研究林分林龄仅为12 a,土壤容重是0~100 cm的平均值,而容重一般随土层加深而增大,故推测本研究的容重等土壤物理性质与林分改造初期差别不大,这与何友均等(2013b)2484-2495等对近自然改造早期土壤物理性质的研究结论一致。在土壤化学性质方面,土壤全磷和有效P含量与早期对当地杉木林改造的结果相近(薛立等,20051417-1421;李国平等,200775-79),但部分模式存在极端值,如模式I和Ⅲ的全磷含量(1.1 g·kg-1)虽与亚热带一些地区相近(何友均等,2013b)2484-2495,却是本研究中其他模式的数倍,而其速效P含量相对又较低,表明这两种模式的P元素可能被吸附或沉淀固定。在热带亚热带地区的酸性土壤中,P元素容易与Fe、Al等离子形成难溶性的磷酸盐沉淀物(Chapin et al.,2002)。与李国平等(2007)75-79对杉木林的研究相比,多数模式的土壤有机质、全氮和有效N含量均比杉木林有所提高,全磷和有效 P则降低。何友均等(2013b)2484-2495认为这可能是因为杉木快速生长吸收P元素,同时林分改造导致微环境改变,淋溶加强后造成养分流失。

表6 不同树种组合模式林分的综合评价Table 6 Comprehensive evaluation for plantations of different species composition patterns
3.2不同树种组合对针阔混交林碳储量的影响
不同树种组合模式的针阔混交林碳储量(110.01~183.24 t·hm-2)低于我国森林生态系统平均碳储量(258.83 t·hm-2),更远低于针阔混交林和常绿阔叶林(周玉荣等,2000)518-522,这主要是由于本研究林分的林龄较低,森林生态系统的碳储量还远远未达到其潜力。林分碳储量的空间分布格局以土壤层最高,其次是植被层,与此前的研究一致(Finér et al.,2003;周玉荣等,2000518-522)。本研究区12 a生杉木林0~100 cm土层的土壤碳储量为162.1 t·hm-2(薛立等,2012),高于本研究的所有针阔混交林,可能是因为后者是采用皆伐的方式改造形成,皆伐导致了土壤碳排放升高,而针阔混交林在幼龄林阶段土壤碳输入低于中龄杉木林,故土壤碳储量有所下降。针阔混交林的乔木层碳储量(15.17~41.71 t·hm-2)是植被层碳储量最大的组分,不同模式间的差异较大,这主要是由树种组成不同造成的。本研究区在进行杉木纯林改造时,所选择的树种既有速生的乡土阔叶树种,如黧蒴、米老排等,又有景观树种,如大叶紫薇、凤凰木、红花羊蹄甲等,还有格木、红锥、香樟等珍贵树种,不同类型的树种其生长差异较大,因此不同的树种往往对碳储量有较大的影响(Thompson et al.,2005)355-358。此外,种间关系对树木的生长也有很大的影响,如杉木和火力楠的纯林碳储量低于二者的混交林(黄宇等,2005),这可能是因为纯林中个体间因生态位重叠而竞争激烈,树木生长缓慢,而混交林中不同种类个体间的竞争减弱,反而有利于生长。
3.3不同树种组合对生物多样性的影响
本研究的 12种林分在改造初始阶段均为杉木与3~4种阔叶树种混交,但现阶段生物多样性在不同树种组合模式间有着明显的差别,结合碳储量的特征分析可知,不同林分对其他树种的入侵和竞争的响应不同:模式Ⅲ和Ⅳ受其他树种入侵后,分布均匀度相对较低,降低了个体间的竞争,植株生长状况良好,乔木层碳储量较高,目前不少研究已证实物种丰富度与生物量增量之间的正相关关系(Tilman,1999;Balvanera et al.,2006),补偿效应(生态位分化和促进)和采样效应(特殊功能特征的选择)是解释这种关系的两个主要机理(Paquette et al.,2011);而模式Ⅰ的树种组合面对其他树种的入侵,物种间分布相对均匀,个体间竞争较强,植株生长状况不佳(表 1),植被层碳储量较低,即多样性与生物量之间存在负相关关系(Thompson et al.,2005355-358;Symstad et al.,2003)。此外还应注意生态系统功能受环境要素的影响可能更大(Hooper et al.,2005),且环境异质性甚至可以掩盖物种多样性与生态系统功能之间的关系(Fridley,2002)。
3.4林分改造的综合评价
本研究综合评价得分较高的 3种树种组合模式,其共同点主要是土壤物理性质较好,但是对评价结果的应用仍需谨慎,这是因为主成分分析的应用具有一定的局限性,尤其是主成分各指标系数的正负,可能与评价的实际意义不符(侯文,2006)。本研究中,第一主成分因子负荷较大的主要是土壤物理性质指标,第二主成分对应的主要是土壤化学性质指标,可见综合评价中土壤特征的权重较大。综合评价主要是为了提高低效人工林改造的成效,除改良土壤外,提高碳储量和生物多样性等指标一般也是林分改造的主要目的,因此对林业生产提供指导和参考时,评价结果还需根据林分改造的目的进行一定的修正。本研究的模式Ⅱ除丰富度指数略低外,多数指标水平较高;得分最高的模式Ⅵ的土壤肥力相对较差,土壤碳储量也较低,但乔木层碳储量较高,凋落物碳储量最大,表明该模式植物可能通过快速吸收土壤养分来维持快速生长,但凋落物养分归还较慢,可能会限制林分后期的生长;模式Ⅴ的有效P含量和丰富度指数均较低,乔木层碳储量也不高,可能是受到P限制。模式Ⅷ和Ⅺ的综合评价得分较低,前者土壤养分最差,乔木层碳储量也偏低,植物生长可能受到土壤养分的限制;后者的土壤养分水平较好,速效P含量最高,但乔木层碳储量偏低,可能是树种间的竞争过于激烈,因此应尽量避免运用这两种树种组合模式改造本地区的杉木林。
4 结论
对南亚热带杉木人工林进行皆伐改造,不同树种组合模式间土壤理化性质的差异显著。生态系统碳储量和生物多样性在不同树种组合模式间差异较大,表明不同树种组合对非造林树种的入侵和竞争的响应不同;林分固定的碳主要集中于0~100 cm土壤层,其次是植被层。主成分分析显示土壤理化性质在综合评价中权重较大。在南亚热带地区的杉木人工林改造中,可根据改造目标采用综合评价得分较高的树种组合模式。
AUGUSTO L, RANGER J, BINKLEY D, et al. 2002. Impact of several common tree species of European temperate forests on soil fertility [J]. Annals of Forest Science, 59(3): 233-253.
BALVANERA P, PFISTERER A B, BUCHMANN N, et al. 2006. Quantifying the evidence for biodiversity effects on ecosystem functioning and services [J]. Ecology letters, 9(10): 1146-1156.
BINKLEY D. 1995. The Influence of Tree Species on Forest Soils: Processes and Patterns [C]//Proceedings of the trees and soil workshop,7: 1-33.
CHAPIN III F S, MATSON P A, MOONEY H A. 2002. Principles of Terrestrial Ecosystem Ecology [M]. New York: Springer-Verlag New York.
FINÉR L, MANNERKOSKI H, PIIRAINEN S, et al. 2003. Carbon and nitrogen pools in an old-growth, Norway spruce mixed forest in eastern Finland and changes associated with clear-cutting [J]. Forest Ecology and Management, 174(1/3):51-63.
FRIDLEY J D. 2002. Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities [J]. Oecologia, 132(2): 271-277.
GÄRTNER S, REIF A. 2004. The impact of forest transformation on stand structure and ground vegetation in the southern Black Forest, Germany [J]. Plant and soil, 264(1-2): 35-51.
GETZIN S, DEAN C, HE F, et al. 2006. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island [J]. Ecography, 29(5): 671-682.
HOOPER D U, CHAPIN III F S, EWEL J J, et al. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge [J]. Ecological monographs, 75(1): 3-35.
KELTY M J. 2006. The role of species mixtures in plantation forestry [J]. Forest Ecology and Manage, 233(2): 195-204.
KNOKE T, AMMER C, STIMM B, et al. 2008. Admixing broadleaved to coniferous tree species: a review on yield, ecological stability and economics [J]. European Journal of Forest Research, 127(2): 89-101.
MENG J, LU Y, ZENG J. 2014. Transformation of a degraded Pinus massoniana plantation into a mixed-species irregular forest: Impacts on stand structure and growth in southern China [J]. Forests, 5(12): 3199-3221.
PAQUETTE A, MESSIER C. 2011. The effect of biodiversity on tree productivity: from temperate to boreal forests [J]. Global Ecology and Biogeography, 20(1): 170-180.
SYMSTAD A J, CHAPIN IIIF S, WALL D H, et al. 2003. Long-term and large-scale perspectives on the relationship between biodiversity and ecosystem functioning [J]. Bioscience, 53(1): 89-98.
THOMPSON K, ASKEW A P, GRIME J P, et al. 2005. Biodiversity,ecosystem function and plant traits in mature and immature plant communities [J]. Functional Ecology, 19(2): 355-358.
TILMAN D. 1999. The ecological consequences of changes in biodiversity: A search for general principles [J]. Ecology, 80(5): 1455-1474.
曾小平, 蔡锡安, 赵平, 等. 2008. 南亚热带丘陵3种人工林群落的生物量及净初级生产力[J]. 北京林业大学学报, 30(6): 148-152.
陈龙池, 汪思龙, 陈楚莹. 2004. 杉木人工林衰退机理探讨[J]. 应用生态.
学报, 15(10): 1953-1957.
郭琦, 王新杰. 2014. 不同混交模式杉木人工林林下植被生物量与土壤物理性质研究[J]. 中南林业科技大学学报, 34(5): 70-74.
何友均, 梁星云, 覃林, 等. 2013a. 南亚热带马尾松红椎人工林群落结构、物种多样性及基于自然的森林经营[J]. 林业科学, 49(4): 24-33.
何友均, 梁星云, 覃林, 等. 2013b. 南亚热带人工针叶纯林近自然改造.
早期对群落特征和土壤性质的影响[J]. 生态学报, 33(8): 2484-2495.侯文. 2006. 对应用主成分法进行综合评价的探讨[J]. 数理统计与管理,25(2): 211-214.
黄宇, 冯宗炜, 汪思龙, 等. 2005. 杉木、火力楠纯林及其混交林生态系统C、N贮量[J]. 生态学报, 25(12): 3146-3154.
康冰, 刘世荣, 蔡道雄, 等. 2010. 南亚热带不同植被恢复模式下土壤理化性质[J]. 应用生态学报, 21(10): 2479-2486.
李德军,莫江明,方运霆,等. 2004. 鼎湖山自然保护区不同演替阶段.
森林土壤中有效微量元素状况研究[J]. 广西植物, 24(6): 529-534.李国平, 薛立, 冼干标, 等. 2007. 阔叶幼林取代杉木林后的土壤肥力研究[J]. 水土保持通报, 27(6): 75-79.
李静鹏, 徐明锋, 苏志尧, 等. 2013. 小尺度林分碳密度与碳储量研究[J].华南农业大学学报, 34(2): 213-218.
林德喜, 樊后保, 苏兵强, 等. 2004. 马尾松林下套种阔叶树土壤理化性质的研究[J]. 土壤学报, 41(4): 655-659.
刘恩, 王晖, 刘世荣. 2012. 南亚热带不同林龄红锥人工林碳贮量与碳固定特征[J]. 应用生态学报, 23(2): 335-340.
刘庆, 尹华军, 程新颖, 等. 2010. 中国人工林生态系统的可持续更新问题与对策[J]. 世界林业研究, 23(1): 71-75.
刘世荣, 温远光 2005. 杉木生产力生态学[M]. 北京: 气象出版社: 1-3.
罗应华, 孙冬婧, 林建勇, 等. 2013. 马尾松人工林近自然化改造对植物自然更新及物种多样性的影响[J]. 生态学报, 33(19): 6154-6162.
吕海龙, 董希斌. 2011. 基于主成分分析的小兴安岭低质林不同皆伐改造模式评价[J]. 林业科学, 47(2): 172-178.
马克平, 刘玉明. 1994. 生物群落多样性的测度方法 I. a多样性的测度方法(下)[J]. 生物多样性, 2(4): 231-239.
明安刚, 贾宏炎, 陶怡, 等. 2012. 桂西南 28年生米老排人工林生物量及其分配特征[J]. 生态学杂志, 31(5): 1050-1056.
田大伦, 沈燕, 康文星, 等. 2011. 连栽第1和第2代杉木人工林养分循环的比较[J]. 生态学报, 31(17): 5025-5032.
王卫霞, 史作民, 罗达, 等. 2013. 我国南亚热带几种人工林生态系统碳氮储量[J]. 生态学报, 33(3): 925-933.
薛立, 薛晔, 列淦文, 等. 2012. 不同坡位杉木林土壤碳储量研究[J]. 水土保持通报, 32(6): 43-46.
薛立,向文静,何跃君,等. 2005. 不同林地清理方式对杉木林土壤肥力的影响[J]. 应用生态学报, 16(8): 1417-1421.
杨昆, 管东生. 2007. 珠江三角洲地区森林生物量及其动态[J]. 应用生态学报, 18(4): 707-712.
姚迎九, 康文星, 田大伦. 2003. 18年生樟树人工林生物量的结构与分布[J]. 中南林学院学报, 23(1): 1-5.
俞月凤, 宋同清, 曾馥平, 等. 2013. 杉木人工林生物量及其分配的动态变化[J]. 生态学杂志, 32(7): 1660-1666.
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522.
庄雪影, 洪文君, 黄川腾, 等. 2012. 11种乡土阔叶树在广州南沙的早期生长表现[J]. 林业科技开发, 26(5): 17-21.
Evaluation of Different Tree Species Composition Patterns in Stand Conversion of Cunninghamia lanceolata in Southern Subtropical Region of China
HUANG Yuhui1, 2, ZHANG Weiqiang1, 2, GAN Xianhua1, 2, TANG Honghui1, 2, PAN Lijun3, XIAN Ganbiao3
1. Guangdong Provincial Key Laboratory of Bio-control for the Forest Disease and Pest, Guangzhou 510520, China;2. Guangdong Academy of Forestry, Guangzhou 510520, China; 3. Maintenance Center for Yunyong Ecological Forest of Foshan, Foshan 528518, China
Stand conversion of Cunninghamia lanceolata plantations plays an important role in forestry in China, which aims at changing Cunninghamia lanceolata plantations into needle and broadleaved mixed forests with complex structure and enhanced ecological function. In this study, needle and broadleaved mixed plantations composed of different tree species were selected to compare the soil physical and chemical properties, carbon density and biodiversity. The patterns of different tree species composition were evaluated by the principal component analysis (PCA). The results showed that there were significant differences of soil physicochemical properties among different patterns. The carbon density of different mixed plantations varied from 110.01 to 183.24 t·hm-2. Carbon was mainly stored in soil layer (70.1%~85.7%), followed by the vegetation layer (11.5%~25.8%) and the litter layer (0.8%~5.2%). More than 85% carbon of vegetation was stored in arbor layer. The biodiversity of different mixed plantations apparently differed from each other, with highest difference for species richness index. By PCA, five principal components which were extracted from 26 indices explained 92.7% variations of the tree species composition patterns. Based on the comprehensive evaluation, three patterns of tree species composition: (1) Cunninghamia lanceolata + Cinnamomum burmanni + Mytilaria laosensis + Castanopsis hystrix + Michelia macclurei; (2) Cunninghamia lanceolata + Mytilaria laosensis + C. burmanni + Elaeocarpus sylvestris + Liquidambar formosana; And (3) Cunninghamia lanceolata + Rhodoleia championii + Erythrina variegata + Spathodea campanulata + Mangletia glauca were suitable for stand conversion of Cunninghamia lanceolata plantation in this region. This study could be referenced by stand conversion of inefficient needle plantations, forest management and evaluation of ecosystem services function in south subtropics of China.
Cunninghamia lanceolata; stand conversion; species composition; principal component analysis; comprehensive evaluation; south subtropics
10.16258/j.cnki.1674-5906.2016.06.007
X173; S718.5
A
1674-5906(2016)06-0956-09
国家林业公益性行业科研专项(201404301);广东省林业科技创新专项项目(2015KJCX027);广东省林业科技创新专项项目(2015KJCX029);广东省省级生态公益林效益补偿专项资金项目“广东森林生态环境监测及服务功能评估”
黄钰辉(1981年生),男,助理研究员,博士,研究方向为森林生态学。E-mail: huangyh@sinogaf.cn
张卫强(1976年生),男,研究员,博士,主要从事森林水文与植物生理生态。E-mail: happyzwq@sina.com
2016-04-18
