贝加尔针茅草原羊草光合特征对氮沉降的响应
2016-09-19刘红梅李洁陈新微杨殿林皇甫超河农业部环境保护科研监测所天津300191
刘红梅,李洁,陈新微,杨殿林,皇甫超河农业部环境保护科研监测所,天津 300191
贝加尔针茅草原羊草光合特征对氮沉降的响应
刘红梅,李洁,陈新微,杨殿林,皇甫超河
农业部环境保护科研监测所,天津 300191
以贝加尔针茅草甸草原优势种羊草(Leymus chinensis)为研究对象,通过设置氮沉降量分别为0 kg·hm-2(CK)、30 kg·hm-2(N30)、50 kg·hm-2(N50)、100 kg·hm-2(N100)、150 kg·hm-2(N150)的5个处理进行连续6 a(2010—2015年)模拟氮沉降的野外控制试验,测定羊草光合特性和叶片功能特性对氮沉降的响应。结果表明:N30、N50、N100、N150处理羊草的光饱和点(LSP)分别比CK处理低40.32%、19.73%、38.18%、37.75%,差异达显著水平(P<0.05)。N30、N50、N100、N150处理羊草的最大净光合速率(Pnmax)分别比CK处理低22.51%、48.56%、27.43、50.72%,差异达显著水平(P<0.05)。N30、N50、N100、N150处理羊草叶片N含量(Nmass)、叶片P含量(Pmass)和建成成本(CCmass)均显著高于CK。N30、N50、N100和N150光和能量利用效率(PEUE)和光合氮利用效率(PNUE)均显著低于CK。5种氮素处理羊草Pnmax与PNUE、PEUE呈极显著正相关,叶片Nmass与比叶面积(SLA)、Pmass、CCmass呈极显著正相关, CCmass与SLA之间呈极显著正相关。综合研究表明,氮沉降持续增加会降低羊草的PEUE和PNUE,提高叶片CCmass和叶片N/P比值。
氮沉降;贝加尔针茅草原;羊草;光合特性;叶片功能特性
引用格式:刘红梅, 李洁, 陈新微, 杨殿林, 皇甫超河. 贝加尔针茅草原羊草光合特征对氮沉降的响应[J]. 生态环境学报,2016, 25(6): 973-980.
LIU Hongmei, LI Jie, CHEN Xinwei, YANG Dianlin, HUANGFU Chaohe. Photosynthetic Characteristics of Leymus chinensis in Response to Simulated Nitrogen Deposition in Inner Mongolia, China [J]. Ecology and Environmental Sciences, 2016, 25(6): 973-980.
高氮沉降作为全球变化的重要的现象之一,其所带来的一系列生态问题日益严重,影响了陆地生态系统的结构和功能,成为近年来国内外生态学家关注的热点问题之一(Janssens et al.,2010;Maskell et al.,2010;何亚婷等,2010)。在全球范围内,高氮沉降的增加呈现出全球化的趋势(Throop,2005;Galloway et al.,2004)。在工业发达的欧洲和北美,高氮沉降对草原生态系统健康造成的影响已严重威胁到草原生物多样性和生态系统功能(Maston et al.,2002)。我国成为继北美和欧洲之后第三大氮沉降集中区域(Mo et al.,2008),自1961年到2008年氮沉降量增加了59%(Lu et al.,2014)。大量而持续的氮沉降对草地生态系统的组成及功能必将造成深远的影响。研究和预测氮沉降对天然草地生态系统的影响,并通过草原生态系统的科学管理,充分利用目前及未来氮素输入的可能变化成为当前亟待解决的重要科学问题。制定科学的草地生态系统管理对策,对于实现天然草地的可持续发展具有重要的理论和实践意义。
光合作用是植物对环境变化最敏感的生理过程之一。叶是植物通过光合作用获取能源和合成光合产物的主要器官,叶片的结构性状一定程度上决定了叶片光合生理活性及其对资源的利用效率。养分保持是植物对贫瘠生境的适应策略之一,较高的养分保持能力说明植物具有较强的适应能力(Aerts et al.,2000)。比叶面积(Specific leaf area,SLA)和叶片养分浓度是衡量植物养分保持能力的两个主要叶片功能特性。比叶面积反映了植物获取环境资源的能力(黄菊莹等,2009;Wright et al.,2004)。叶片养分浓度反映了植物对土壤养分的吸收特性(Grime et al.,1997)。养分保持能力强的植物,通常具有较低的叶片养分浓度、比叶面积和光合作用能力(Wright et al.,2004;Reich et al.,1997;Wright et al.,2003)。氮素是植物体内氨基酸、蛋白质、核酸、辅酶以及光合色素分子等的主要组成,叶片含氮量与光合速率密切相关(郑淑霞等,2007)。氮含量较高的叶片通常具有较高的最大光合速率。但是,过度施氮将可能引起叶片产生碳限制,导致CO2同化速率降低(许振桂等,2004;Nakaji et al.,2001)。研究不同氮沉降水平对植物光合特性的影响,可以加深对植物生理特性、生态适应、生产潜能和光能利用效率的认识。
贝加尔针茅(Stipa baicalensis)草原是欧亚大陆草原的重要组成部分,是内蒙古草甸草原的代表类型之一,在我国畜牧业生产中占有重要的地位(赵帅等,2011)。本研究经过连续6年模拟氮沉降野外控制试验,选取贝加尔针茅草原优势种羊草(Leymus chinensis)作为研究对象,进行光合-光强响应(Pn-PAR)测定,通过模型拟合得出最大净光合速率(maximum net photosynthetic rate,Pnmax)、光饱和点(light saturation point,LSP)、光补偿点(light compensation point,LCP)和表观量子效率(apparent quantum yield,AQY)等光合生理生态特性,并比较分析不同氮沉降处理下羊草光合特性及叶片比叶面积(specific leaf area,SLA)、氮含量(leaf N content,Nmass)、磷含量(leaf P content,Pmass)、建成成本(leaf construction cost,CCmass)、光合氮利用效率(photosynthetic nitrogen use efficiency,PNUE)及光合能量利用效率(photosynthetic energy use efficiency,PEUE)等的差异,并综合分析不同氮沉降量处理下羊草植物光合特性和叶片特性之间的相关性,探讨典型草原生态系统中植物光合特性变化、叶片功能性状对氮沉降水平改变的响应机制,以期为天然草地优化管理提供参考依据
1 研究区域概况与实验方法
1.1研究区域概况
研究区域位于大兴安岭西麓,内蒙古自治区鄂温克自治旗伊敏苏木境内,地理位置为北纬48°27′~48°35′,东经 119°35′~119°41′,海拔高度为760~770 m,地势平坦,属于温带草甸草原区。半干旱大陆性季风气候,年均气温-1.6 ℃,年降水量328.7 mm,年蒸发量 1478.8 mm,≥0 ℃年积温2567.5 ℃,年均风速4 m·s-1,无霜期113 d。土壤类型为暗栗钙土(Zhang et al.,2008)。试验开始时的土壤基础理化性质为:土壤pH 7.07,有机C 27.92 g·kg-1,全N 1.85 g·kg-1,全P 0.45 g·kg-1。
植被类型为贝加尔针茅草甸草原,贝加尔针茅为建群种,羊草为优势种,草地麻花头(Serratulay amatsutanna)、日荫菅(Carex pediformis)、变蒿(Artemisia commutata)、扁蓿豆(Melissitus ruthenica)、多茎野豌豆(Vicia multicaulis)、祁州漏芦(Rhaponticum uniflorum)、寸草苔(Carex duriuscula)、肾叶唐松草(Thaictrum petaloideum)等为常见种或伴生种。共有植物66种,分属21科49属。
1.2研究方法
1.2.1样地设置
试验开始于2010年6月,在围栏样地内设置养分添加试验,氮素添加处理设5个水平,依次为:0 kg·hm-2(CK)、30 kg·hm-2(N30)、50 kg·hm-2(N50)、100 kg·hm-2(N100)、150 kg·hm-2(N150),每年分2次施入。第1次6月15日施氮50%处理水平;第2次7月15日施氮50%处理水平,氮素为NH4NO3。根据氮处理水平,将每个小区每次所需要施加的NH4NO3溶解在8 L水中(全年增加的水量相当于新增降水1.0 mm),完全溶解后均匀喷施到各个小区内。CK小区同时喷洒等量的水。共5个处理小区,6次重复,小区面积8 m×8 m,小区间设2 m隔离带,重复间设5 m隔离带。
1.2.2样品采集与测定
于2015年8月10日开展野外光合实验测定和样品采集。此时羊草已经完成叶片形态建成。光合-光强(Pn-PAR)响应测定:于上午9:00—11:00采用Li-6400便携式光合测定仪(Li-Cor Inc.,Lincoln,NE,USA)对羊草的净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)等指标进行测定(邓钰等,2012)。叶室温度设为 25 ℃,CO2浓度控制在 400 μmol·mol-1,设置有效光合辐射梯度为2000、1500、1000、800、500、300、100、50和0 μmol·m-2·s-1,利用自动测量程序进行光合-光强响应的测定(朱慧等,2009)。选取完全展开的成熟叶片进行测定,测定时保持叶片自然生长角度不变,每片叶测3次,每区组重复测定5次。叶片比叶面积测定:采用 Li-3100A叶面积仪(Li-Cor,Lincoln,Nebraska,USA)测定叶面积。将测定完叶面积的叶片单独装在自封袋中,带回实验室烘干至恒重,测定叶片干重。按照以下公式计算叶面积:
SLA=ULA/DY(1)
式中,SLA为叶片比叶面积,单位为cm2·g-1;ULA为单位叶片面积,单位为cm2;DY为叶片干重,单位为g。
土壤和叶片样品的采集和测定(鲍士旦,2000):用土壤采样器在各个处理小区内按照 S型取样法选取10个点,去除表面植被,取0~10 cm土壤混匀,去除根系和土壤入侵物,采用四分法留取1 kg土壤,迅速装入无菌封口袋,将其分成两部分,一部分于-20 ℃超低温冰箱中保存,用于土壤速效养分分析,一部分土壤样品自然风干后,研磨,分别过100目和20目孔筛并保存,用于土壤理化性质分析。在每个小区内随机选取生长相对一致的具有代表性的植株各 15株,用自封袋保存于低温冰盒中。植物样品105 ℃杀青30 min,然后65 ℃(72 h)烘干至恒重,叶片粉碎过100目筛,混匀后保存在塑封袋中以备分析。土壤有机C测定采用水合热重铬酸钾氧化-比色法,土壤NH4+-N和NO3--N含量采用流动注射分析仪(QC8000,Lachat,USA)测定,植物和土壤全N用凯氏定氮法,全P用钼锑抗比色法测定。
叶片灰分浓度(Ash)的测定:在马弗炉中500 ℃灼烧6 h,剩余残渣称重。
干重热值采用氧弹式热量计(HWR-15E,上海上立检测仪器厂)测定:取0.5 g左右植物样品粉末,经压片,完全燃烧测定热值,每个样品分别测定3个重复,取平均值作为干重热值,测定前用苯甲酸标定(Williams et al.,1987)。

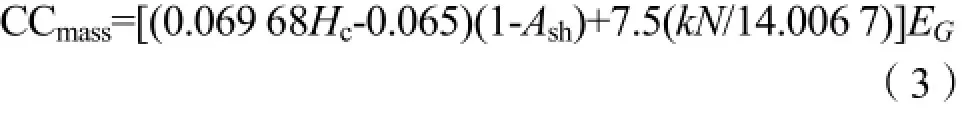
式中,Hc为去灰分热值,单位为kJ·g-1;Ash为灰分浓度,单位为%;CV为干重热值,单位为kJ·g-1;CCmass为叶片单位质量建成成本,单位为gglucose·g-1;N为有机氮质量分数,单位为mg·g-1;EG为生长效率;k为N的价数(若为NO3-,k=5,若为 NH4+,k=-3)。不同物种的生长效率为 0.87 (Penningde et al.,1974),对于每个样品,先以NH4+和NO3-作为N的存在形式分别计算,再按照土壤中NH4+-N和NO3--N的比例,求加权平均值作为叶片CCmass。式中,Narea为叶片单位面积氮含量,单位为g·m-2;PEUE为光合能量利用效率,单位为µmol·gglucose-1·s-1;PNUE为光合氮利用效率,单位为µmol·g-1·s-1。




1.3数据分析
光合-光强响应曲线采用Ye(2007)、叶子飘等(2008)提出的直角双曲线修正模型进行拟合,模型方程为:

式中,α是光响应曲线的初始斜率,即表观量子效率(AQY);β和γ为系数,单位为m2·s·μmol-1;I为光合有效辐射,单位为μmol·m-2·s-1;Rd为暗呼吸速率,单位为mg· g-1· h-1。
水分利用率(water use efficiency,WUE)的计算公式为:

式中,Tr为蒸腾速率,单位为mmol·m-2·s-1。
应用SPSS 16.0进行统计分析,采用单因素方差分析(one-way ANOVA)和最小显著差数法(LSD法)进行不同处理间均值的方差分析和差异显著性比较(P=0.05)。叶片 Nmass与植物 SLA、CCmass、Pn之间的关系采用Pearson相关分析,并进行双尾显著性检验。
2 结果与分析
2.1不同氮素添加下羊草光合作用对有效光合辐射的响应
随着有效光合辐射的增强,5种氮素添加处理下羊草的Pn均呈先增加后降低的趋势,当光照强度在0~300 μmol·m-2·s-1范围内时5种氮素处理的Pn几乎呈线性增长(图1)。当达到一定光合辐射强度时,5种氮处理的Pn达到最大值,即为光饱和点。CK(R2=0.864,P<0.01)、N30(R2=0.716,P<0.05)、N50(R2=0.802,P<0.01)、N100(R2=0.748,P<0.05)、N150(R2=0.837,P<0.01)处理的Pn与有效光合辐射的相关性均达显著水平。由直角双曲线的修正模型方程可以计算出5种植物的光合响应参数(表1)。由表1可知,5种氮素处理羊草的LSP均高于900 μmol·m-2·s-1,其大小顺序为CK>N50>N150>N100>N30,N30、N50、N100、N150处理的LSP分别比 CK处理低 40.32%、19.73%、38.18%、37.75%,差异达显著水平(P<0.05)。LCP随着氮添加水平的增加而增大,大小顺序为N150>N100>N50>N30>CK,N30、N50、N100、N150处理的LCP分别比CK处理高11.34%、12.43%、13.77%、22.18%,差异达显著水平(P<0.05)。AQY随着氮添加水平的增加表现为先升高后降低的趋势,N100处理的AQY显著高于其它4种氮素处理。5个处理Pnmax的大小顺序为 CK>N30>N100>N50>N150,N30、N50、N100、N150处理的Pnmax分别比 CK处理低22.51%、48.56%、27.43、50.72%,差异达显著水平(P<0.05)。
气孔是植物叶片与外界进行气体交换的通道,是叶片蒸腾及光合原料CO2进入细胞内的通道,其行为与叶片的蒸腾和光合性能有着密切的关系,陆生植物通过气孔开放调节水分的出入(Farquhar et al.,1982)。水分利用效率是指消耗单位重量的水,植物所固定的CO2,是衡量植物水分消耗与物质生产之间关系的重要综合性指标。由图1可以看出,随着有效光合辐射的增加,5种氮素处理羊草的Gs、Tr和WUE均有一定程度的增加。在有效光合辐射大于1500 μmol·m-2·s-1时,N50和N100处理的Gs随着光照强度的增加而下降,CK、N30、N150处理Gs随着光照强度增加而增加。CK处理的Gs随有效光合辐射增加而增加的幅度明显高于其它4种氮素处理。5种氮素处理羊草的Tr均随着有效光合辐射的增加而增加。在模拟光辐射逐渐增强初期,各处理羊草的WUE快速升高,当模拟光辐射达到800 μmol·m-2·s-1时,各处理羊草WUE达到最大值;此后,随着光强的增加,WUE则缓慢下降。CK处理的WUE随着有效光合辐射增加而增加的幅度明显高于其它4种氮素处理,说明CK处理羊草对水分的利用效率更高。

图1 不同氮素添加下羊草光合-光响应曲线的比较Fig. 1 Comparison of Pn-PAR curve of Leymus chinensis under different nitrogen addition
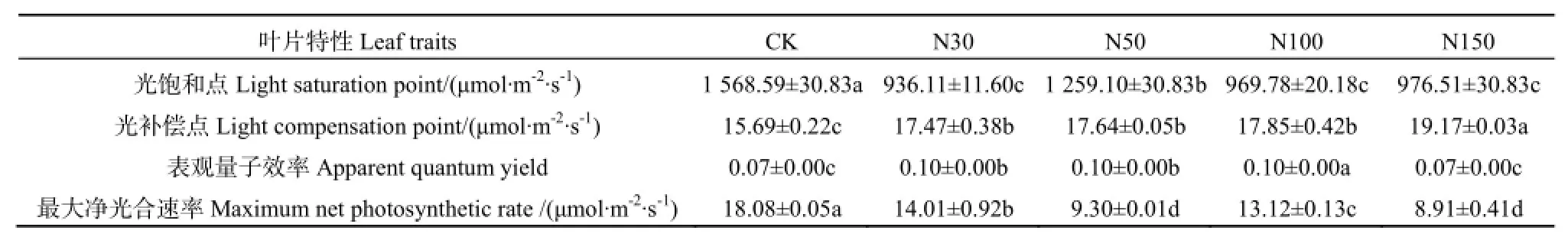
表1 不同氮素添加下羊草的光合响应特征参数Table 1 Photosynthetic parameters of Leymus chinensis under different nitrogen addition
2.2不同氮素添加下羊草叶片特性的比较
由表2可以看出,羊草叶片SLA随着氮素添加量的增大表现为先升高后降低的趋势,N50、N100 和N150处理的叶片SLA显著高于CK,N30处理的SLA高于CK,但无显著差异。叶N含量随着氮素添加量的增大而增加,N30、N50、N100和N150处理的叶N含量分别比CK 高21.95%、36.04%、47.45%和51.52%,差异达显著水平(P<0.05)。叶P含量无一致的变化规律,N30、N50、N100和N150处理的叶P量均显著高于CK。叶N/P、CCmass随着氮素添加量的增大而增大。叶片 Ash随着氮素添加量的增大而减小,N30、N50、N100和N150处理的叶Ash显著低于CK。5种氮素处理的叶片Hc之间没有显著差异。PEUE、PNUE没有一致的变化规律,N30、N50、N100和N150的PEUE和PNUE均显著低于CK。2.3不同氮素添加下Pnmax与叶片Nmass、Pmass、SLA、CCmass、PEUE、PNUE之间的相关性

表2 不同氮素添加下羊草叶片特性的比较Table 2 Comparison of leaf traits of Leymus chinensis under different nitrogen addition
由表 3可以看出,5种氮素添加下羊草 Pnmax与PNUE、PEUE呈极显著正相关(P<0.01),与叶片 Nmass、Pmass、SLA 和 CCmass呈极显著负相关(P<0.01);羊草叶片Nmass叶片SLA、Pmass和CCmass呈极显著正相关(P<0.01),与Pnmax呈极显著负相关(P<0.01);叶片CCmass与叶片SLA之间也呈极显著正相关(P<0.01)。
3 讨论
净光合速率 Pn是评估光合作用强弱的重要指标。从图1可以看出,在相同有效光合辐射的情况下,4种添加N处理的羊草净光合速率低于CK,且最大净光合速率显著低于CK。LSP与LCP是反映植物对强光和弱光利用能力的指标。LCP较低,LSP较高的植物对光环境的适应性较强(Yang et al.,2005)。本试验研究表明,对照CK的LCP最低,而LSP最高,说明氮沉降降低了羊草利用弱光的能力,即对光环境的适应性降低。综合分析认为,氮沉降降低了羊草的光合作用能力。这与蒋丽(2012)研究结果一致。而与肖胜生等(2010)、李明月等(2013)研究认为氮素添加对植物光合速率有一定的促进作用的结论不一致。可能是研究的植物种类、施氮年限、生境条件等不同造成的。国内外一些研究认为,适量的氮素添加在一定时间内有利于光合同化和植物生长,但是在过量或高浓度氮素处理下,植物体内营养元素出现失衡,造成植物光合能力降低(Nakaji et al.,2001)。本研究认为施氮对贝加尔针茅草原羊草的光合作用受水分条件的限制和氮素积累的影响,从而导致羊草的光合作用受到限制。

表3 不同氮素添加下羊草Pnmax与叶片Nmass、Pmass、SLA、CCmass、PEUE、PNUE之间的相关性Table 3 Correlation coefficient among Pnmax, Nmass, Pmass, SLA, CCmass, PEUE and PNUE of Leymus chinensis under different nitrogen addition
叶片 SLA是植物调节和控制碳同化最重要的植物特性(Ye,2007;韦兰英等,2008)。当植物可获取的营养状况改变后,植物可以通过改变叶片结构、叶片养分含量浓度来获取充足的光资源(肖胜生等,2010)。本研究表明随着氮沉降水平的升高,叶片SLA表现为先升高后降低的趋势。N100处理的叶片SLA最高,N150处理的叶片SLA低于N100。这与万宏伟等(2008)研究结果一致。叶片N含量随着N添加水平的升高而升高,与万宏伟等(2008)对内蒙古温带草原羊草叶片含氮量在氮添加小于175 kg·hm-2时随氮添加量的增大而增加,宾振钧等(2014)对青藏高原高寒草甸垂穗披碱草叶片含N量随氮添加量增大而增加的研究结果一致。叶片 P含量无一致的变化规律,4 种N添加处理的叶片P含量均显著高于对照CK。N沉降的增加显著提高了植物的 N/P比值,本研究叶片N/P比值随着氮添加水平的升高而增加,羊草中叶片N/P比值从13.02增加到16.63,这与Stevens et al.,(2011)研究结果一致。N沉降提高叶片中N/P比值,其原因可能是N增加会提高土壤有效N浓度,增加了土壤对植物有效N的供应,从而提高了N/P比值;另一种原因可能是随着N的持续增加,土壤出现了酸化(Prietzel et al.,2006),降低了土壤P的矿化速率和有效P含量(Pape et al.,1989),减少了土壤对植物有效 P的供应,叶片中P含量降低,从而提高了叶片的N/P比值。国内外一些研究认为,蛋白质、氨基酸等含氮化合物的合成需要较高的能量,叶片N浓度的增加会提高叶片的呼吸作用,因此叶片N浓度的增加可能提高叶片建成成本。本研究中CCmass随着氮添加量增大而增加,这与Zha et al.(2002)的研究结论一致。
水分和氮素是植物光合作用和生长的必要因子(Novriyanti et al.,2012),水分利用效率和光合氮利用效率是评估植物净光合速率的重要指标。在资源匮乏的环境中,植物会采用资源保护的策略来提高自身的竞争力(Funk et al.,2007)。Lambers et al.(1992)研究认为,在能够支持植物生长的光强下,氮含量较低的植物叶片的PNUE较大,这是因为这些植物合理地将叶片氮分配到光合中,以达到最优的光合氮利用效率。而另一些研究者认为,高氮叶片的Rubisco酶活性降低(Cheng et al.,2000;Ray et al.,2003),高氮叶片下增加的Rubisco酶更多以氮库形式存在,并没有催化能力,即单位Rubisco酶的羧化效率降低(Warren et al.,2003)。在本研究中,CK处理的水分利用效率、光合能量利用效率、光合氮利用效率都显著高于其他4个氮素添加处理。相关性分析发现,叶片 N含量与PEUE、PNUE呈极显著负相关。这与Lambers et al.(1992)的研究结论一致。Pnmax直接反映了植物本身积累干物质的能力,由表 3可知,羊草的 Pnmax与PNUE呈极显著正相关,这与郑淑霞等(2007)的研究一致。Pnmax和PNUE的降低主要是因为植物本身分配到光合作用中的N减少,而不是因为叶片中N含量的减少。PEUE是表征植物叶片能量利用效率的重要指标,羊草的 PEUE与 Pnmax、PNUE均呈极显著正相关(表3),N沉降增加使得羊草Pnmax降低、Nmass增加,进而降低了羊草 PEUE。对于羊草光合特性与环境的关系,研究者大多从土壤水分状况、干旱胁迫以及放牧影响的角度进行研究(邓钰等,2012;米雪等,2015;李林芝等,2009),有关氮沉降对羊草光合作用的影响机理仍有待深入探讨。
4 结论
(1)氮沉降降低了羊草的光合速率、气孔导度、蒸腾速率和水分利用效率。
(2)氮沉降增加了羊草叶片的 Nmass、Pmass、SLA、CCmass、N/P比值,降低了羊草叶片的养分保持能力。
(3)5个模拟氮沉降处理的 Pnmax与 PNUE、PEUE呈极显著正相关;植物叶片Nmass与叶片SLA、CCmass呈极显著正相关。综合以上分析,氮沉降的增加降低了羊草的光合能量利用效率和光合氮利用效率。
AERTS R, CHAPIN III F S. 2000. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns [J]. Advances in Ecological Research, 30(8): 1-67.
CHENG L, FUEHIGAMI L H. 2000. Rubisco activation state decreases with increasing nitrogen content in apple leaves [J]. Journal of Experimental Botany, 51(351): 1687-1694.
FARQUHAR G D, SHARKEY T D. 1982. Stomatal conductance and photosynthesis [J]. Annual review of Plant Physiology, 33(1): 317-345.
FUNK J L, VITOUSEK P M. 2007. Resource-use efficiency and plant invasion in low-resource systems [J]. Nature, 446(7139): 1079-1081.
GALLOWAY J N, DENTENER F J, CAPONE D G, et al. 2004. Nitrogen cycles, past, present, and future [J]. Biogeochemistry, 70(2): 152-226.
GRIME J P, THOMPSON K, HUNT R, et al. 1997. Integrated screening validates primary axes of specialisation in plants [J]. Oikos, 79(2): 259-281.
JANSSENS I A, DIELEMAN W, LUYSSAERT S. 2010. Reduction of forest soil respiration in response to nitrogen deposition [J]. Nature Geoscience, 3(5): 315-322.
LAMBERS H, POORTER H. 1992. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences [J]. Advances in Ecological Research, 23: 187-261.
LU C Q, TIAN H Q. 2014. Half-century nitrogen deposition increase across China: A gridded time-series data set for regional environmental assessments [J]. Atmospheric Environment, 97: 68-74.
MASKELL L C, SMART S M, BULLOCK J M. 2010. Nitrogen deposition causes widespread loss of species richness in British habitats [J]. Global Change Biology, 16(2): 671-679.
MASTON P A, LOHSE K A, HALL S J. 2002. The globalization of nitrogen deposition: consequences for terrestrial ecosystems [J]. Ambio,31(2): 113-119.
MO J M, ZHANG W, ZHU W X. 2008. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China [J]. Global Change Biology, 14(2): 403-412.
NAKAJI T, FUKANI M, DOKAYA Y, et al. 2001. Effects of high nitrogen load on growth, photosynthesis and nutrient status of Cryptomeria japonica and Pinus densijlora seedlings [J]. Trees, 15(8): 453-461.
NOVRIYANTI E, WATANABE M, MAKOTO K, et al. 2012. Photosynthetic nitrogen and water use efficiency of acacia and eucalypt seedlings as afforestation species [J]. Photosynthetica, 50(2): 273-281.
PARE D, BERNIER B. 1989. Phosphorus-fixing potential of Ah and H horizons subjected to acidification [J]. Canadian Journal of Forest Research, 19(1): 132-134.
PENNINGDE VRIES F W T, BRUNSTING A H M, VAN LAAR H H. 1974. Products, requirements and efficiency of biosynthesis: A quantitative approach [J]. Journal of Theoretical Biology, 45(2): 339-377.
PRIETZEL J, STETTER U,KLEMMT H J, et al. 2006. Recent carbon and nitrogen accumulation and acidification in soils of two Scots pine ecosystems in Southern Germany [J]. Plant and Soil, 289(1): 153-170.
RAY D, SHESHSHAYEE M S, MUKHOPADYOOY K, et al. 2003. High nitrogen use efficiency in rice genotypes is associated with higher net photosynthetic rate at lower Rubisco content [J]. Biologia Plantarum,46(2): 251-256.
REICH P B, WALTERS M B, ELLSWORTH D S. 1997. From tropics to tundra: global convergence in plant functioning [J]. Proceedings of the National Academy of Sciences of the United States of America, 94(25): 13730-13734.
STEVENS C J, DUPRE C, DORLAND E, et al. 2011. The impact of nitrogen deposition on acid grasslands in the Atlantic region of Europe [J]. Environmental Pollution, 159(10): 2243-2250.
THROOP H L. 2005. Nitrogen deposition and herbivory affect biomass production and allocation in an annual plant [J]. Oikos, 111(1): 91-100.
WARREN C R, DREYER E, ADAMS M A. 2003. Photosynthesis-Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores [J].Trees, 17(4): 359-366.
WILLIAMS K, PERCIVAL F, MERINO J, et al. 1987. Estimation of tissue construction cost from heat of combustion and organic nitrogen content [J]. Plant, Cell & Environment, 10(9): 725-734.
WRIGHT I J, REICH PB, WESTOBY M, et al. 2004. The worldwide leaf economics spectrum [J]. Nature, 428(6895): 821-827. WRIGHT I J, WESTOBY M. 2003. Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species [J]. Functional Ecology, 17(1): 10-19.
YANG X H, ZOU Q, ZHAO S J. 2005. Photosynthetic characteristics and chlorophyll fluorescence in leaves of cotton plants grown in full light and 40% sunlight [J]. Journal of Plant Ecology, 29(1): 8-15.
YE Z P. 2007. A new model for relationship between light intensity and the rate of photosynthesis in Oryza sativa [J]. Photosynthetica, 45(4): 637-640.
ZHA T S, WANG K Y, RYYPPO A, et al. 2002 Needle dark respiration in relation to within-crown position in Scots pine trees grown in long-term elevation of CO2 concentration and temperature [J]. New Phytologist, 156(1): 33-41.
ZHANG N L, WAN S Q, LI L H. 2008. Impacts of urea N addition on soil microbial community in a semiarid temperate steppe in northern China [J]. Plant and Soil, 311(1/2): 19-28.
鲍士旦. 2000. 土壤农化分析(第三版)[D]. 北京: 中国农业出版社.
宾振钧, 王静静, 张文鹏, 等. 2014. 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响[J]. 植物生态学报, 38(3): 231-237.
邓钰, 柳小妮, 辛晓平, 等. 2012. 不同放牧强度下羊草的光合特性日动态变化[J]. 草业学报, 21(3): 308-313.
何亚婷, 齐玉春, 董云社, 等. 2010. 外源氮输入对草地土壤微生物特性影响的研究进展[J]. 地球科学进展, 25(8): 877-886.
黄菊莹, 袁志友, 李凌浩. 2009. 羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化[J]. 植物生态学报, 33(3): 442-448.
蒋丽. 2012. 松嫩草地光合特性对增温和氮素添加的响应[D]. 长春: 东北师范大学.
李林芝, 张德罡, 辛晓平, 等. 2009. 呼伦贝尔草甸草原不同土壤水分梯度下羊草的光合特性[J]. 生态学报, 29(10): 5271-5279.
李明月, 王健, 王振兴, 等. 2013. 模拟氮沉降条件下木荷幼苗光合特性、生物量与C、N、P分配格局[J]. 生态学报, 33(5): 1569-1577.
米雪, 李晓兵, 王宏, 等. 2015. 内蒙古典型草原不同放牧强度下羊草光合生理生态特性分析[J]. 中国草地学报, 37(3): 92-98.
万宏伟, 杨阳, 白世勤, 等. 2008. 羊草草原群落6种植物叶片功能特性对氮素添加的响应[J]. 植物生态学报, 32(3): 611-621.
韦兰英, 上官周平. 2008. 黄土高原不同退耕年限坡地植物比叶面积与养分含量的关系[J]. 生态学报, 28(6): 2526-2535.
肖胜生, 董云社, 齐玉春, 等. 2010. 内蒙古温带草原羊草叶片功能特性与光合特征对外源氮输入的响应[J]. 环境科学学报, 30(12): 2535-2543.
许振桂, 周广胜. 2004. 植物氮代谢及其环境调节研究进展[J]. 应用生态学报, 15(3): 511-516.
叶子飘, 于强. 2008. 光合作用光响应模型的比较[J]. 植物生态学报,32.
赵帅, 张静妮, 赖欣, 等. 2011. 放牧与围栏内蒙古针茅草原土壤微生物生物量碳、氮变化及微生物群落结构PLFA分析[J]. 农业环境科学学报, 30(6): 1126-1134.
郑淑霞, 上官周平. 2007. 不同功能型植物光合特性及其与叶氮含量比叶重的关系[J]. 生态学报, 27(1): 171-181.
朱慧, 马瑞君. 2009. 入侵植物马缨丹(Lantana camara)及其伴生种的光合特性[J]. 生态学报, 29(5): 2701-2709.
Photosynthetic Characteristics of Leymus chinensis in Response to Simulated Nitrogen Deposition in Inner Mongolia, China
LIU Hongmei, LI Jie, CHEN Xinwei, YANG Dianlin, HUANGFU Chaohe
Agro-Environmental Protection Institute, Ministry of Agriculture, Tianjin 300191, China
Increasing atmospheric nitrogen deposition is one of the hot topics related to global climate change. Studying the effects of nitrogen deposition on morphological and physiological characteristics of plants increasingly becomes a major focus in the field of ecological sciences. We conducted a field manipulation experiment to study the response of Photosynthetic characteristics and leaf functional traits of Leymus chinensis in response to simulated nitrogen deposition in a Stipa baicalensis grassland in inner Mongolia. This study was conducted during 2010—2015, five treatments including 0 kg·hm-2(CK)、30 kg·hm-2(N30)、50 kg·hm-2(N50)、100 kg·hm-2(N100)、150 kg·hm-2(N150)were conducted. Forty-eight (8 m×8 m) plots were established with 2 m belts between treatment areas, respectively. The results showed that the light saturation point (LSP) of N30, N50, N100, N150 significantly lower than that of CK by 40.32, 19.73, 38.18, 37.75 percent, respectively (P<0.05). Maximum net photosynthetic rate (Pnmax) of N30, N50,N100, N150 were significantly lower than that of CK by 22.51, 48.56, 27.43, 50.72 percent, respectively (P<0.05). Leaf N content (Nmass), leaf P content (Pmass) and leaf construction cost (CCmass) of N30, N50, N100, N150 were significantly higher than that of CK. Photosynthetic energy use efficiency (PEUE) and photosynthetic nitrogen use efficiency (PNUE) of N30, N50, N100, N150 were significantly lower than that of CK. Correlation analysis showed that Pnmaxwere positively correlated with PEUE and PNUE (P<0.01), leaf Nmasswere positively correlated with specific leaf area (SLA), Pmassand CCmass(P<0.01), CCmasswere positively correlated with SLA (P<0.01). In summary, nitrogen deposition induced the decrease of PEUE, PNUE of Leymus chinensis, and the increased leaf mass and leaf N/P.
nitrogen deposition; Stipa baicalensis grassland; Leymus chinensis; photosynthetic characteristics; leaf traits
10.16258/j.cnki.1674-5906.2016.06.009
Q945.79; X17
A
1674-5906(2016)06-0973-08
国家自然科学基金项目(31170435)
刘红梅(1976年生),女,副研究员,主要研究方向为生物多样性与生态安全。E-mail: liuhongmei@caas.cn
2016-04-16
