表达小鹅瘟病毒VP2蛋白重组鸭瘟病毒的构建及其生物学特性
2016-09-19华炯钢叶伟成
陈 柳,余 斌,倪 征,华炯钢,叶伟成,云 涛,张 存
(浙江省农业科学院畜牧兽医研究所,杭州 310021)
表达小鹅瘟病毒VP2蛋白重组鸭瘟病毒的构建及其生物学特性
陈 柳,余 斌,倪 征,华炯钢,叶伟成,云 涛,张 存
(浙江省农业科学院畜牧兽医研究所,杭州 310021)
【目的】鸭瘟和小鹅瘟是番鸭和鹅的两种重要传染病,鸭瘟最主要的防治措施是定期接种鸭瘟病毒减毒活疫苗。根据2012年国际病毒分类委员会(ICTV)的报告,DEV被归为疱疹病毒科的α疱疹病毒亚科马立克氏病毒属。疱疹病毒如伪狂犬病毒、马立克氏病毒、火鸡疱疹病毒等已广泛用于病毒活载体的研究,而近几年也有关于鸭瘟病毒(DEV)作为疫苗活载体的报道。为了为免疫防控鸭瘟和小鹅瘟提供新手段,本研究拟在鸭瘟病毒疫苗株感染性克隆的基础上,构建表达小鹅瘟病毒(GPV)主要免疫原蛋白VP2的重组病毒rDEV-VP2,并研究其生物学特性,进而探讨重组病毒rDEV-VP2作为防治DEV和GPV的二联重组活载体疫苗的可能性。【方法】将密码子优化的 GPV VP2基因通过常规基因克隆的方法插入转移载体 pEP-BGH-end,构建含有 GPV VP2表达框pCMV-VP2-BGH-pA的重组表达质粒。在鸭瘟病毒(DEV)疫苗株细菌人工染色体克隆pDEV-EF1的基础上,通过“Red E/T”两步重组法将GPV VP2基因表达框插入到DEV US7和US8基因之间构建了突变体克隆pDEV-VP2。利用磷酸钙法转染鸡胚成纤维细胞(CEFs)拯救获得重组病毒rDEV-VP2和删除Bac质粒序列的rDEV-VP2-Cre,并对重组病毒细胞体外生长曲线、蚀斑大小和VP2蛋白表达情况进行测定。将rDEV-VP2接种番鸭,在不同时间采集血清,采用间接ELISA法检测血清中GPV VP2抗体产生情况。【结果】间接免疫荧光检测和Western blot分析表明,外源蛋白VP2在CEFs细胞成功表达。病毒生长曲线和蚀斑大小测定结果显示,rDEV-VP2在CEFs细胞上的增殖滴度与亲本株相比无显著差异,表明外源基因VP2的插入不影响rDEV重组病毒的增殖。动物试验结果表明,7日龄雏番鸭接种rDEV-VP2可以诱导产生针对GPV VP2的抗体,免疫后3周抗体阳性率为50%(4/8)。【结论】将小鹅瘟病毒的主要免疫原基因VP2插入到DEV疫苗株基因组的US7和US8基因间构建了表达该免疫原性基因的重组鸭瘟病毒细菌人工染色体,继而在鸡胚成纤维细胞(CEFs)上拯救获得了重组病毒rDEV-VP2,病毒细胞生长特性与亲本株基本一致,且能诱导鸭体产生GPV VP2特异性的抗体。该研究为研制DEV-GPV二联重组活载体疫苗奠定了基础。
鸭瘟病毒;鹅细小病毒;VP2;重组病毒
0 引言
【研究意义】鸭瘟(即鸭病毒性肠炎),是鸭、鹅、天鹅的一种急性、热性、败血性传染病。其病原体为鸭瘟病毒(duck plaque virus,DPV),又称鸭肠炎病毒(duck enteritis virus,DEV);小鹅瘟是由鹅细小病毒(goose parvovirus,GPV)引起的雏鹅和雏番鸭的一种急性或亚急性的败血性传染病,死亡率高。目前,已有新型GPV感染樱桃谷鸭的病例出现。迄今为止,接种弱化疫苗仍是预防和控制致死性 DEV和GPV感染的主要途径。与常规弱化疫苗相比,以鸭瘟病毒作为载体表达 GPV抗原的新型重组活载体疫苗不仅可一针防两病,节约成本,也可减少免疫副反应,更便于鉴别诊断。因此,本研究拟在建立的鸭瘟病毒疫苗株反向遗传操作平台的基础上,进一步探索以DEV作为载体表达GPV抗原、研发GPV-DEV二联苗的可能性。【前人研究进展】DEV在分类上属疱疹病毒科,α-疱疹病毒亚科。α-疱疹病毒是一类极具开发前景的病毒活载体,不仅具有基因组大、非必需基因多、能插入外源基因的容量大、遗传稳定、受母源抗体干扰小、体内存活时间较长等优点,而且已成功研制了许多基因缺失疫苗株,是作为病毒活载体极佳的候选之一[1,2]。表达鸡传染性法氏囊病(IBD)病毒VP2蛋白的火鸡疱疹病毒(HVT)活载体疫苗,可以有效防控IBD和马立克氏病,疫苗已经在养禽业得到广泛应用[3]。此外,以HVT、伪狂犬病病毒(PRV)、马立克氏病毒(MDV)、马1型疱疹病毒(EHV- 1)、牛1型疱疹病毒(BHV-1)和牛疱疹病毒4型(BHV-4)等作为载体的重组病毒研究已经非常广泛,这些疱疹病毒成为表达其他病毒性疾病保护性抗原基因比较理想的动物病毒活载体[4-17]。鸭瘟疫苗具有良好的免疫效果和安全性,疫苗生产和应用技术成熟。近年来,随着鸭瘟病毒基因组全序列的解析和反向遗传系统技术的建立[18-20],鸭瘟病毒作为疫苗活载体研究取得了显著的进展。WANG等分别以UL44(gC)基因缺失的鸭瘟病毒强毒株和疫苗株为载体,表达了H5N1禽流感病毒的HA基因[19,21];ZOU等也以鸭瘟病毒疫苗株为载体表达了同源和异源高致病性H5N1禽流感病毒的HA基因,携带HA基因的重组鸭瘟病毒能诱导机体产生应答H5病毒的体液反应和T细胞免疫,且对H5N1和DEV感染产生快速、持久的保护[22];LIU等研究证实,在DEV疫苗株UL41基因内部插入外源基因既不影响病毒复制表型,也不增加鸭瘟病毒疫苗株对鸭的毒力[23];LIU等成功构建了US2基因缺失或gI/gE双基因缺失的重组鸭瘟病毒,并以之为载体表达了鹅H5亚型禽流感病毒的HA基因[25];CHEN等分别以 DEV作为载体表达了鸭坦布苏病毒的主要抗原蛋白,重组病毒皆能诱导机体产生鸭坦布苏病毒的中和抗体[26-28]。【本研究切入点】目前关于以鸭瘟病毒作为载体表达鹅细小病毒抗原基因的研究仍是空白,研制出这两种病毒的重组疫苗对生产实践具有重要的意义。【拟解决的关键问题】本研究拟将GPV的主要保护性抗原基因VP2插入到DEV疫苗株基因组的US7和US8基因间,构建表达该免疫原性基因的重组鸭瘟病毒细菌人工染色体,继而在鸡胚成纤维细胞(CEFs)上拯救获得了重组病毒rDEV-VP2,并对其在CEFs细胞上的生物学特性进行了初步研究,该研究为研制GPV-DEV基因重组活载体疫苗奠定了基础。
1 材料与方法
试验于2013年1月至2014年7月在浙江省农业科学院畜牧兽医研究所完成。
1.1菌株、质粒和病毒株
pEP-BGH-end质粒、鸭瘟病毒感染性克隆质粒pDEV-vac和pDEV-EF1,大肠杆菌菌株GS1783和鸭瘟病毒重组病毒株 rDEV-Cre均由浙江省农科院畜牧兽医研究所构建并保存[20,28]。
1.2主要试剂
快速连接试剂盒、限制性内切酶、DNA凝胶纯化试剂盒,均购于大连宝生物工程有限公司;质粒提取纯化试剂盒购于OMEGA公司;DMEM、胎牛血清均购于 Gibcol BRL公司;磷酸钙转染试剂盒购于Promega公司。GPV VP2多克隆抗体由本实验室制备;HRP标记的山羊抗小鼠IgG和FITC标记山羊抗小鼠IgG购于中杉金桥生物公司。
1.3序列优化、引物设计、及合成
鹅细小病毒 VP2基因序列参考 GenBank (U25749.1),由GenScript公司以禽为宿主进行优化合成。DEV和GPV VP2引物分别参考序列GenBank (EU082088.2)、GPV VP2优化序列设计,由生工公司合成,序列见表1。pDEV vac-in-s和pDEVvac-in-as用于将表达框pCMV-VP2-BGH-pA插入到DEV基因组的 US7和 US8基因之间。下划线部分与pEP-BGH-VP2同源,粗体部分分别与位于US7和US8基因间插入位点上、下游的序列同源。引物Rec-JD-F 和 Rec-JD-R用于外源基因插入 BAC基因组中的验证。
1.4pEP-BGH-VP2质粒的构建
以禽为宿主进行优化合成的GPV VP2基因合成时在其5′和3′端分别引入了KpnI和Not I酶切位点,将片段用 KpnI和 Not I进行双酶切之后插入到pEP-BGH-end相应的酶切位点中。通过PCR和酶切鉴定筛选获得阳性克隆,送Invitrogen生物公司测序。
1.5pDEV-VP2突变体的构建
通过RedE/T重组将Pcmv-VP2-BGH-pA表达框插入到egfp基因CMV启动子替换为EF1的鸭瘟病毒疫苗株BAC克隆pDEV-EF1[28]。获得的Bac质粒命名为pDEV-VP2。以 pEP-BGH-VP2质粒为模板,用引物pDEVvac-in-s和pDEVvac-in-as(序列见表1)扩增长度约为3 913 bp的片段,胶回收后电转化至pDEV-EF1/ GS1783感受态细胞。最后以pDEV-VP2为模板,用引物Rec-JD-F和Rec-JD-R(序列见表1)扩增,获得大小约3 408 bp的片段,胶回收后克隆至pMD18-T载体,选取阳性克隆送至Invitrogen公司测序。
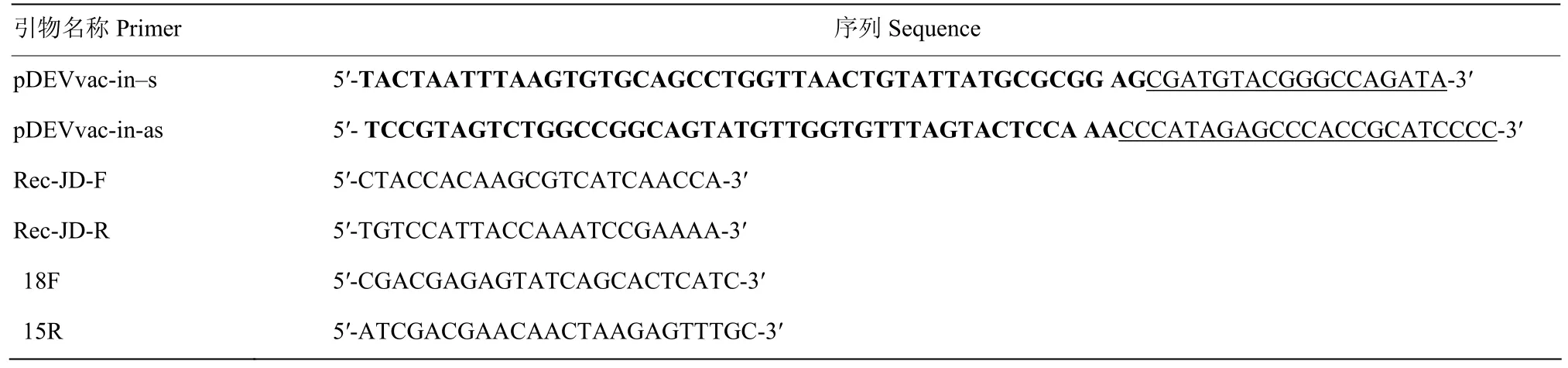
表1 本文所用引物Table 1 Primers used in this study
1.6重组病毒在CEFs细胞中的拯救
用碱裂解法分别提取 pDEV-vac、pDEV-EF1和pDEV-VP2质粒,根据Promega磷酸钙转染试剂盒说明书转染 CEFs,于 37℃ 5%CO2继续培养,待出现70%—80%病变后收取病毒,拯救的病毒分别命名为rDEV-BAC、rDEV-EF1和rDEV-VP2。
为了去除 rDEV-VP2病毒基因组上的 mini-F序列,将4 μg pDEV-VP2和1 μg Cre重组酶表达质粒pCAGGS-NLS/Cre以磷酸钙法共转染至CEFs细胞,第二天铺上含有1.5%甲基纤维素的DMEM培养基,置于CO2培养箱,37℃继续培养,直至出现病毒蚀斑,挑取非荧光蚀斑进行稀释和继代,经过几轮重复,直至获得纯化的非荧光蚀斑病毒。抽提病毒DNA,采用18F/15R引物对病毒进行PCR扩增,鉴定mini-F序列是否去除。鉴定正确的病毒命名为rDEV-VP2-Cre。
1.7重组病毒的细胞增殖特性
1.7.1重组病毒蚀斑大小的测定 将105.5TCID50rDEVBAC、rDEV-EF1和rDEV-VP2病毒冻存液按照10的倍数进行稀释,接种于12孔板中的单层CEFs上,2 h后,换上含有1.5%甲基纤维素的DMEM培养基,放于CO2培养箱,37℃培养48 h,在荧光显微镜下每种病毒各拍100张蚀斑照片,用Image J软件来测量不同病毒的蚀斑面积,计算各病毒的平均值,将rDEV-BAC蚀斑面积设成 100%,其他病毒蚀斑面积以之为标准换算成百分比。
1.7.2病毒生长曲线 以常规病毒学方法测定rDEV-BAC、rDEV-EF1和rDEV-VP2的体外增殖特性。CEFs细胞单层接种0.02MOI病毒后,依次吸附90 min,PBS(pH 7.2)洗涤2次,以冰浴的CBS缓冲液(40 mmol·L-1柠檬酸钠、10 mmol·L-1氯化钾、135 mmol·L-1氯化钠,pH3.0)处理3 min,PBS(pH 7.2)洗涤2次,加入1 mL维持液继续培养。接种后0、12、24、36、48、60、72 h分别收集细胞培养上清和细胞,细胞用PBS(pH 7.2)洗涤2次。收集的上清和细胞冻存于-80℃直至进行滴度测定。分别取100 μL细胞培养上清和细胞裂解液按照常规的方法测定TCID50,每个稀释度重复接种3孔。计算上述各时间点收获的病毒毒价,绘制病毒在体外的多步生长曲线。
1.8GPV VP2蛋白的表达分析
1.8.1间接免疫荧光检测(IFA)蛋白表达 分别接种rDEV-VP2-Cre和对照毒株rDEV-Cre[26]于96孔板的CEFs细胞中,培养72 h,PBS(pH7.0)洗涤3次,用预冷的甲醇/丙酮(1∶1)固定液于-20 ℃固定 30 min,PBS-1%NP40洗涤。一抗采用小鼠抗GPV VP2多克隆抗体(1∶100),于37 ℃湿盒中孵育1 h,洗涤3次之后,滴加伊文斯兰溶液稀释的FITC标记的山羊抗小鼠IgG(1∶100),再次于37 ℃孵育1 h,同样洗涤之后,碱性甘油封片,于Olympus荧光显微镜下观察。
1.8.2Western blot方法检测蛋白表达 蛋白样品于SDS-PAGE电泳完毕,采用半干转印法将蛋白转移至PVDF膜上,取出PVDF膜,放入封闭液中4 ℃封闭过夜。以小鼠抗GPV VP2多克隆抗体(1:500稀释)作为一抗,HRP标记的山羊抗小鼠IgG(1:5 000稀释)作为二抗于37 ℃孵育1 h,最后用DAB显色法进行显色。
1.9动物试验
将24只7日龄无鸭瘟抗体的雏番鸭随机分为3组(n = 8),饲养于不同的房间。分别接种 1×106TCID50rDEV-EF1、1×106TCID50rDEV-VP2,并设立细胞培养物作为阴性对照组。免疫后1、2和3W采血,分离血清,以大肠杆菌 PET28a(+)/BL21系统表达,His-tag凝胶纯化的GPV VP2重组蛋白为一抗、HRP标记的抗禽IgG单克隆抗体为二抗进行间接ELISA,测定VP2抗体水平。
2 结果
2.1pDEV-VP2突变体的鉴定
分别提取pDEV-vac、pDEV-EF1、pDEV-kanVP2 和pDEV-VP2 DNA,经过BamH I和Xhol I酶切鉴定,电泳图谱与预测结果基本一致(图1),且以引物对Rec-JD-F和Rec-JD-R进行PCR扩增片段测序结果也与预期一致,说明外源基因GPV VP2按照预期结果插入。
2.2重组病毒的拯救
磷酸钙转染细胞 48 h后荧光显微镜下观察发现rDEV-BAC、rDEV-EF1和rPRV-VP2孔内均出现荧光噬斑,待细胞80%以上出现荧光时收获病变细胞培养物,即获得拯救病毒(图2-A-C)。
将 pDEV-VP2和 pCAGGS-NLS/Cre共转染至CEFs细胞,经过3轮非荧光蚀斑挑选和PCR鉴定,获得了去除载体序列的病毒rDEV-VP2-Cre(图2-D)。
2.3病毒蚀斑大小测定
rDEV-BAC、rDEV-EF1和rDEV-VP2感染细胞2 d后,各拍摄病毒蚀斑照片100个,并用ImageJ软件测量各个蚀斑面积并计算平均值。将 rDEV-VP2蚀斑面积分别与rDEV-BAC和rDEV-EF1进行比较,发现rDEV-VP2蚀斑面积较rDEV-BAC增加了0.3% (P = 0.119),较rDEV-EF1减少了12.2%(P =0.061)(图3),差异不显著,表明VP2基因的插入,对鸭瘟病毒细胞间感染能力无显著影响。

图1 重组克隆pDEV-VP2的BamH I和Xhol I酶切鉴定Fig.1 Indentification of pDEV-VP2 by BamH I and Xhol I digestion

图2 拯救重组病毒Fig.2 Rescued recombinant viruses (100×)
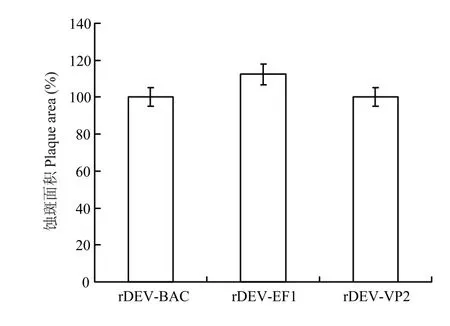
图3 rDEV-BAC、rDEV-EF1和rDEV-VP2在CEFs上的蚀斑面积测定及比较Fig.3 Plaque area measurement of rDEV-BAC,rDEV-EF1 and rDEV-VP2 on CEFs
2.4重组病毒的体外增殖特性
测定重组病毒 rDEV-VP2的生长曲线,并与rDEV-BAC和rDEV-EF1进行比较,结果如图4所示,rDEV-VP2在细胞内的滴度从12 h到72 h稳步增加,并于60 h达到最高,12 h到 48 h之内病毒滴度较rDEV-EF1和rDEV-BAC稍有降低,而60 h时高于后两者。对36 h和60 h病毒滴度进行统计学分析,结果表明rDEV-VP2较rDEV-BAC(P = 0.105,P = 0.100)和rDEV-EF1(P = 0.100,P = 0.080)的差异无统计学意义。rDEV-VP2病毒在上清中的滴度,从12 h到72 h稳步增加,与对照rDEV-BAC和rDEV-EF1无明显差异,统计学分析表明该结论具有统计学意义(P< 0.05)。
图4 rDEV-BAC、rDEV-EF1和rDEV-VP2在CEFs细胞上的体外生长曲线Fig.4 Multi-step growth curves of rDEV-BAC,rDEV-EF1 and rDEV-VP2
2.5外源蛋白VP2表达分析
将rDEV-VP2-Cre和对照rDEV-Cre感染CEFs,以鼠抗GPV VP2多抗为一抗,FITC标记羊抗鼠IgG为二抗进行间接免疫荧光检测,结果表明rDEV-VP2-Cre感染细胞中检测出特异的绿色荧光(图5 A),而对照病毒感染细胞呈阴性(图5 B),表明VP2蛋白在重组病毒感染细胞中获得了表达。
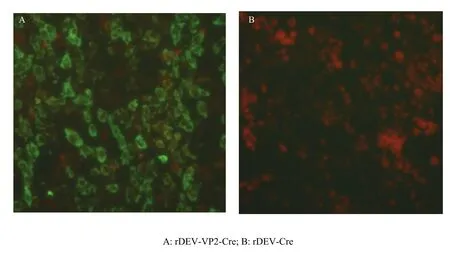
图5 IFA检测重组病毒rDEV-VP2-Cre感染细胞中蛋白表达Fig.5 Detection of VP2 expression in rDEV-VP2-Cre -infected cells by IFA
Western blot检测结果显示rDEV-VP2和rDEVVP2-Cre感染的细胞样品较rDEV-EF1感染的细胞样品在72 kD处有一条特异性条带,与预期蛋白大小一致(图6),说明VP2蛋白获得了有效表达。
2.6重组病毒接种鸭抗体生成情况
为了明确重组病毒rDEV-VP2感染鸭是否能产生针对VP2蛋白的抗体,3组7日龄雏番鸭均于免疫后1W、2 W和3 W采血检测GPV VP2抗体,rDEV-VP2重组病毒免疫抗体阳转率分别为1/8、3/8 和4/8,而rDEV-EF1组和细胞培养液组VP2抗体均为阴性。

图6 Western blot检测病毒感染细胞中VP2的表达Fig.6 Detection of VP2 expression in rDEV-VP2 -infected cells by Western blot
3 讨论
鸭瘟病毒和鹅细小病毒(GPV)可以引起番鸭、鹅小鹅瘟、鸭和半番鸭“喙萎缩综合征”,给养禽业造成了严重的危害。VP2是GPV的主要结构蛋白和保护性抗原,是目前GPV疫苗亚单位疫苗研究的主要候选基因之一[29]。因此,用鸭瘟病毒疫苗株为活载体来表达GPV VP2基因,构建rDEV-VP2研制小鹅瘟-鸭瘟活载体疫苗具有重要实际应用价值。
构建表达载体存在两方面的难题:一是将目的基因插入到病毒载体基因组;二是插入基因是否可以在细胞内获得表达。本文是在鸭瘟病毒疫苗株全基因组感染性克隆基础上,采用“Red E/T”两步重组法对基因组进行基因替换、外源基因插入等序列修饰,该技术不需要特定的酶切位点,仅需35—50 bp碱基序列作为同源臂,从而实现PCR产物或寡核苷酸可以直接作为供体分子对目标DNA进行打靶修饰。“Red E/T”两步重组技术不仅简化了同源重组的操作过程,也提高了重组的效率,同时,因为同源臂不受碱基序列的组成限制,可以人为随意选择,所以运用该技术可以方便地对各种DNA分子尤其是基因组或染色体大分子进行修饰。该技术不仅操作简单、省时、省力,而且可以精准地在全长感染性克隆的基础上进行序列修饰[30-31]。
rDEV-VP2与rDEV-BAC毒株相比,其蚀斑面积和生长曲线皆无显著差异,说明外源基因VP2的插入不影响DEV病毒在CEFs细胞上的增殖特性。本试验中,IFA和Western blot结果皆表明VP2获得了表达,但Western blot分析显示VP2蛋白分子量约72 kD。根据ORF推测,GPV VP2分子量为61 kD,但李茂祥等的研究表明VP2分子量位于60—72 kD之间[32],JU等用杆状病毒表达的GPV VP2其分子量为75 kD[29],与笔者获得的72 kD大小相近。对VP2 氨基酸序列的糖基化、乙酰化、磷酸化位点进行预测,结果表明VP2序列上存在上述位点,说明VP2蛋白存在不同程度的翻译后修饰。对照野生型鹅细小病毒 VP2蛋白分子量与本试验结果的一致更验证了笔者的分析。VP2插入不影响病毒增殖特性以及VP2蛋白在DEV病毒载体中成功表达,为动物试验诱导抗体产生提供了保证。
体液免疫在抵抗小鹅瘟病毒感染过程中发挥着主要作用。经过GPV感染康复的雏鹅以及曾经隐性感染的成年鹅,均能产生高水平的抗体,并持续较长时间,而且能将抗体通过卵黄传给后代,使孵出的雏鹅获得抵抗GPV感染的能力。通过免疫母鹅或用高免血清注射雏鹅都对此病有预防作用[33]。JU等利用杆状病毒表达系统在昆虫细胞中分别表达了GPV 结构蛋白VP1、VP2和VP3,它们皆能自组装成病毒样颗粒(VLPs),且具有良好的免疫原性[29]。LEE等利用昆虫-杆状病毒表达系统表达GPV VP2,加以佐剂免疫9日龄鸭,首次免疫后14d进行二次免疫,能产生良好的抗体应答[34]。CHEN等同样利用昆虫-杆状病毒表达系统表达了密码子优化的GPV VP2基因,VP2能组装成VLPs,且VLPs保持了良好的免疫原性和免疫反应原性[35]。笔者的研究表明rDEV-VP2感染雏番鸭能诱导产生针对VP2蛋白的特异性抗体,推测其对小鹅瘟具有免疫保护能力。本研究是在鸭瘟病毒疫苗株BAC基础上开展,具有良好的安全性,且由该 BAC拯救出来的病毒rDEV-BAC和rDEV-Cre两病毒株皆对鸭瘟强毒株的攻击具有100%保护作用[26],推测本研究构建的重组病毒也具有保护鸭瘟强毒株攻击的能力,但攻毒保护试验有待进一步研究。
4 结论
成功构建了表达鹅细小病毒VP2蛋白的重组鸭瘟病毒。该重组病毒细胞生长特性与亲本毒株基本一致,且重组病毒在感染细胞中能表达外源蛋白VP2,接种雏番鸭能诱导VP2特异性抗体生成,具有研制小鹅瘟-鸭瘟二联活载体疫苗的潜力。
References
[1] 韦平. 重要动物病毒分子生物学. 北京: 科学出版社,2008: 501-516. WEI P. Molecular Biology of Important Animal Virus. Beijing: Science Press,2008: 501-516. (in Chinese)
[2] 刘光清. 动物病毒反向遗传学. 2版. 北京: 科学出版社,2014: 493-536. LIU G Q. Reverse Genetics of Animal Virus. 2nd edition. Beijing: Science Press,2014: 493-536. (in Chinese)
[3] DARTEIL R,BUBLOT M,LAPLACE E,BOUQUET J F,AUDONNET J C,RIVIÈRE M. Herpesvirus of turkey recombinant viruses expressing infectious bursal disease virus (IBDV) VP2 immunogen induce protection against an IBDV virulent challenge in chickens. Virology,1995,211(2): 481-490.
[4] LIU S,SUN W,CHU J,HUANG X,WU Z,YAN M,ZHANG Q,ZHAO P,IGIETSEME JU,BLACK CM,HE C,LI Y. Construction of recombinant HVT expressing PmpD,and immunological evaluation against Chlamydia psittaci and Marek's Disease Virus. PLoS One,2015,10(4): e0124992.
[5] KAPCZYNSKI D R,ESAKI M,DORSEY K M,JIANG H,JACKWOOD M,MORAES M,GARDIN Y. Vaccine protection of chickens against antigenically diverse H5 highly pathogenic avian influenza isolates with a live HVT vector vaccine expressing the influenza hemagglutinin gene derived from a clade 2. 2 avian influenza virus. Vaccine,2015,33(9): 1197-1205.
[6] KLINGBEIL K,LANGE E,TEIFKE J P,METTENLEITER T C,FUCHS W. Immunization of pigs with an attenuated pseudorabies virus recombinant expressing the haemagglutinin of pandemic swine origin H1N1 influenza A virus. Journal of General Virology,2014,95(Pt 4): 948-959.
[7] 邓晓辉,陈柳,叶伟成,李军星,崔尚金,余斌,云涛,崔言顺,孙元,仇华吉,张帅,张存. 共表达猪细小病毒VP2蛋白和猪圆环病毒2型Cap蛋白的重组伪狂犬病毒的构建及其鉴定. 中国预防兽医学报,2012,34(4): 251-256. DENG X H,CHEN L,YE W C,LI J X,CUI S J,YU B,YUN T,CUIY S,SUN Y,QIU H J,ZHANG S,ZHANG C. Construction and characterization of a recombinant pseudorabies virus expressing VP2 of porcine parvovirus and Cap of porcine circovirus type 2. Chinese Journal of Preventive Veterinary Medicine,2012,34(4): 251-256. (in Chinese)
[8] GIMENO I M,CORTES A L,FAIZ N M,HERNANDEZ-ORTIZ B A,GUY J S,HUNT H D,SILVA R F. Evaluation of the protection efficacy of a serotype 1 Marek's disease virus-vectored bivalent vaccine against infectious laryngotracheitis and Marek's disease. Avian Diseases,2015,59(2): 255-262.
[9] ZHANG Z,MA C,ZHAO P,DUAN L,CHEN W,ZHANG F,CUI Z. Construction of recombinant Marek's disease virus (rMDV) coexpressing AIV-H9N2-NA and NDV-F genes under control of MDV's own bi-directional promoter. PLoS One,2014,9(3): e9067.
[10] SAID A,LANGE E,BEER M,DAMIANI A,OSTERRIEDER N. Recombinant equine herpesvirus 1 (EHV-1) vaccine protects pigs against challenge with influenza A(H1N1)pmd09. Virus Research,173(2): 371-376.
[11] SAID A,DAMIANI A,MA G,KALTHOFF D,BEER M,OSTERRIEDER N. An equine herpesvirus 1 (EHV-1) vectored H1 vaccine protects against challenge with swine-origin influenza virus H1N1. Veterinary Microbiology,2011,154(1-2): 113-123.
[12] VAN DE WALLE G R,MAY M A,PETERS S T,METZGER S M,ROSAS C T,OSTERRIEDER N. A vectored equine herpesvirus type 1 (EHV-1) vaccine elicits protective immune responses against EHV-1 and H3N8 equine influenza virus. Vaccine,2010,28(4): 1048-1055.
[13] REN X G,XUE F,ZHU Y M,TONG G Z,WANG Y H,FENG J K,SHI H F,GAO Y R. Construction of a recombinant BHV-1 expressing the VP1 gene of foot and mouth disease virus and its immunogenicity in a rabbit model. Biotechnology Letters,2009,31(8): 1159-1165.
[14] SCHRIJVER R S,LANGEDIJK J P,KEIL G M,MIDDEL W G,MARIS-VELDHUIS M,VAN OIRSCHOT J T,RIJSEWIJK F A. Immunization of cattle with a BHV1 vector vaccine or a DNA vaccine both coding for the G protein of BRSV. Vaccine,1997,15(17-18): 1908-1916.
[15] DONOFRIO G,CAVIRANI S,SIMONE T,VAN SANTEN V L. Potential of bovine herpesvirus 4 as a gene delivery vector. Journal of Virological Methods,2002,101(1-2): 49-61.
[16] CAPOCEFALO A,MANGIA C,FRANCESCHI V,JACCA S,VAN SANTEN V L,DONOFRIO G. Efficient heterologous antigen gene delivery and expression by a replication-attenuated BoHV-4-based vaccine vector. Vaccine,2013,31(37): 3906-3914.
[17] DONOFRIO G,FRANCESCHI V,LOVERO A,CAPOCEFALO A,CAMERO M,LOSURDO M,CAVIRANI S,MARINARO M,GRANDOLFO E,BUONAVOGLIA C,TEMPESTA M. Clinical protection of goats against CpHV-1 induced genital disease with a BoHV-4-based vector expressing CpHV-1 gD. PLoS One,2013,8(1): e52758.
[18] LI Y,HUANG B,MA X,WU J,LI F,AI W,SONG M,YANG H. Molecularcharacterization of the genome of duck enteritis virus. Virology,2009,391: 151-161.
[19] WANG J,OSTERRIEDER N. Generation of an infectious clone of duck enteritis virus (DEV) and of a vectored DEV expressing hemagglutinin of H5N1 avian influenza virus. Virus Research,2011,159(1): 23-31.
[20] CHEN L,YU B,HUA J,YE W,NI Z,YUN T,DENG X,ZHANG C. Construction of a full-length infectious bacterial artificial chromosome clone of duck enteritis virus vaccine strain. Virology Journal,2013,10: 328.
[21] WANG J,GE A,XU M,WANG Z,QIAO Y,GU Y,LIU C,LIU Y,HOU J. Construction of a recombinant duck enteritis virus (DEV)expressing hemagglutinin of H5N1 avian influenza virus based on an infectious clone of DEV vaccine strain and evaluation of its efficacy in ducks and chickens. Virology Journal,2015,12: 126.
[22] ZOU Z,HU Y,LIU Z,ZHONG W,CAO H,CHEN H,JIN M. Efficient strategy for constructing duckenteritisvirus-based live attenuated vaccine against homologous and heterologous H5N1 avian influenza virus and duckenteritisvirus infection. Veterinary Microbiology,2015,46: 42.
[23] LIU J,CHEN P,JIANG Y,WU L,ZENG X,TIAN G,GE J,KAWAOKA Y,BU Z,CHEN H. A duck enteritis virus-vectored bivalent live vaccine provides fast and complete protection against H5N1 avian influenza virus infection in ducks. Journal of Virology,2011,85(21): 10989-10998.
[24] LIU J,CHEN P,JIANG Y,DENG G,SHI J,WU L,LIN Y,BU Z,CHEN H. Recombinant duck enteritis virus works as a single-dose vaccine in broilers providing rapid protection against H5N1 influenza infection. Antiviral Research,2013,97(3): 329-333.
[25] LIU X,WEI S,LIU Y,FU P,GAO M,MU X,LIU H,XING M,MA B,WANG J. Recombinant duck enteritis virus expressing the HA gene from goose H5 subtype avian influenza virus. Vaccine,2013,31(50): 5953-5959.
[26] CHEN P,LIU J,JIANG Y,ZHAO Y,LI Q,WU L,HE X,CHEN H. The vaccine efficacy of recombinant duck enteritis virus expressing secreted E with or without PrM proteins of duck tembusu virus. Vaccine,2014,32(41): 5271-5277.
[27] ZOU Z,LIU Z,JIN M. Efficient strategy to generate a vectored duck enteritis virus delivering envelope of duck Tembusu virus. Viruses,2014,6(6): 2428-2443.
[28] 陈柳,余斌,倪征,华炯钢,叶伟成,云涛,张存. 表达鸭坦布苏病毒 E蛋白的重组鸭瘟病毒的构建及其生物学特性研究. 浙江农业学报,2015,27(11): 1889-1895. CHEN L,YU B,NI Z,HUA J,YE W,YUN T,ZHANG C. Construction and characterization of a recombinant duck enteritis virus expressing E protein of duck Tembusu virus. Acta Agriculturae Zhejiangensis,2015,27(11): 1889-1895. (in Chinese)
[29] JU H,WEI N,WANG Q,WANG C,JING Z,GUO L,LIU D,GAO M,MA B,WANG J. Goose parvovirus structural proteins expressed by recombinant baculoviruses self-assemble into virus-like particles with strong immunogenicity in goose. Biochemicaland Biophysical Research Communications,2011,409(1): 131-136.
[30] TISCHER B K,VON EINEM J,KAUFER B,OSTERRIEDER N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques,2006,40(2): 191-197.
[31] TISCHER B K,SMITH G A,OSTERRIEDER N. En passant mutagenesis: a two step markerless red recombination system. Methods Molecular Biology,2010,634: 421-430.
[32] 李茂祥,李俊宝,郑玉美. 小鹅瘟病毒纯化及其理化特性的研究.病毒学报,1990,6(2): 155-159. LI M X,LI J B,ZHENG Y M. Purification and physico-chemical characteristics of a goslin plaque virus. Chinese Journal of Virology,1990,6(2): 155-159. (in Chinese)
[33] 殷震. 动物病毒学. 2版. 北京: 科学出版社,1997: 1165-1168. YIN Z. Aminal Virology. 2nd edition. Beijing: Science Press,1997: 1165-1168. (in Chinese)
[34] LEE J W,LIN Y M,YEN T Y,YANG W J,CHU C Y. CpG oligodeoxynucleotides containing GACGTT motifs enhance the immuneresponses elicited by a goose parvovirus vaccine inducks. Vaccine,2010,28: 7956-7962.
[35] CHEN Z,LI C,ZHU Y,WANG B,MENG C,LIU G. Immunogenicity of virus-like particles containing modified goose parvovirus VP2protein. Virus Research,2012,169: 306- 309.
(责任编辑 林鉴非)
Construction and Characterization of a Recombinant Duck Enteritis Virus Expressing VP2 Gene of Goose Parvovirus
CHEN Liu,YU Bin,NI Zheng,HUA Jiong-gang,YE Wei-cheng,YUN Tao,ZHANG Cun
(Institute of Animal Husbandry and Veterinary Medicine,Zhejiang Academy of Agricultural Sciences,Hangzhou 310021)
【Objective】Duck enteritis virus (DEV) and goose parvovirus (GPV) are considered to be two of the most important and widespread viruses infecting ducklings,Muscovy ducklings and goslings. According to the most recent virus taxonomy reported in 2012 by the International Committee on Taxonomy of Viruses (ICTV),DEV (also referred to Anatid herpesvirus 1) is classifiedinto the genus Mardivirus,the subfamily Alphaherpesvirinae of Herpesviridae. Many herpesviruses,such as Pseudorabies virus (PRV),Marek's disease virus (MDV),Herpesvirus of turkey(HVT)have been widely made as live viral vector for the expression of foreign antigens,and there were some reports about DEV as live viral vector in recent years. To control DEV and GPV infection,a recombinant vectored DEV expressing GPV VP2 was constructed in this study based on the bacterial artificial chromosome (BAC)clone pDEV-EF1 which carries DEV full-length genome (Chen L,et al. ,2015),and then the biological characteristics of the obtained recombinant virus rDEV-VP2 were analyzed to explore the possibility of rDEV-VP2 as duplex live carrier vaccine. 【Method】 The recombinant BAC clone pDEV-VP2 carrying GPV VP2 gene was generated by two-step Red/ET recombination in E. coli. pDEV-VP2 was constructed by inserting codon optimized-GPV VP2 expression cassette between DEV US7 and US8 genes on pDEV-EF1. The recombinant viruses rDEV-VP2 and rDEV-VP2-Cre without BAC sequence were rescued from chicken embryo fibroblasts (CEFs) by calcium phosphate precipitation. And the growth curve in vitro,plaque size and expression of GPV VP2 in CEFs were analyzed. The antibody level of GPV VP2 in sera of rDEV-VP2-incoculated ducklings was detected by an indirect-ELISA method based on the GPV VP2 protein. 【Result】 The recombinant viruses rDEV-VP2 and rDEV-VP2-Cre were rescued from chicken embryo fibroblasts (CEFs) by calcium phosphate precipitation. Growth curves show that the growth kinetics of rDEV-VP2 was basically consistent with those of parental virus in vitro. And the plaque size of rDEV-VP2 was slightly increased compared to the parental virus rDEV-BAC. Immunofluorescence assay and Western blot analysis showed that GPV VP2 protein is expressed in recombinant virus-infected CEFs. And the rDEV-VP2 infection could induce 7-day-old Muscovy ducklings to produce antibody specific for GPV VP2. 【Conclusion】 In this study,the antigen gene VP2 of GPV was inserted into the genome of DEV US7 and US8,and an recombinant infectious BAC clone of DEV was successfully constructed. Then the corresponding recombinant virus rDEV-VP2 was rescued,and its cellular growth characteristics were basically consistent with those of parental virus,and rDEV-VP2 could induce Muscovy ducklings to produce VP2-specific antibody. These studies have laid a foundation for developing bivalent vaccine controlling DEV and GPV infection.
duck enteritis virus; goose parvovirus; VP2; recombinant virus
2016-02-19;接受日期:2016-07-12
浙江省科技计划项目优先主题重点国际科技合作合作研究项目(2011C14011)、浙江省自然科学基金(LY15C180002)和浙江省公益技术应用研究项目(2016C32070)
联系方式:陈柳,Tel:0571-86404257;E-mail:haoliuzi@126.com。通信作者张存,Tel:0571-86404182;E-mail:zhangcun@aliyun.com
