CBF冷应答途径基因在热处理诱导的香蕉果实抗冷性中的作用
2016-09-19王海波苏新国张昭其庞学群
王海波,李 璐,苏新国,张昭其,庞学群
(1广东食品药品职业学院,广州 510520;2华南农业大学生命科学学院,广州 510642;3华南农业大学园艺学院,广州 510642)
CBF冷应答途径基因在热处理诱导的香蕉果实抗冷性中的作用
王海波1,李 璐2,苏新国1,张昭其3,庞学群2
(1广东食品药品职业学院,广州 510520;2华南农业大学生命科学学院,广州 510642;3华南农业大学园艺学院,广州 510642)
【目的】探讨CBF/DREB(C-repeat binding transcription factor/dehydrate responsive element binding factor)冷应答途径在热处理诱导香蕉抗冷性中的作用,为研究香蕉果实中CBF冷应答途径的信号转导过程提供参考依据。【方法】从香蕉基因组数据库(http∶//banana-hub.southgreen.fr/)中挑选 CBF冷应答途径的7个基因序列,分别设计特异引物,利用实时荧光定量 PCR分析这7个基因在热处理诱导的香蕉抗冷性中的表达模式。【结果】香蕉果实MaICE在7℃处理1 h时,表达迅速升高并达到最大值,DREB类基因(MaDREB1D、MaDREB1E、MaDREB1G和MaDREB3)在7℃处理1 h时,均存在一个表达高峰,MaCOR413在7℃下4 h时有一个表达高峰。这些结果表明,香蕉果实中在 7℃低温胁迫下中存在 CBF冷应答途径,即 ICE(inducer of CBF expression)-CBF-COR(Cold-regulated genes)通路。香蕉果实CBF冷应答途径的7个基因表达均在热处理(52℃,3 min)后0.5 h迅速达到最大值,进一步研究发现,香蕉果实经热处理后于7℃低温贮藏5 d时,MaDREB1D、MaDREB1E、MaDREB2C、MaDREB3和MaCOR413的表达均强于对照。【结论】7℃低温胁迫下MaICE、DREB类基因、MaCOR413表达依次增强,说明低温下香蕉果实中存在CBF冷应答途径。CBF冷应答途径相关基因表达增强可能参与了热处理诱导采后香蕉果实的抗冷性。
CBF/DREB;香蕉果实;热处理;抗冷性
0 引言
【研究意义】冷害是热带亚热带果蔬贮运保鲜中存在的主要问题之一[1]。热处理是提高果蔬抗冷性的最有效采后处理之一[2],但热处理诱导的抗冷机制特别是有关的信号传导途径目前还不太清楚。模式植物上的研究表明,低温转录因子CBF/DREB调控的信号传导途径可能是诱导植物抗冷性提高的最重要途径[3-5],但是有关 CBF冷应答途径在热处理诱导抗冷性中的作用目前很少报道。对CBF冷应答途径在热处理诱导香蕉抗冷性中的作用展开深入研究,将为了解香蕉果实中CBF冷应答途径的信号转导过程提供参考。【前人研究进展】模式植物上的研究表明,低温转录因子CBF调控的信号传导途径可能是诱导植物抗冷性提高的最重要途径,即ICE-CBF-COR通路。CBF是该途径中基因表达的“主开关”,受ICE调控的CBF转录因子能够特异结合启动子中含有CRT/DRE(C-repeat/ dehydration responsive element)的顺式元件,可以激活下游一系列冷调节基因 COR的表达,从而提高拟南芥等植物的抗寒性[6-8]。高温和低温均能诱导 CBF冷应答途径相关基因的表达。4℃低温诱导了蝴蝶兰PaCBF1在4 h开始表达,24 h达到最高峰。在拟南芥中过表达PaCBF1,诱导了AtCOR6.6和RD29a表达的增强[9]。桃果实经过0℃低温处理6 h时,PpCBF5表达水平增加约4倍,然后迅速增加至72 h的4 000多倍。PpCBF1和PpCBF6的表达,在0℃下72 h可分别被诱导575倍和6倍[10]。38℃热处理1 h诱导了黄瓜幼苗CBF1的表达,同时增加了其抗热性[11];高温胁迫诱导了拟南芥中DREB2A蛋白的产生,此蛋白作为一个调控因子参与拟南芥的热激胁迫反应[12]。高温和低温都能诱导新疆野苹果 MsDREB2C[13]、玉米ZmDREB2A[14]的表达增强。【本研究切入点】笔者前期研究结果表明,采用52℃热水处理香蕉果实3 min,能有效减轻香蕉果实的冷害症状[15-17]。然而热水处理是否通过CBF冷应答途径提高香蕉果实的抗冷性,目前尚不清楚。【拟解决的关键问题】研究香蕉果实在低温下是否存在CBF冷应答途径,以及CBF冷应答途径中的相关基因是否参与了热水处理诱导的香蕉抗冷性。
3.加强供应商关系管理,转变物资储备方式,改进资源占有方式。供应商关系管理是指运用供应链管理思想,通过搜寻、考核、评价等方式,对作为企业重要外部资源的供应商分层次建立合作关系,并对其关系进行管理和维护的活动。实行供应商资格预审制及供应商许可供应产品目录制,将其优势产品列入可供应产品目录。对每个有交易的供应商进行动态量化考核,建立供应商动态量化评价体系。考核得分由整体实力、供应量、合同执行情况、网上报价率、奖惩等5个部分组成。对考核优秀的供应商在招投标、付款、供应份额上给予倾斜,不断培养优秀的主力供应商群体,并逐步与之建立稳定的合作关系。
1 材料与方法
试验于2015年在华南农业大学园艺学院进行。
1.1材料及处理
香蕉品种为‘巴西’(Musa spp. AAA group cv. Brazil),采自广州市番禺香蕉园。挑选饱满度约 7成的绿熟香蕉,运回广东省果蔬保鲜重点实验室。挑选大小均匀、无病虫害及机械损伤的单个香蕉,先后用0.1%漂白粉和0.05%施保功各浸泡5 min,晾干备用。
热水及低温贮藏处理:将香蕉果实一次性浸入热水处理机(体积为400 L)中,温度为52℃,时间为3 min,热水处理前后温度差异在0.5℃以内。热水处理(hot water dipping,HWD)后将香蕉果实放入20℃恒温箱中贮藏3 h(前期研究表明热水处理后于20℃恢复3 h抗冷效果最好[16]),然后置于7℃下贮藏5 d,对照为未经过热水处理,其他贮藏条件一样。热处理香蕉100个,对照处理100个,每处理3个重复。取样时间点为:20℃下0、0.5、1.5、3 h;7℃下1、4、8、24、48、120 h。每个取样点10个香蕉,取香蕉中部果皮用液氮冷冻置于-80℃冰箱保存。
[16] WANG H B,ZHANG Z Q,XU L Y,HUANG X M,PANG X Q. The effect of delay between heat treatment and cold storage on alleviation of chilling injury in banana fruit. Journal of the Science of Food and Agriculture,2012,92: 2624-2629.
1.2.1实时定量引物设计 从香蕉基因组数据库(http://banana-genome.cirad. fr/)中搜索并挑选 CBF冷信号途径中的7个相关基因序列,设计实时定量引物,验证后备用。引物序列为:MaICE:5-TTGAGGTCA GGGTAAGAG和3-CTTCCTTGGATTGCTCAG;MaDREB1D:5-CATGGATTTG GGCTTCGGTTAC和3-GTCCACGTCGTCCCAGTTACA;MaDREB1E:5-CGATGAATGAGGAGGACGAGAT和3-CACC AAACTGATGCTGGAGGA;MaDREB1G:5-AACT TCGGAATGCAAGGGTA和3-CCGGTCTCAGATGG AATAGC;MaDREB2:5-GACGAGCACCAGCGA GTCAAG和3-AGAGCGGAATGTCACTGAAAGC;MaDREB3:5-CGAGTTCCTGTACGAAGACCCG和3-AGATGGTGGTGGGAACAACAGTG;MaCOR413:5-CTGACTGGTTGGAGATGC和3-TACCAAGCGCC AGATAGG。
1.2.2实时荧光定量 PCR(qRT-PCR) 实时荧光定量 PCR使用仪器为 BIO-RAD CFX9600,采用TOYOBO THUNDERBIRD SYBR®qPCR Mix 进行Real-time PCR扩增。qRT-PCR反应体系(20 μL)包括10 μL SYBR-Green PCR Master Mix,0.25 μL上游(10 μmol·L-1)和0.25 μL下游引物(10 μmol·L-1),2 μL稀释的cDNA以及7.5 μL ddH2O。反应条件为:95℃ 3 min,95℃ 10 s,59℃ 10 s,72℃ 20 s,循环数为40。为了确认产物的质量和引物的特异性,在溶解曲线中分析了产物的 Tm(分析溶解温度为 65—95℃)。利用Actin1[18]作为内参基因。所有的qRT-PCR反应根据内参基因Actin1进行Ct值校准,目的基因的相对表达水平利用公式2-ΔΔCt进行计算,重复3次。
1.2.3数据处理 用MEGA5.1软件对氨基酸序列构建系统进化树,采用 Sigmaplot12.5计算标准误并作图。
2 结果
2.1香蕉DREB/CBF基因的系统进化树分析
李镇西老师说:“让人们因我的存在而感到幸福。”学生及家长群里均是一致为此次自评与他评的活动点赞,有些学生还说,看了统计数据,后悔自己当初写得还不够用心,字太丑。因为是寒假,不少学生都表示这个是比红包还让人开心的新年礼物,而且很有纪念意义,特别是某些同学还很用心地画了素描或漫画肖像,温暖而贴心。不少家长说,原来自己的儿女在同学、朋友中为人处世还蛮多优点,自己平时真的难有机会发现,感谢老师、同学们的用心。让学生及其家长看了那些用心的自评与他评而感动快乐,我也感受到了班级的幸福指数直线上升,新年里班级Q群中氛围轻松愉悦。
从香蕉基因组数据库搜索到5个DREB类基因:MaDREB1D、MaDREB1E、MaDREB1G、MaDREB2C 和MaDREB3。参考SAKUMA[19]、MIZOI[20]等的研究结果,从拟南芥、水稻等物种选取已克隆的DREB/CBF转录因子的氨基酸序列,用MEGA5.1软件构建系统进化树。从图1可知,MaDREB1E和MaDREB1G属于 DREB/CBF 转录因子 A-1亚类,MaDREB1D属于A-6亚类、MaDREB2C属于A-3亚类、MaDREB3属于A-4亚类。
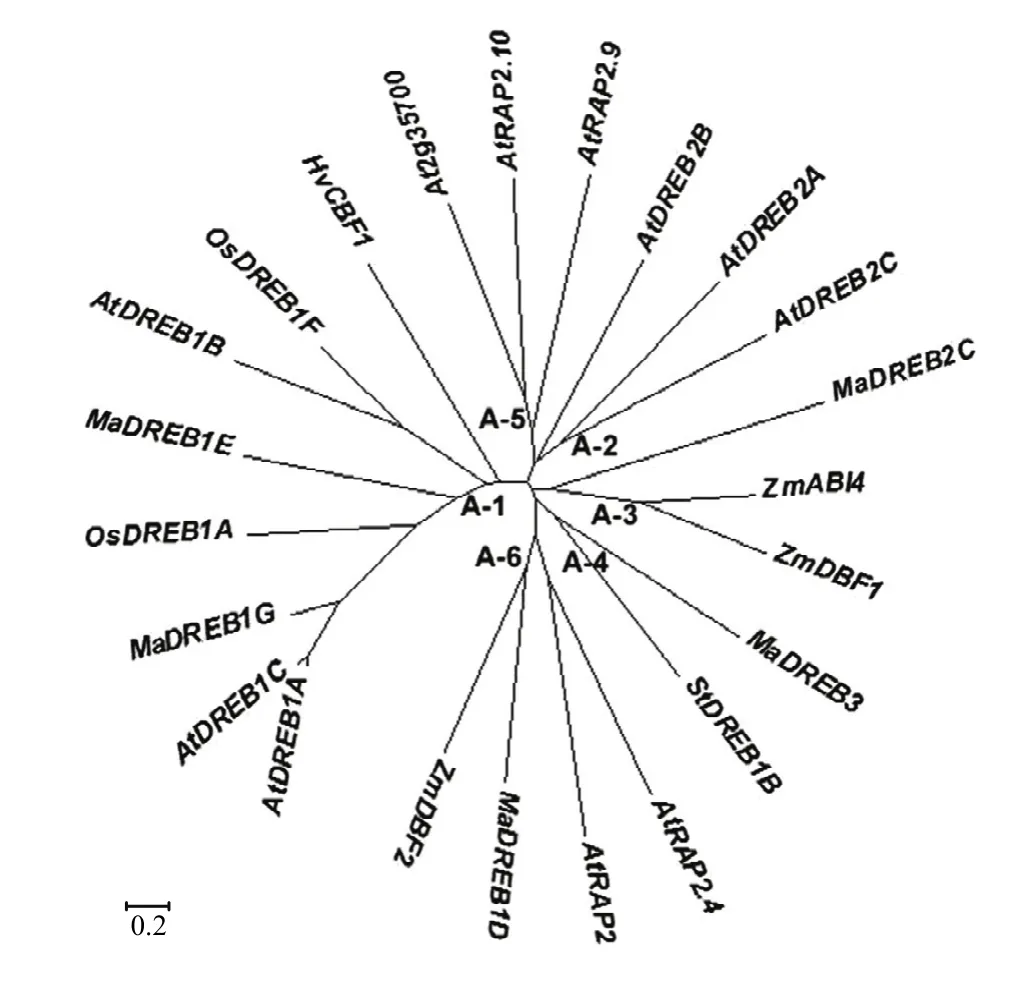
图1 植物中DREB类基因的系统进化树Fig.1 Phylogenetic tree of the DREB subfamily transcription factors in plants
2.2热激处理及热激后低温贮藏对香蕉 MaICE表达的影响
[7] WU L H,ZHOU M Q,SHEN C,LIANG J,LIN J. Transgenic tobacco plants over expressing cold regulated protein CbCOR15b from Capsella bursa-pastoris exhibit enhanced cold tolerance. Journal of Plant Physiology,2012,169: 1408-1416.

图2 热处理和低温贮藏对香蕉果实MaICE表达的影响Fig.2 Effect of hot water dipping (HWD) and cold storage on the expression of MaICE in banana fruits
2.3热激处理及热激后低温贮藏对香蕉DREB类基因表达的影响
从图3—图5可知,未经热处理的香蕉MaDREB1D、MaDREB1E和MaDREB1G这 3个基因表达均在7℃1 h有一个高峰。说明7℃低温能在1 h内快速诱导这3个基因表达的升高。MaDREB1D、MaDREB1E 和MaDREB1G这3个基因表达均在热处理后0.5 h有一个高峰。在 7℃低温贮藏过程中,HWD处理的MaDREB1D表达一直高于对照,MaDREB1E表达在4 —24 h高于对照,而MaDREB1G表达与对照差异不大。
《北爱》之后我开始认真学习,唯一接的工作是《快乐大本营》。现在回头看看,那时候整个人都土炸了,穿得很随便,人又胖,还脑子抽筋地表演了一段东北扭秧歌,用现在的话说就是黑历史。

图3 热处理和低温贮藏对香蕉果实MaDREB1D表达的影响Fig.3 Effect of HWD and cold storage on the expression of MaDREB1D in banana fruits
从图6可知,对照的MaDREB2C表达在7℃下4 h后逐渐增强,直至24 h有一个高峰,之后逐渐下降至较低的水平。而HWD处理后的0.5 h,其MaDREB2C表达有一个短暂而迅速的升高,低温下其MaDREB2C表达变化趋势与对照一致,并且均高于对照。
从图7可知,对照的MaDREB3表达在7℃下1 h有一个高峰,而HWD处理后MaDREB3表达在HWD 后0.5 h有一个小高峰,表达比对照稍高。在7℃低温贮藏过程中,MaDREB3表达在前期低于对照,后期则高于对照。
对于有关企业人力资源管理工作的高素质人才,要适当进行鼓励。比如说,对于高效率完成相关工作的工作人员,要适当给予一定程度上的福利待遇,激励员工更加努力有效地工作。另外,对于长期处于压力状态下的企业部门,企业的人力资源管理部门应组织相关人员进行适当的放松。例如,可以组织相关人员进行内部聚餐,或者在工作告一段落之后,组织相关工作人员出去游玩,放松心情等等,都是企业人力资源部门应该组织开展的有效策略。

图4 热处理和低温贮藏对香蕉果实MaDREB1E表达的影响Fig.4 Effect of HWD and cold storage on the expression of MaDREB1E in banana fruits

图5 热处理和低温贮藏对香蕉果实MaDREB1G表达的影响Fig.5 Effect of HWD and cold storage on the expression of MaDREB1G in banana fruits
2.4热激处理及热激处理后低温贮藏对香蕉MaCOR413表达的影响
[2] 陆旺金,张昭其,季作梁. 热带亚热带果蔬低温贮藏冷害及御冷技术. 植物生理学通讯,1999,35(2): 158-163. LU W J,ZHANG Z Q,JI Z L. Chilling injury and approaches to reduce chilling injury of tropical and subtropical fruits and vegetables during low temperature storage. Plant Physiology Communications,1999,35(2): 158-163. (in Chinese)

图6 热处理和低温贮藏对香蕉果实MaDREB2C表达的影响Fig.6 Effect of HWD and cold storage on the expression of MaDREB2C in banana fruit

图7 热处理和低温贮藏对香蕉果实MaDREB3表达的影响Fig.7 Effect of HWD and cold storage on the expression of MaDREB3 in banana fruit
3 讨论
[9] PENG P H ,LIN C H ,TSAI H W ,LIN T Y. Cold response in Phalaenopsis aphrodite and characterization of PaCBF1 and PaICE1. Plant and Cell Physiology,2014,55(9): 1623-1635.

图8 热处理和低温贮藏对香蕉果实MaCOR413表达的影响Fig.8 Effect of HWD and cold storage on the expression of MaCOR413 in banana fruit
香蕉果实 MaDREB2C在低温下的表达模式与其他DREB类基因有所不同。香蕉果实(对照)在7℃低温胁迫下24 h才出现表达高峰,这与其他DREB类基因均在 7℃低温 1 h出现表达高峰不同。说明MaDREB2C在香蕉果实低温贮藏后期产生响应。以上结果表明,7℃低温胁迫下,MaICE、DREB类基因、MaCOR413表达依次增强,说明低温下香蕉果实中存在CBF冷应答途径,即ICE-CBF-COR通路。
热处理是提高采后果实抗冷性的有效手段,笔者前期研究结果表明,采用 52℃热水处理香蕉果实 3 min,能有效减轻香蕉果实的冷害症状[15-17]。同时,热处理也能够诱导CBF冷应答途径相关基因的表达。例如,40℃高温处理能诱导马铃薯植株中的AtCBF3表达增强,并且发现在40℃胁迫l h时,叶中AtCBF3表达量最大[27]。将军菊苣幼苗高温37℃处理1 h后,叶片和根中CiDREB2A表达量即上升,并一直维持高表达,24 h表达量达到峰值[28]。CAO等[29]利用低温、高温和干旱对巨桉处理,对17个EgrDREB1/CBF和6个EgrDREB2研究,发现高温能诱导所有基因的表达增强,低温能诱导16个EgrDREB1/CBF的表达,认为 EgrDREB1/CBF亚家族主要是受低温诱导,而EgrDREB2主要是对干旱敏感。从本研究中的图2至图8可知,香蕉果实经过热处理后,CBF冷应答途径相关的7个基因表达均在热处理后的0.5 h有一个短暂而迅速的升高,在3 h恢复到处理前较低的水平。香蕉果实CBF冷应答途径的7个基因均在热处理后0.5 h的迅速表达增强,说明香蕉果实CBF冷应答途径的相关基因能受到热处理诱导快速表达增强。
进一步研究发现,热激处理后在低温贮藏时,CBF冷应答途径相关基因也会受到诱导增强。猕猴桃果实AcCBF在冷藏期间的相对表达量呈现先升高后下降的趋势,表达高峰在贮藏20 d时出现。而利用35℃和45℃热处理与对照相比显著提高了 AcCBF的表达峰值[30]。香蕉果实经热处理后于7℃低温贮藏5 d,发现MaDREB1D、MaDREB1E、MaDREB2C、MaDREB3 和MaCOR413等基因的表达均强于对照,MaICE表达出现峰值的时间从 1 h(对照)推迟到 8 h,而MaDREB1G表达与对照差异不大。以上研究表明,香蕉果实经过热处理后,7个CBF冷应答途径基因在热处理后0.5 h的迅速表达增强,可能是作为一个早期信号,诱导了后期CBF冷应答途径相关基因的进一步表达增强,导致香蕉果实抗冷性增强。这说明热处理诱导采后香蕉果实的耐冷性可能与 CBF冷应答途径密切相关。目前有关热处理诱导CBF冷应答途径相关基因表达方面的研究报道很少,热处理诱导的CBF冷应答途径相关基因表达提高与植物抗冷性之间的关系需要进一步研究。
4 结论
7℃低温胁迫下,MaICE、DREB类基因、MaCOR413表达依次增强,说明低温下香蕉果实中可能存在CBF冷应答途径,即ICE-CBF-COR通路。热激处理能快速诱导CBF冷应答途径相关基因的表达,并且DREB类基因和MaCOR413在7℃低温胁迫下表达强于对照。推测CBF冷应答途径相关基因表达增强可能参与了热处理诱导采后香蕉果实的抗冷性。
References
[1] 张昭其,庞学群. 南方水果贮藏保鲜技术. 南宁: 广西科学技术出版社,2008. ZHANG Z Q,PANG X Q. Technique of Postharvest Handling and Storage of South China Fruits. Nanning: Guangxi Technology Press,2008. (in Chinese)
从图8可知,对照的MaCOR413表达在7℃下4 h有一个高峰,其他时间的表达量均较低。而HWD的MaCOR413表达在热处理后0.5 h有一个高峰,在7℃下4 h有一个高峰,整个过程,HWD的MaCOR413表达量均高于对照。
在用Photoshop修图时,先复制背景图层,然后点击“滤镜—模糊—动感模糊”,用滑块调整模糊程度。我们最高调到了500像素。接下来添加蒙版,用不透明度15%的黑色笔刷涂抹你想保留的部分,比如这张照片里的码头。
[3] GILMOUR S J,ZARKA D G,STOCKINGER E J,SALAZAR M P,HOUGHTON J M,THOMASHOW M F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression,Plant Journal,1998,16: 433-442.
我国天然气生产和供给能力也持续增强。2017年,我国生产天然气1480.3亿立方米,同比增长8.2%,在全世界天然气生产国中增速最快。不过,天然气需求增长更快,爆发式的增长造成了短期的天然气供应出现了问题。从我国天然气产量可以看出,国内天然气正处于很辉煌的发展时期,但现阶段供应主要体现在进口方面。2017年,我国天然气进口量快速增长至946亿立方米,占天然气消费的40%。
[4] NAKAMURA J,YUASA T,HUONG T T,HARANO K,TANAKA S,IWATA T,PHAN T,IWAYA-INOUE M. Rice homologs of inducer of CBF expression (OsICE) are involved in cold acclimation. Plant Biotechnology,2011,28(3): 303-309.
[11] TALANOVA V V,TITOV A F,TOPCHIEVA L V,MALYSHEVA I E. Effect of stress factors on expression of the gene encoding a CBF transcription factor in cucumber plants. Doklady Biological Sciences,2008,423: 419-421.
[6] CHEN Y,CHEN Z L,KANG J Q,KANG D M,GU H Y,QIN G J. AtMYB14 regulates cold tolerance in Arabidopsis. Plant Molecular Bbiology Reporter,2013,31: 87-97.
前期研究表明,7℃(冷害温度)下,香蕉果实在第5天已经出现明显的冷害症状,利用52℃热水处理(HWD)3 min能有效提高香蕉果实在7℃低温贮藏中的抗冷性[16-17]。香蕉用 52℃热水处理(HWD)3 min ,然后在20℃下放置3 h后置于7℃下贮藏5 d。从图2可知,未经热处理的香蕉(对照)MaICE表达在7℃处理1 h时迅速升高达到最大值,之后又迅速降低为低温前的水平。香蕉经热处理后的0.5 h,其MaICE表达迅速升高,之后又下降到对照水平,7℃低温贮藏下8 h有一个高峰。说明香蕉MaICE在20℃下表达量很低,7℃低温能在1 h内快速诱导MaICE表达的升高。热处理能在0.5 h内快速诱导MaICE表达的升高,而7℃低温下出现高峰的时间从1 h推迟到8 h。
[8] HADI F,GILPIN M,FULLER M P. Identification and expression analysis of CBF/DREB1 and COR15 genes in mutants of Brassica oleracea var. botrytis with enhanced praline production and frost resistance. Plant Physiology and Biochemistry,2011,49: 1323-1332.
多个植物物种中的研究表明,低温转录因子CBF调控的信号传导途径可能是诱导植物抗冷性提高的最重要途径。DREB/CBF转录因子是AP2/EREBP转录因子家族中的1个亚家族,含有1个保守的AP2/EREBP结构域。拟南芥基因组DREB/CBF亚族又分为6个亚类(A1—A6)[19]。DREB1/CBF类主要受到低温诱导,DREB2类主要受到干旱和高盐诱导,而对于A3—A6目前研究较少。从图1的系统进化树发现香蕉的5个DREB基因属于不同的亚类。众多研究证实,CBF/DREB类基因在多种高等植物低温下的表达模式均是在低温下短时间内快速诱导增强,最终通过诱导COR表达增强从而提高植物的抗冷性。例如,拟南芥2.5℃低温15 min即可诱导AtCBF1、AtCBF2和AtCBF3的表达,2 h后达到表达高峰,随后下降,同时在低温2 h后COR开始被诱导表达[21-22]。过量表达CBF1或CBF3可促进 COR高水平表达,使转基因拟南芥植株的抗冷性提高[23]。宽叶独行菜LICBF在4℃低温胁迫下1 h的表达迅速增强,之后4—48 h逐渐降低至低温处理前的水平[24]。本研究结果表明,香蕉果实(对照)的MaICE表达在7℃处理1 h时迅速升高达到最大值,DREB类基因(MaDREB1D、MaDREB1E、MaDREB1G、和MaDREB3)表达在7℃处理1 h均存在一个高峰,随后MaCOR413表达在7℃下4 h出现一个高峰。其中MaDREB1D的表达模式与其他DREB有所不同,除了在1 h有一个高峰,MaDREB1D在120 h(冷害症状明显)时还出现一个表达最高峰。采后香蕉果实在低温胁迫下的 CBF冷应答基因表达模式与拟南芥类等植物类似,进一步验证了CBF冷应答途径在高等植物中保守性很高。将CBF冷应答途径基因转入拟南芥中,均提高了拟南芥的抗冷性。例如,ZHUANG等[25]将具有抗冷性草地早熟禾中的PpCBF3转化至拟南芥幼苗中,通过瞬时表达,发现4℃下0.5 h,PpCBF3表达开始增强,在1 h迅速最高峰,之后3—24 h迅速下降至较低的水平。进一步研究发现,4℃低温能诱导野生型和转基因拟南芥COR15a、COR6.6、COR47和COR78的表达,且转基因拟南芥中这些COR的表达高于野生型拟南芥。类似地,将中国野生葡萄中的VaICE1/2在拟南芥中过表达,增强了下游基因CBF1、COR15A和COR47的表达,提高了拟南芥的抗冷性[26]。
[10] LIANG L,ZHANG B,YIN X R,XU C J,SUN C D,CHEN K S. Differential expression of the CBF gene family during postharvest cold storage and subsequent shelf-life of peach fruit. Plant Molecular Biology Reporter,2013,31(6): 1358-1367.
月季(Rosa chinensis Jacq.)是蔷薇科蔷薇属植物,原产于我国,在我国有 2000 多年的栽培历史[1]。由于其极强的观赏性和适应性,广泛应用于园林绿化中,成为我国北方城市北京、天津等多个城市市花,建立了多处月季品种齐全、集中的专类公园。
不同施工方法沿地铁区间隧道方向地表沉降曲线见图8,台阶法施工引起地表变形量为最大,达到18.5 mm,施工引起的地表沉降越靠近高铁盾构隧道会越小,超过高铁盾构隧道中心截面距离30 m以外时变形基本趋于一致。
[5] ZHANG X,FOWLER S G,CHENG H,LOU Y,RHEE S Y,STOCKINGER E J,THOMASHOW M F. Freezing-sensitive tomato has a functional CBF cold response pathway,but a CBF regulon that differs from that of freezing-tolerant Arabidopsis. Plant Journal,2004,39(6): 905-919.
[12] SCHRAMM F,LARKINDALE J, KIEHLMANN E,GANGULI A,ENGLICH G,VIERLING E,KOSKULL-DÖRING P V. A cascade of transcription factor DREB2A and heat stress transcription factor HsfA3 regulates the heat stress response of Arabidopsis. Plant Journal,2008,53(2): 264-274.
中国传统文化与医学生的思想政治教育相融合,能有效提高医学生的意识自主性,能让医学生在接收医学知识的基础上,加深对生命意义的理解,从而使其不断提高自我品行修养。
[13] ZHAO K,SHEN X J,YUAN H Z,LIU Y,LIAO X,WANG Q,LIU L L,LI F,LI T H. Isolation and characterization of dehydrationresponsive element-binding factor 2C (MsDREB2C) from Malus sieversii Rome. Plant and Cell Physiology,2013,54(9): 1415-1430.
[14] QIN F,KAKIMOTO M,SAKUMA Y,MARUYAMA K,OSAKABE Y,TRAN L S,SHINOZAKI K,YAMAGUCHISHINOZAKI K. Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant Journal,2007,50( 1): 54-69.
[15] WANG H B,ZHANG Z Q,HUANG X M,JIANG Y M,PANG X Q. Hot water dipping induced chilling resistance of harvested banana fruit. Acta Horticulturae,2008,804: 513-522.
1.2测定方法
至臻天文台认证由瑞士联邦计量研究院 (METAS)核准,其精准程度毋庸置疑。只有具备卓越精准度和防磁性能的腕表,才能获得至臻天文台认证。
[17] 王海波,庞学群,黄雪梅,张昭其. 活性氧在热处理诱导香蕉耐冷性中的作用. 中国农业科学,2012,45(5): 936-942. WANG H B,PANG X Q,HUANG X M,ZHANG Z Q. The role of reactive oxygen species in heat treatment-induced chilling tolerance in banana fruit. Scientia Agricultura Sinica,2012,45(5): 936-942. (in Chinese)
[18] CHEN L,ZHONG H Y,KUANG J F,LI J G,LU W J,CHEN J Y. Validation of reference genes for RT-qPCR studies of gene expression in banana fruit under different experimental conditions. Planta,2011,234(2): 377-390.
[19] SAKUMA Y,LIU Q,DUBOUZET J G,ABE H,SHINOZAKI K,YAMAGUCHI-SHINOZAKI K. DNA-binding specificity of the ERF /AP2 domain of Arabidopsis DREBs transcription factors involved in dehydration and co1d-induciblegene expression. Biochemical and Biophysical Research Communications,2002,290(3): 998-1009.
[20] MIZOI J,SHINOZAKI K,YAMAGUCHI-SHINOZAKI K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochimica et Biophysica Acta,2012,1819(2): 86-96.
[21] STOCKINGER E J,GILMOUR S J,THOMASHOW M F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE,a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proceedings of the National Academy of Sciences of the United States of America,1997,94: 1035-1040.
[22] ZARKA D G,VOGET J T,COOK D AND THOMASHOW,M F. Cold induction of Arabidopsis CBF genes involves multiple ICE (inducer of CBF expression) promoter elements and a cold-regulatory circuit that is desensitized by low temperature. Plant Physiology,2003,133: 910-918.
[23] LIU Q,KASUGA M,SAKUMA Y,ABE H,MIURA S,YAMAGUCHI-SHINOZAKI K,SHINOZAKI K. Two transcription factors,DREB1 and DREB2,with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought and low temperature responsive gene expression,respectively,in Arabidopsis. Plant Cell,1998,10: 1391-1406.
[24] AKHTAR M,JAISWAL A,JAISWAL J P,QURESHI M I,TUFCHIM,SINGH N K. Cloning and characterization of cold,salt and drought inducible C-repeat binding factor gene from a highly cold adapted ecotype of Lepidium latifolium L. Physiology and Molecular Biology of Plants,2013,19(2): 221-230.
[25] ZHUANG L L, YUAN X Y,CHEN Y,XU B,YANG Z M,HUANG B R. PpCBF3 from Cold-Tolerant kentucky bluegrass involved in freezing tolerance associated with up-regulation of cold-related genes in transgenic Arabidopsis thaliana. PLoS One,2015,10(7): e0132928.
[26] XU W R,JIAO Y T,LI R M,ZHANG N B,XIAO D M,DING X L,WANG Z P. Chinese wild-growing Vitis amurensis ICE1 and ICE2 encode MYC-type bHLH transcription activators that regulate cold tolerance in Arabidopsis. PLoS One,2014,9(7): e102303.
[27] DOU H O,XV K P,MENG Q W,LI G,YANG X H. Potato plants ectopically expressing Arabidopsis thaliana CBF3 exhibit enhanced tolerance to high-temperature stress. Plant Cell Environ,2015,38(1): 61-72.
[28] 赵龙. 将军菊苣 DREB家族基因的克隆、功能研究及其遗传转化体系的建立[D]. 南京: 南京农业大学,2013. ZHAO L. Isolation and characterization of the dreb transcription factor from commander chicory and establishment of genetic transformation system of commander chicory [D]. Nanjing: Nanjing Agricultural University,2013. (in Chinese)
巷道周边破裂围岩在支护过程中,锚杆、锚索预紧力或支架提供的初撑力提高了围压。为了研究围压增量对岩石峰后蠕变的影响,对HS7-1与HS7-2进行了施加1MPa围压增量后的峰后蠕变试验,并与施加围压增量前的HS6-1与HS6-2作对比分析。HS7-1与HS7-2试验特征参数见表2,施加围压增量前后分别加载峰后蠕变曲线如图7所示。从图7可以看出,施加围压增量后,各级应力水平下蠕变变形速率明显减小,等速蠕变阶段延长,最后一级应力水平下进入加速蠕变阶段时间延后。在各级应力水平下的蠕变量上,依据试验数据拟合了应力水平-蠕变量的关系曲线见图8。
医养结合下的社区养老设施体系涵盖了社区居家养老设施与机构养老设施所包含的各类设施.本文研究社区层面的养老设施体系构建,结合目前居住区分级、老年人自身的特点以及需要、养老设施建设利用的效率综合考虑层级划分.
[29] CAO P B,AZAR S ,SANCLEMENTE H,MOUNET F,DUNAND C,MARQUE G ,MARQUE C,TEULIÈRES C. Genome-wide analysis of the AP2/ERF family in Eucalyptus grandis: an intriguing overrepresentation of stress-responsive DREB1/CBF genes. PLoS One,2015,10(4): e0121041.
完善统一的法律法规体系,既是公共部门的行为“标尺”,也是私人部门的利益“保护网”。PPP模式下的城市社区居家养老服务项目是公共部门和私人部门的共有产权,两者利益共享、风险共担,任何一方的失信或失责都会给整个项目的运营造成极大的损害。公私部门双方应明确各自的职责与定位,公共部门做好土地、政策、补贴等工作,私人部门投入资金、技术和管理等。因此,需要一整套统一完善的法律法规体系,以法律来明确双方的权责关系,保证项目运营的协调性,以法规规章来统一标准、规范程序,保证项目运营的规范性。
[30] MA Q S,SUO J T,HUBER D J,DONG X Q,HAN Y,ZHANG Z K,RAO J P. Effect of hot water treatments on chilling injury and expression of a new C-repeat binding factor (CBF) in ‘Hongyang’kiwifruit during low temperature storage. Postharvest Biology and Technology,2014,97: 102-110.
(责任编辑 赵伶俐)
The Role of CBF Cold Response Pathway Gene in Heat Treatment-Induced Chilling Tolerance in Banana Fruits
WANG Hai-bo1,LI Lu2,SU Xin-guo1,ZHANG Zhao-qi3,PANG Xue-qun2
(1Guangdong Food and Drug Vocational College,Guangzhou 510520;2College of Biotechnology,South China Agricultural University,Guangzhou 510642;3College of Horticulture,South China Agricultural University,Guangzhou 510642)
【Objective】The objective of this study is to investigate the role of C-repeat binding transcription factor (CBF)cold-resistance pathway in heat treatment -induced chilling tolerance in banana fruits and to provide reference for the study of the signal transduction of CBF cold response pathway in banana fruits.【Method】The sequences of 7 genes related to CBF cold response pathway were selected from the banana genome database (http://banana-genome-hub.southgreen.fr/),and then specific primers were designed respectively. Analysis of these 7 genes’ expression patterns of the heat-induced chilling tolerance in banana fruits was conducted using quantitative real-time PCR.【Result】The expression of MaICE gene of banana fruit increased rapidly and reached the maximum level at 7℃ for 1 hour. The expression of DREB (MaDREB1D,MaDREB1E,MaDREB1G and MaDREB3) genes displayed a peak at 7℃ for 1 hour. The expression of MaCOR413 gene had a peak at 7℃ for 4 hours. These results indicated that the CBF cold response pathway (ICE-CBF-COR pathway) exist in banana fruit during cold storage at 7℃. The expression of 7 gene related to CBF cold response pathway reached the maximum levels at 0.5 hour after heat treatment(52℃ 3 min). Furthermore,when the heat treated banana fruits storage at 7℃ for 5 days,the expressions of MaDREB1D,MaDREB1E,MaDREB2C,MaDREB3 and MaCOR413 genes were higher than the non-heated control.【Conclusion】 MaICE,DREB,and MaCOR413 increased in turn when banana fruits stored at 7℃. The CBF cold response pathway exist in banana fruits during cold storage at 7℃. Enhancement of gene expression related to CBF cold response pathway may be involved in chilling tolerance induced by heat treatment in banana fruits.
CBF/DREB; banana fruit; heat treatment; chilling tolerance
2016-03-14;接受日期:2016-05-25
国家自然科学基金(31301584)、广东省自然科学基金(S2013010013987)、广东省高校优秀青年教师项目(2014)、广东省科技计划项目(2012A020100007)、国家科技支撑计划(2015BAD16B07)
联系方式:王海波,Tel:020-29164658;E-mail:haibovip@126.com。通信作者庞学群,Tel:020-85283413;E-mail:xqpang@scau.edu.cn
