无患子苗期生物量积累和分配规律研究
2016-09-13周世勇
周世勇
(建阳市林业局,福建 建阳 354200)
无患子苗期生物量积累和分配规律研究
周世勇
(建阳市林业局,福建 建阳 354200)
通过定量分析无患子苗期生物量的积累和分配规律,并通过SPSS进行回归分析,拟合苗木根与茎生物量的异速生长方程、各器官及全株生物量与苗高、地径的回归关系。结果表明:无患子1年生播种苗生物量积累可划分为幼苗期、速生期和生长后期3个阶段。幼苗期根生物量分配比例最少,叶生物量分配比例最大,分别占全株生物量的16.85%和58.80 %;进入速生期后,生物量的积累中心逐渐转移到根、茎,而茎是生物量积累的最主要器官,占全株生物量的61.55 %。进入生长后期,根、茎进一步木质化,而叶片由于脱落生物量分配比例骤减,仅占全株生物量的11.44 %。根与茎生物量的异速积累、苗木各器官及全株生物量与苗木地径、苗高生长量均存在着极显著的相关关系,拟合的数学模型W=0.05(D2H)0849可靠性较高,可用于指导生产。
无患子;生物量;积累;分配规律;数学模型
苗木生物量的积累与分配是苗木季节性生长的结果。作为影响苗木质量的主要因素之一,生物量的大小能够较好地反映植物的生存能力,初期生长快、生物量大的苗木,具有较强的环境适应能力[1-2]。因此,研究苗木生物量积累和各器官分配规律,分析苗木的生产力和适应能力,培育出优质的苗木,从而提高苗木造林效果。
无患子(Sapindus mukorossi)为无患子科无患子属落叶大乔木,树高可达25m,原产于我国长江流域以南及中南半岛各地,印度和日本,对立地条件要求不高,在低山丘陵、石灰岩山地等酸性、碱性钙质土壤上均能较好地生长。因其生长迅速、树冠庞大、外形美观、秋叶金黄,且对汽车尾气、工业废气具有较强的吸收能力,故常作为行道树、庭荫树,是我国南方常见的园林绿化树种之一[3-5]。当前,对无患子的研究多集中于化学成分及分离技术[6-8]、遗传变异[9-10]、分子生物标记[11-12]、生理生化活动[13-15]和优树选择[16]等方面,而对无患子苗木生长规律的研究仅只是报道了地径、苗高的生长节律[17],未见苗木生物量积累分配规律的研究。本研究探讨1年生无患子播种苗生物量积累和分配规律,为开发利用这一生物质能源树种,培育优质壮苗提供参考依据。
1 试验地概况
试验地设在建阳市闽北武夷山南面,地处北纬27°06′~27°43′,东经117°31′~118°38′,属中亚热带季风气候,温和湿润,四季分明,日照充足,雨季集中。年平均降雨量1 907mm;年平均气温18.1 ℃,极端最高气温41.3 ℃,极端最低气温-8.7 ℃,无霜期280d。年平均日照总时数1 802.7h,7月最多,达250h;2月最少,为87.7h。土壤类型主要为红壤,占所有土壤类型面积的80%。
2 材料与方法
2.1试验材料及设计
试验种子由建阳市林业局种苗站提供。播种前将无患子种子放入缸中用自然水温浸泡,用水选法筛选出发育良好的种子,2~3d后捞起种子冲净待播。3月25日播种,在整好的苗床上,将处理好的种子按株行距10cm×15cm(密度控制在53 株/m2)播种,3.0cm深穴,浇水后点播,播种时将种胚朝下。播种后2周左右幼芽陆续出土,至4月下旬出苗基本整齐,随机设置3个观测小样方,样方面积1m×1m,每个样方内随机选取15株长势良好且基本一致的幼苗编号,定时(间隔30d左右)定株测量苗高和地径,得出该样方内平均苗高、地径,然后再在样方内不同的3个方向选择有代表性的平均样苗3株,全株挖出洗净,逐株测定其苗高、地径、主根长、侧根数,然后在105 ℃的烘箱烘干,测其干质量,求其平均值。试验均重复3次,苗木生长期间按常规育苗技术管理。
2.2数据分析方法
数据的分析作图采用Excel2003,回归方程的拟合采用SPSS18.0。
苗木不同器官间的生物量关系可用幂函数的形式来表示[18],其数学表达式为:Y=AXB
式中:A为待定常数;B为异速生长常数。
前人的研究成果表明[18-20],苗木生物量随地径、苗高的生长呈线性或非线性幂函数型增长,其数学模型如下:
W=a+bD+cC
(1)
W=a(D2H)b
(2)
W= aDbHc
(3)
式中:W为某一时期苗木器官或全株生物量;a、b、c均为待定系数。
3 结果与分析
3.1苗木生物量积累规律
将无患子全株生物量积累划分为幼苗期、速生期和生长后期3个阶段。其中,从4月28日(34d)—6月25日(92d)为幼苗期,这一时期苗木主要是对周围环境的适应生长,此时苗木各器官生长开始分化,并不能对所生存环境的光、温、水、养分等资源充分吸收利用,个体竞争力较弱,只能满足正常的生长需求,苗木生物量积累缓慢;从6月25日(92d)—10月24日(213d)苗木进入生物量快速积累期,苗木个体逐渐适应周边生存环境,充分利用外界资源迅速生长,根、茎的木质化程度增大,尤其是茎生物量积累远超过根和叶;之后从10月24日(213d)—11月25日(245d) 苗木进入生长后期,由于光照强度减弱、光合作用时间缩短、降水减少,苗木生物量积累变缓。无患子苗木生长不同时间根、茎、叶及全株生物量干物质积累调查结果见表1。
由表1可知,无患子苗木各器官的生物量积累在不同时期表现各异。在幼苗期,各器官的生物量积累表现为叶>茎>根;进入速生期后,根吸收养分的能力增强,叶片数也相应增加,使得茎的木质化程度显著加快,而根的生物量积累也逐渐超过叶。进入生长后期,根、茎的生物量积累变缓,其中根的生物量积累超过茎和叶,而叶由于温度降低,为满足其生理需要开始脱落,生物量有所下降。随着时间的推移,叶片全部落光,苗木停止生长,其生物量的积累过程也停止。
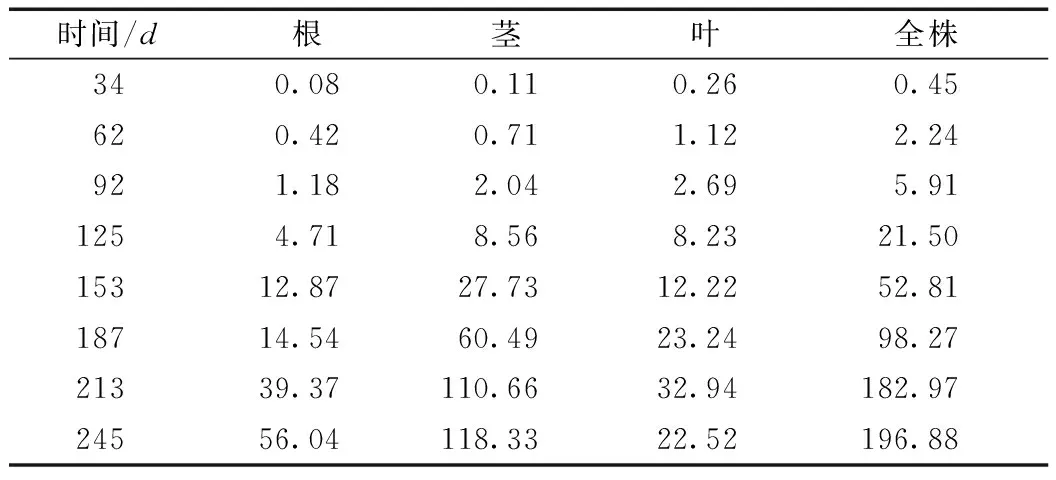
表1 无患子苗木各器官生物量的累积
3.2苗木生物量分配规律
苗木各器官生物量分配比例见表2。

表2 苗木各器官生物量分配比例
从表2可看出,无患子苗根的生物量在全年有2个生长高峰,分别为8月底(153d)和11月底(245d),说明在速生期和生长后期以地下部分的生长为主要,且生长后期分配比例上升速度超过速生期,这可能与当地的气候和无患子本身的生物学特性有关。茎生物量分配比例总体上处于上升趋势,在4月底(34d)分配比例最小,随后迅速增长,9月底(187d) 生物量分配比例达到最大值,之后略有下降。叶生物量所占比例在4月底时(34d)最大,之后一直持续下降直至停止生长。纵观整个苗期叶生物量分配比例的变化,其在幼苗期所占的比例均较大,说明无患子在幼苗期以叶生长为主。综上所述,在幼苗期,生物量的积累主要分布在生产器官叶中。进入速生期后,生物量的积累中心逐渐转移到根、茎,而茎是生物量积累的最中心。进入生长后期,根、茎进一步木质化,而叶片脱落生物量分配比例骤减。从苗木地上与地下部分生物量分配比例来看,在整个生长期,地上部分生物量积累显著高于地下部分,而这部分贡献主要来自于茎,说明无患子幼苗在生长过程中,光合产物优先运输到茎中。
3.3根、茎生物量积累的异速现象
无患子苗的根、茎生物量在不同时期表现出不一致的积累速度,这一现象即为异速生长。为此,利用SPSS18.0曲线估计中的幂函数数学模型将不同生长期根、茎之间的生物量(表1)进行拟合,得到两器官间的异速生长方程根、茎易速生长方程及回归假设检验结果见表3。

表3 根、茎异速生长方程及回归假设检验结果
由表3可知,无患子的根、茎异速生长方程决定系数达0.991,方差分析表明两者间存在着极显著的回归关系,且异速生长常数B值小于1。
3.4生物量积累与地径、苗高的回归关系
为进一步揭示无患子在苗期各器官生物量积累与地径、苗高之间的回归关系,将不同生长时期苗木全株及各器官的生物量与相应时期地径、苗高的生长量进行回归分析。经SPSS回归分析、计算、筛选,发现模型W=0.005(D2H)0.849比其他模型更适合于无患子苗木生物量的估测(表4)。由表4可知,无患子苗木各器官生物量积累与地径、苗高回归关系极显著,方程拟合效果好,能够较准确地估测出无患子各器官的生物量。
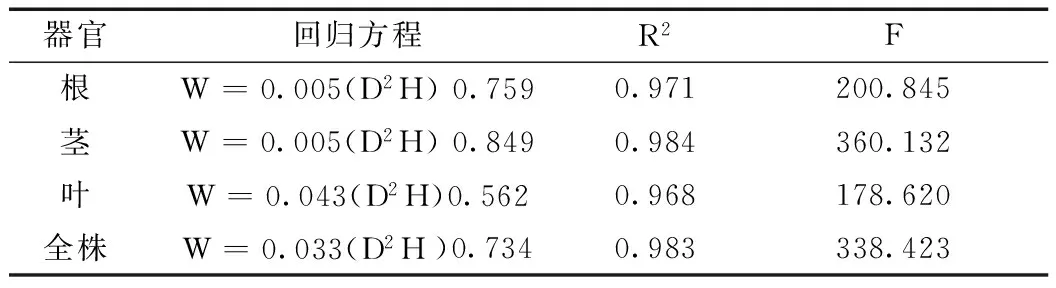
表4 无患子生物量回归方程
4 结 语
对无患子1年生播种苗生物量积累分配规律的研究发现,苗木根生物量分配比例在全年有2个生长高峰,分别为8月底和11月底。茎生物量分配总体上处于上升趋势,且一直超过根,在4月底生物量分配比例最小,随后迅速增长,在9月底时生物量分配比例达到最大值,之后略有下降,但变化不大,直至生长结束时生物量占全株的60.1%。叶生物量分配比例在幼苗期最大,速生期与根基本持平,生长后期最小,最终在外界环境(如风吹、雨打、霜冻等)和自身生理活动作用下全部脱落。无患子当年生播种苗前期侧重于茎的高生长和增加叶面积,有利于增强对有限光资源的吸收和利用能力,前期这种生长特性使苗木在适应外界环境过程中能够率先占据有利生态位。当进入秋冬时节,温度一般较低,气候较干旱土壤易缺水,叶片中光合同化产物向根部运输比例增大,导致叶生物量减少,但苗木地下部分生长加强,有利于苗木根系的深扎和扩大根表面积,增强对土壤有限的水、养分资源的吸收和利用能力,保证苗木安全越冬。
前人研究[21-23]结果表明,当根与茎生物量间的B值大于1时,植物生长中心以地下为主,而当B小于1,则生长中心以地上为主。结合根、茎异速生长模型的拟合结果表明,无患子苗期生长趋向于地上部分,这与实际的观察是一致的。对各器官生物量积累与地径、苗高回归关系的拟合结果表明,根、茎、叶及全株的生物量积累与地径、苗高的生长量有着极显著的相关性,能够较好地估测各器官生物量分配,从而揭示幼苗生长的内在规律,为更好地培育无患子壮苗提供科学依据。
[1]严成,尹林克,朱金星,等.干旱沙漠区侧柏苗木生物量的研究[J].干旱区研,1999,15(2):22-26.
[2]王月海,许景伟,韩友吉,等.黄河三角洲五个耐盐树种苗木生物量比较[J].林业科技开发,2013,23(4):52-55.
[3]陈存及.阔叶树种栽培[M].北京:中国林业出版社,2000.
[4]《中国树木志》编辑委员会.中国树木志:6卷[M].北京:中国林业出版社,2004.
[5]贾黎明,孙操稳.生物柴油树种无患子研究进展[J].中国农业大学学报,2012,17(6):191-196.
[6]YangChaohsun,HuangYuchun.Foamproperties,detergentabilitiesandlongtermpreservativeEfficacyofthesaponinsfromSapindus mukorossi[J].JournalofFoodandDrugAnalysis,2010,18(3):155-160.
[7]AparnaUpadhyay,SinghDK.PharmacologicaleffectsofSapindusmukorossi[J].RevInstMedTropSaoPaulo,2012,54(5):273-280.
[8]张海蓉,蔡宇杰,廖祥儒,等.无患子提取物对Fusariumsp.发酵产脂肪酶的影响及其双水相萃取分离[J].食品与生物技术学报,2013,30(6):928-933.
[9]邵文豪,刁松,姜景民,等.基于种实性状的无患子天然群体表型多样性研究[J].林业科学研究,2014,34(6):1452-1460.
[10]邵文豪,刁松,姜景民,等.无患子实生群体种实表型性状变异研究[J].西北农林科技大学学报(自然科学版),2014,42(5):75-83.
[11]MaharKS,RanaTS,RanadeSA.Molecularanalysesofgeneticvariabilityinsoapnut(Sapindus mukurossiGaertn.)[J].IndCropsProd,2011,34(1):1111-1118.
[12]彭珠清,范辉华,刘宝,等.无患子SRAP-PCR反应体系的优化[J].浙江农林大学学报,2014,31(2):322-328.
[13]邵文豪,刁松,董汝湘,等. 无患子果实发育动态及内含物含量变化[J].生态学报,2014,27(5):697-701.
[14]邵文豪,刁松,董汝湘,等. 无患子光合生理日变化及其与生理生态因子的关系[J].西北植物学报,2014,34(4):828-834.
[15]闫瑞,钱春.无患子幼苗对水分胁迫的生理响应[J].西南大学学报(自然科学版),2014,36(4):29-33.
[16]范辉华,张天宇,姚湘明,等.高产油脂无患子优树的选择[J]. 中南林业科技大学学报,2014,34(5):4-8.
[17]潘良田,夏尚光,丁增发.皖东地区无患子大苗培育试验及苗期生长规律研究[J].安徽农学通报,2013,19(16):107-108.
[18]陈存及,刘春霞,陈长雄,等.乳源木莲苗木生物量的研究[J].福建林学院学报,2004,24(2):107-110.
[19]张纪卯.毛红椿1年生播种苗各器官生长量生物量变化规律[J].西南林业大学学报,2011,31(1):11-16.
[20]郑益兴,彭兴民,赵保荣,等.印楝实生苗年生长规律研究[J].林业科学研究,2006,19(2):182-187.
[21]郎华安,黄粮增,顾介安.柳杉苗木生物量初步研究[J].福建林学院学报,1990,10(4):427-432.
[22]林生明,徐土根,周国模.杉木人工林生物量的研究[J].浙江林学院学报,1991,8(3):288-294.
[23]周国模,吴家胜,应叶青,等.喜树种源苗期生物量研究[J].林业科学研究,1999,12(4):386-391.
(责任编辑韩明跃)
The Research on Biomass Accumulation and Distribution Laws of Sapindus mukorossiinSeedlingStage
Zhou Shiyong
(Forestry Bureau of Jianyang in Fujian Province, Jianyang Fujian 354200, China)
Inthispaper,theseedlingbiomassofSapindus mukorossiofaccumulationanddistributionlawswasanalyzedquantitatively,thegrowthallometricequationofseedlingrootandstembiomasswasformulated,andthecorrelationofeachorgan,wholeplantbetweenseedlingheight,grounddiameterwasdescribedbymeansofSPSSregressionanalyzing.Theresultsshowedthat:theannualbiomassaccumulationoftheone-year-oldSapindus mukorossiseedlingcouldbedividedintothreestages,includingtheseedlingstage,fast-growingperiod,laterstage.Theallocationproportionofrootbiomasswastheleastinseedlingstage,butleafwasthelargest,respectivelyaccountingfor16.85 %and58.80 %ofthewholeplant.Enteringthefast-growingperiod,thecenteraccumulationofbiomassgraduallytransferredtotherootsandstems,thestembecomethemostcenterofbiomassaccumulation,accountingfor61.55 %ofthetotalplantbiomass.Atthelaterstageofgrowth,rootandstemwerefurtherlignified,thebiomassallocationproportionofleafreducedsharplybecauseleavesfalling,onlyaccountingfor11.44 %.Thedifferentspeedofbiomassaccumulationofrootandstem,andseedlingbiomassofeachorgans,wholeplant,andgrounddiameter,heightgrowthbothshowedextremelysignificantcorrelation,themathematicalmodelwashighreliability,couldbeusedtoguidethepracticalproduction.
Sapindus mukorossi;biomass;accumulation;distributionlaws;mathematicalmodel

2015-03-20
南平市科技局专项基金资助项目(IV2009244)。
周世勇(1967—),男,工程师。研究方向:森林培育。Email:693414922@qq.com。
10.11929/j.issn.2095-1914.2016.01.024
S781.55
A
2095-1914(2016)01-0140-04
