前扣带皮层区域磷酸激酶Czeta在弗氏安全佐剂致炎性痛大鼠情绪反应中的作用
2016-09-13杜俊英温存邵晓梅房军帆梁宜方剑乔
杜俊英,温存,邵晓梅,房军帆,梁宜,方剑乔
(浙江中医药大学 第三临床医学院 针灸神经生物学实验室,杭州 310053)
前扣带皮层区域磷酸激酶Czeta在弗氏安全佐剂致炎性痛大鼠情绪反应中的作用
杜俊英Δ,温存Δ,邵晓梅,房军帆,梁宜,方剑乔*
(浙江中医药大学 第三临床医学院 针灸神经生物学实验室,杭州310053)
目的观察CFA致炎性痛模型大鼠的情绪反应,探讨前扣带皮层(ACC)磷酸激酶Czeta(PKCzeta)与炎性痛大鼠情绪反应的关系。方法24只清洁级雄性SD大鼠随机分为空白对照组和模型对照组。足底皮下注射弗氏完全佐剂建立慢性炎性痛模型。动态观察各组大鼠造模前(base)、造模后3、7、14、21和28d的体重和痛阈变化;观察造模后14、21、28d所有大鼠在高架O迷宫中的总运动距离、开放臂运动距离、开放臂进入次数和开放臂停留时间百分比。观察造模后14、29d所有大鼠在旷场中的总运动距离、中央象限运动距离、中央象限进入次数和中央象限停留时间。采用免疫印迹法检测造模后14d和29d健侧和患侧ACC区域PKCzeta蛋白表达。结果 各个时间点两组大鼠体重差异无显著性(P>0.05)。造模前,两组大鼠痛阈差异无显著性(P>0.05);造模后1d,模型对照组大鼠痛阈显著降低(P<0.05),且在整个实验过程中均显著低于空白对照组(P<0.05)。与空白对照组比较,模型对照组大鼠于模后28d开放臂运动距离和开放臂停留时间百分比显著减少(P<0.05),于模后29d中央象限运动距离和中央象限进入次数明显减少(P<0.05)。造模后29d,模型对照组大鼠患侧ACC区域PKCzeta蛋白表达明显多于空白对照组(P<0.05)。且开放臂运动距离、开放臂停留时间百分比、中央象限运动距离、中央象限进入次数与患侧PKCzeta蛋白表达变化呈负相关。结论CFA诱导的慢性炎性痛大鼠可出现异常情绪行为;慢性炎性痛情绪样行为可能与ACC区域PKCzeta的高表达相关。
炎性痛;情绪;前扣带皮层;磷酸激酶Czeta;弗氏安全佐剂;大鼠
疼痛是一种与组织损伤或潜在的损伤相关的不愉快的主观感觉和情感体验。根据这个定义可以看出疼痛是由互不相同而又互相联系的多种成分组成:分辨成分(sensorydiscriminative)-感觉,动机成分(affectivemotivational)-情绪和评估成分(cognitiveevaluational)-认知。分辨成分-疼痛感觉的研究已经取得长足的进展,但动机成分-痛情绪的研究相对滞后。越来越多的证据表明慢性疼痛病人经历的痛情绪折磨比疼痛本身更严重[1, 2]。因此,近年对疼痛相关情绪的机制研究成为临床和基础研究的热点。
研究认为内侧痛传导系统通路与疼痛导致的不良情绪的信息处理相关[3]。前扣带皮层是内侧痛传导系统通路的重要组成部分,是痛情绪处理的高级中枢。神经科学研究发现谷氨酸在焦虑情绪中起着重要的调节作用[4],PKC作为谷氨酸活化的关键物质,在情绪反应中的作用也逐渐被发现,已有文献报道,敲除PKCgamma和PKCepsilon基因能够抑制焦虑样情绪反应[5, 6],PKCzeta作为非典型PKC家族的一员是否与情绪样变化相关,目前未见报道。本研究通过动态观察CFA诱导的慢性炎性痛不同时期痛阈和相关情绪反应变化情况,明确慢性炎性痛诱导痛情绪的时间;同时观察前扣带皮层PKCzeta的蛋白表达情况,探讨慢性炎性痛诱导痛情绪的可能中枢机制。
1 材料与方法
1.1材料
1.1.1实验动物
选用清洁级雄性SD大鼠24只,体重280~300g,2月龄。均购于中国科学院上海实验动物中心【SCXK(沪)2013-0016】,由浙江中医药大学实验动物中心饲养。饲养期间给予啮齿类动物标准颗粒饲料(由实验动物中心提供)及自由饮水,12h循环灯光,恒定湿度,室温(23±2)℃。实验大鼠饲养及组织取材均于浙江中医药大学实验动物中心实验设备内进行【SYXK(浙)2013-0184】。本实验所有操作均符合中华人民共和国《实验动物管理条例》。
1.1.2仪器与试剂
弗氏完全佐剂(CFA,Sigma公司,美国),PKCzeta(Abcam,美国),GAPDH(CST,美国),vonFrey丝套件(Thermo,美国),旷场(瑞沃德,中国),高架O迷宫(瑞沃德,中国),Smart3.0分析软件(Panlab,美国),ImageQuantLAS4000(Thermo,美国),ImageQuantTL(Thermo,美国),Western及IP细胞裂解液(碧云天,中国),0.45μmPVDF膜(Bio-Rad,美国),兔抗大鼠PKCzeta(#ab59364,Abcam),兔抗大鼠HRP标记的GAPDH(#3683,CellSignalingTechnology),制胶器(Bio-Rad,美国),电泳仪(Bio-Rad,美国),快速凝胶转印仪(Bio-Rad,美国)。
1.2方法
1.2.1分组及造模
所有大鼠随机分为空白对照组和模型对照组各12只,模型对照组:通过左后足底皮下注射0.1mL弗氏完全佐剂(CompleteFreund’sadjuvant,CFA)建立慢性炎性痛模型。
1.2.2机械痛阈
机械痛阈测量时间固定在9:00-17:00,环境温度为23℃左右。开始前,先将大鼠放置于特定的铁丝网上(UGO),盖以透明的有机玻璃罩(20cm× 20cm× 15cm),适应环境直至大鼠安静(即停止梳理毛发和探索性活动),大约15min左右。按照Chaplan等[7]创建的经典von-Frey刺激丝(StoeltingCo,Thermo)(力量分别是0.4、0.6、1.0、2.0、4.0、6.0、8.0、15.0g和26.0g)Up-Down方法测量机械缩腿阈作为痛阈。首先从4.0g开始,将one-Frey刺激丝垂直刺向大鼠左后足足底中央区域皮肤(避开足垫),垂直用力使one-Frey刺激丝弯曲成S形。刺激时间每次持续≤8s,间隔时间>2min。若大鼠出现缩足/舔足反应为阳性反应,以“X”表示,换大一号力量的one-Frey刺激丝继续刺激;反之则以“O”表示,换小一号力量的one-Frey刺激丝继续刺激;测量6次,可得到如“OXXOXO”的序列,以(10[Xf+κδ])/10 000的公式计算痛阈。其中δ为0.231,κ值根据“O”、“X”表获得,Xf为最后一根vonFrey刺激丝的对数值。
1.2.3旷场实验
操作间保持较暗的光线,避免直射光线,恒定温度(23℃左右),安静条件下进行。测试之前先将待测实验大鼠放入实验环境中2h以适应环境。开始时将实验大鼠头部背对实验者轻轻放入旷场实验箱正中象限中,进行摄像,记录10min的大鼠活动情况。用Smart3.0软件进行数据分析:分析时将旷场划为16个格子,中间4格为中央象限,其他12格为周围象限,分析实验动物在旷场的总运动距离、中央象限运动距离、中央象限进入次数和中央象限停留时间。每只大鼠测量前均用10%酒精清洗以消除上一次大鼠留下的信息(如气味、大便、小便等)。
1.2.4高架O迷宫
环境要求及适应条件同旷场实验。开始时将实验大鼠迅速轻放置于闭合臂和开放臂的交界,要求实验大鼠头面向开放臂。记录5min大鼠在高架0迷宫中的活动情况。用Smart3.0软件进行数据分析:分析时将高架O迷宫分成4个臂,2个为闭合臂,2个为开放臂。观察并计算实验大鼠高架O迷宫总运动距离、开放臂运动距离、开放臂进入次数和开放臂停留时间百分比。每只大鼠测量前均用10%酒精清洗以消除上一次大鼠留下的信息(如气味、大便、小便等)。1.2.5免疫印迹法检测ACC区域PKCzeta蛋白表达各组大鼠于造模后14、29d检测完行为学指标后,以0.35mL/ 0.2kg的10 %水合氯醛腹腔麻醉,冰冻生理盐水(4℃)灌注,参照Paxinos-Watson大鼠脑立体定位图谱快熟取得左右两侧ACC放入Western及IP细胞裂解液冰上超声粉碎,其中功率为200W,工作2s,间隔6s,15次;15 000g离心取上清液,BCA法定量蛋白浓度,取20μg蛋白上样,以5%~10%SDS-PAGE凝胶电泳分离蛋白(80V,30min,再150V,90min),然后通过半干转印法将蛋白转印至PVDF膜(0.45μm,Bio-Rad),10%脱脂奶粉室温封闭1h,分别用兔抗大鼠PKCzeta、兔抗大鼠HRP标记的GAPDH一抗孵育过夜(4℃),HRP标记的山羊抗兔二抗孵育1h(室温),ECL试剂盒显色,采用ImageQuantLAS4000系统拍照,ImageQuantTL系统计算条带的平均光密度值,其中GAPDH为内部对照。
1.3统计分析
2 结果
2.1各组大鼠体重变化情况
我们动态观察了空白对照组和模型对照组大鼠造模前、造模后3、7、14、21d和28d体重变化,结果如图1所示,在整个实验的各个时间点,两组大鼠体重未见明显差异(P>0.05)。
2.2vonFrey刺激丝up-down法观察慢性炎性痛大鼠造模后不同时间点痛阈变化情况
我们动态观察各组大鼠造模前、造模后1、3、7、14、21d和28d的患侧足跖痛阈变化。造模前,空白对照组和模型对照组大鼠患侧足跖痛阈差异无显著性,具有可比性(P>0.05)。造模后1d,模型对照组大鼠患侧足跖痛阈明显低于同期空白对照组(P<0.01),提示成功诱发CFA大鼠患侧足跖痛觉异常;造模后3、7、14、21d和28d,模型对照组大鼠患侧足跖痛阈均显著低于空白对照组大鼠(P<0.01)(详见图2)。
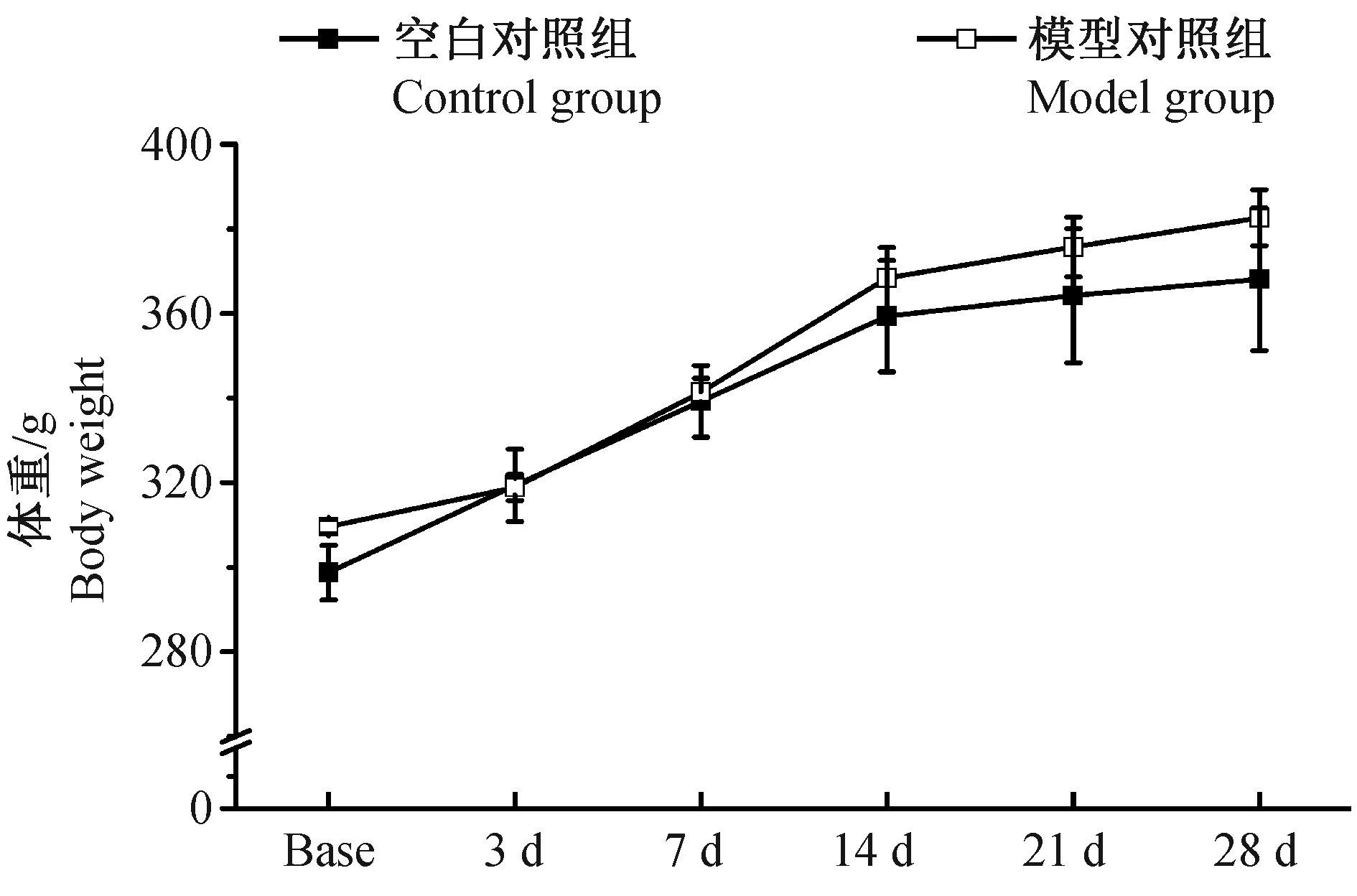
图1 各组大鼠痛体重变化情况±s,g)Fig.1 Changes of the body weight of rats in each group at different ±s,g)
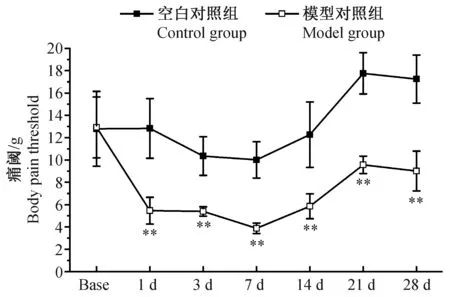
注:与空白对照组比较:*P<0.05,**P<0.01。图2 各组大鼠痛阈变化情况±s,g)Note. Compared with the control group, *P<0.05,**P<0.01.Fig.2 Changes of the pain threshold of rats in each group at different ±s,g)
2.3高架O迷宫实验观察慢性炎性痛大鼠造模后不同时间点的行为学变化情况
我们采用高架O迷宫实验观察慢性炎性痛大鼠造模后14、21d和28d的行为学变化情况,观察5min内实验大鼠在高架O迷宫中的总运动距离,开放臂运动距离,开放臂进入次数和开放臂停留时间百分比。结果如图3显示,造模后14、21d,空白对照组和模型对照组大鼠的总运动距离、开放臂运动距离、开放臂进入次数和开放臂停留时间百分比差异均无显著性(P>0.05);造模后28d,模型组大鼠开放臂运动距离和开放臂停留时间百分比显著小于空白对照组(P<0.05),两组大鼠总运动距离和开放臂进入次数无差异(P>0.05)。
2.4旷场实验观察慢性炎性痛大鼠造模后不同时间点的行为学变化情况
我们采用旷场实验观察造模后14、29d的焦虑情绪行为情况,观察10min内实验大鼠在旷场中的总运动距离,中央象限运动距离,中央象限进入次数和中央象限停留时间。结果如图4显示,造模后14d,空白对照组和模型对照组大鼠的总运动距离、中央象限运动距离、中央象限进入次数和中央象限停留时间百分比差异均无显著性(P>0.05);造模后29d,模型组大鼠中央象限运动距离和中央象限进入次数显著小于空白对照组(P<0.05),中央象限停留时间减少但与空白组差异无显著性(P>0.05)。
2.5慢性炎性痛大鼠造模后不同时间点的ACC区域p-PKCzeta蛋白表达情况
我们采用免疫印迹法观察慢性炎性痛大鼠造模后14、29d健侧和患侧ACC区域PKCzeta蛋白表达情况,结果如图5所示。造模后14d,空白对照组和模型对照组大鼠健侧和患侧ACC区域PKCzeta蛋白表达差异无显著性(P>0.05)。造模后29d,与空白对照组比较,模型对照组大鼠患侧ACC区域PKCzeta蛋白表达明显增多(P<0.05),其健侧PKCzeta蛋白表达差异无显著性(P>0.05)。
2.6慢性炎性痛大鼠情绪样变化与患侧ACC区域PKCzeta的相关分析
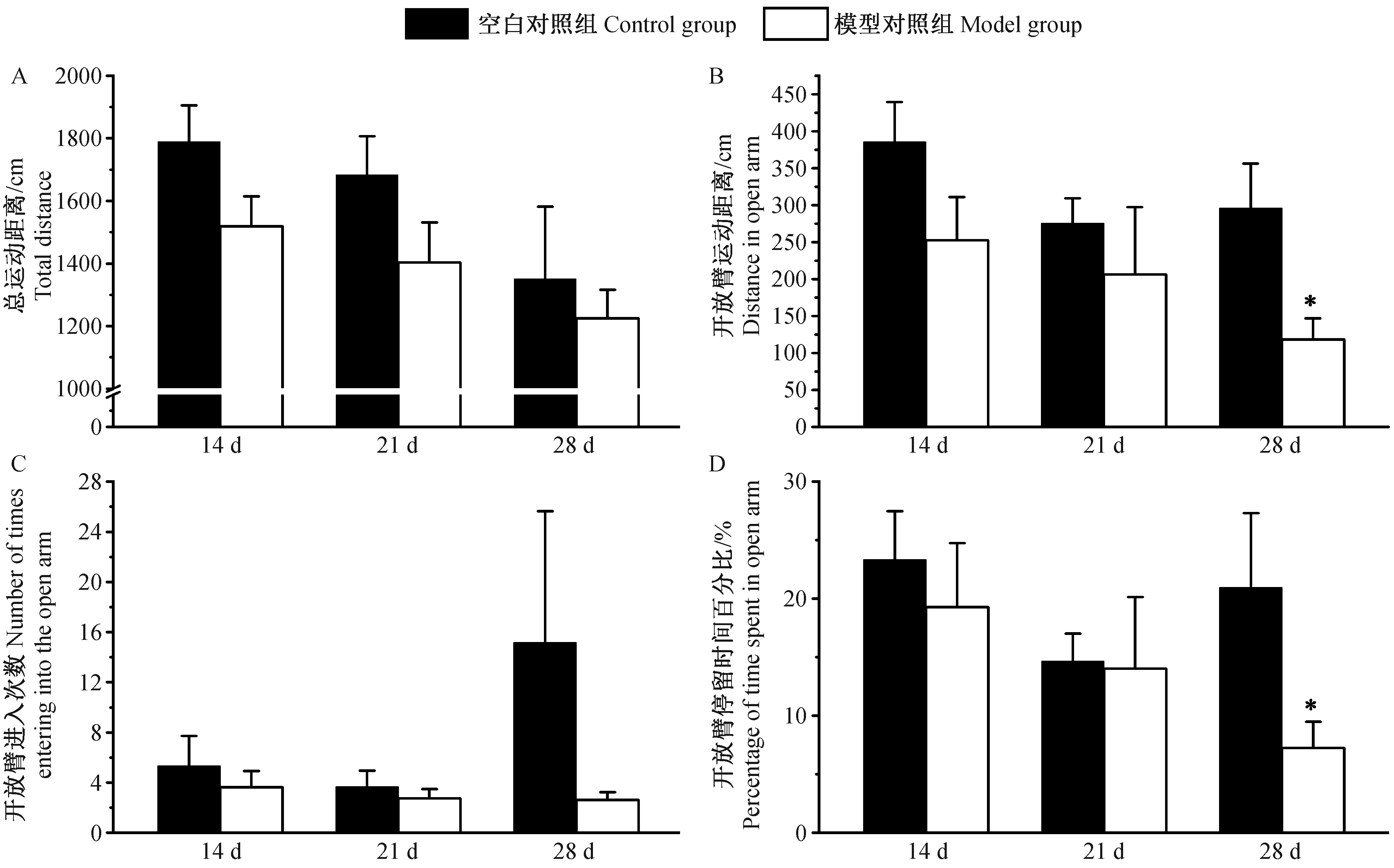
图3 各组大鼠高架O迷宫行为学变化情况±s)Fig.3 Changes of the results of elevated zero maze test of the rats in each group at different time-points

图4 各组大鼠旷场行为学变化情况±s)Fig.4 Changes of the results of open field test of rats in each group at different time-points
为了探讨患侧ACC区域PKCzeta蛋白变化与慢性炎性痛情绪样变化是否相关,我们分别将实验大鼠开放臂运动距离与PKCzeta蛋白表达、开放臂停留时间百分比与PKCzeta蛋白表达、中央象限运动距离与PKCzeta蛋白表达、中央象限进入次数与PKCzeta蛋白表达进行了相关分析检测,结果如图6所示。开放臂运动距离与PKCzeta蛋白表达之间的Pearson相关系数为-0.467,P<0.01;开放臂停留时间百分比与PKCzeta蛋白表达之间的Pearson相关系数为-0.413,P<0.05;中央象限运动距离与PKCzeta蛋白表达之间的Pearson相关系数为-0.570,P<0.01;中央象限进入次数与PKCzeta蛋白表达之间的Pearson相关系数为-0.543,P<0.01。结果提示PKCzeta蛋白表达变化与开放臂运动距离、开放臂停留时间百分比、中央象限运动距离、中央象限进入次数均呈负相关,相关性比较强。因此,患侧ACC区域PKCzeta蛋白表达变化与慢性炎性痛情绪样变化密切相关。
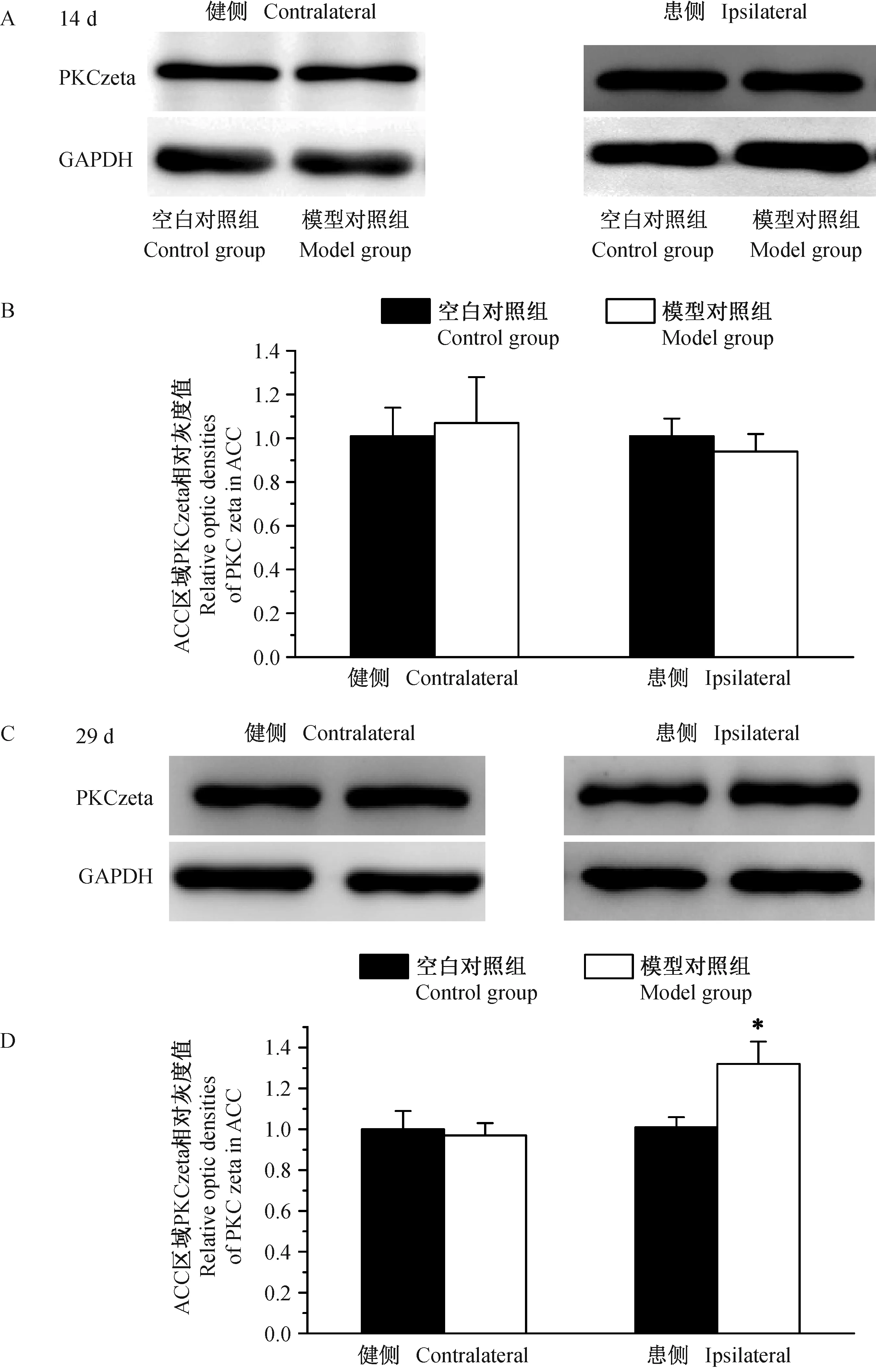
图5 各组大鼠ACC脑区PKCzeta变化情况Fig.5 Changes of the PKCzeta protein expression in ACC of rats in each group at different time-points

图6 慢性炎性痛情绪样行为与患侧PKCzeta蛋白表达的相关分析Fig.6 Correlation analysis between the chronic inflammation-induced pain emotion and PKCzeta protein expression in the ipsilateral ACC
3 讨论
疼痛包含感觉成分和情绪成分。痛的感觉成分编码伤害性刺激的性质、躯体定位、强度和空间特性;痛的情绪成分编码痛情绪、唤醒以及引起逃避行为时的强迫及指向特性。情绪折磨被认为是疼痛的基本特性。常用的痛感觉成分检测方法为冷、热、机械痛阈,常用的痛情绪成分观察方法为旷场、O迷宫等。基于对疼痛认识的不断深入,基础研究者开始采用情绪测量方法和手段观察常见慢性疼痛模型的相关不良情绪及可能机制。
通过足底皮下或关节腔内注射弗氏完全佐剂建立的炎性痛动物模型是最常见的慢性疼痛模型之一,其造模简单,易于复制,广受研究者喜爱,常用于慢性炎性痛痛感觉治疗及机制研究。过往研究认为,慢性炎性痛模型不会诱导痛情绪,但近年来,有研究指出弗氏完全佐剂诱导的慢性炎性痛模型伴有痛情绪反应,且开始采用慢性炎性痛模型探讨痛情绪的相关机制。Borges等[8]采用关节腔内注射弗氏完全佐剂诱导的炎性痛大鼠模型中发现,造模后14d和28d,模型大鼠埋珠个数明显增多;造模后28d,模型大鼠开放臂停留时间百分比明显减少,强迫游泳不动时间明显增多。Grégoire等[9]采用相似的方法在造模后21d也观察到模型大鼠的痛情绪反应。Parent等[10]也观察到弗氏完全佐剂造模后28~30d,模型大鼠的痛情绪行为:其社会交互活动减少、在开放臂停留时间百分比及进入次数减少、进入中央象限的次数减少。Narita等[11]采用相同试剂减半足底注射建立炎性痛大鼠模型也在造模后28d观察到模型大鼠的痛情绪反应。我们课题组也在造模后28d观察到弗氏完全佐剂诱导的慢性炎性痛模型的痛情绪行为。因此,CFA诱导的慢性炎性痛大鼠模型可出现痛情绪行为。
大量研究证实,ACC是疼痛处理的关键脑区[12]。ACC不仅直接接受丘脑传入的伤害性信息,间接接受躯体感觉中枢(S1和S2)的信息,同时还汇集了来自杏仁核、海马等部位的信息,这种广泛的联系及特殊的位置是ACC参与痛情绪调节的解剖基础。早期临床研究发现,切除ACC能有效解除疼痛患者的不良情绪反应(如焦虑、抑郁)。近年来实验研究观察到发现化学损毁ACC的头端,可抑制福尔马林条件位置回避痛情绪动物模型的痛情绪反应[13-14]。
蛋白激酶C(PKC)广泛分布于大脑的各个区域,在调节神经元兴奋性、神经递质释放、基因表达长期改变和突触可塑性变化中起着重要作用[15]。PKC家族至少包括11个亚型,其中8个为典型PKC家族,包括PKCalpha、PKCbeta1、PKCbeta2、PKCgamma、PKCdelta、PKCepsilon、PKCthêta、PKCeta;3个为非典型PKC家族,其中PKCzeta因其与人的PKClota和鼠的PKClambda结构相似,共同组成非典型PKC家族[16]。目前大量的研究证明,中枢PKC的活化与不良情绪有关[15, 17-18]。有学者报道焦躁症患者和实验大鼠的大脑组织中均观察到PKC活性增加[17-18]。亦有文献报道,组氨酸三核苷酸结合蛋白1(Hint1)敲除诱发的情绪异常小鼠中观察到PKCgamma的表达增加,认为PKCgamma与小鼠焦虑和压力相关情绪相关。Bowers等[19]采用高架十字迷宫、黑白箱等观察PKCgamma基因突变小鼠的焦虑情绪行为,研究结果显示,PKCgamma基因突变小鼠进入开放臂的次数增多,在开放臂的停留时间延长,结果提示,PKCgamma与小鼠的焦虑行为相关。VanKolen等[20]报道前额叶皮质PKCepsilon的兴奋性与焦虑样情绪相关。那么,PKCzeta是否焦虑抑郁情绪相关,目前未见报道。本实验研究观察到,造模后28d模型组大鼠开放臂运动距离和开放臂停留时间百分比显著减少;造模后29d中央象限运动距离和中央象限进入次数明显减少,患侧ACC区域PKCzeta蛋白表达明显增多;同时,开放臂运动距离、开放臂停留时间百分比、中央象限运动距离、中央象限进入次数与患侧PKCzeta蛋白表达变化呈负相关。所以我们认为,慢性炎性痛情绪行为可能与ACC区域PKCzeta的高表达相关。
[ 1 ]BairMJ,RobinsonRL,KatonW,etal.Depressionandpaincomorbidity:aliteraturereview[J].ArchInternMed, 2003, 163(20): 2433-2445.
[ 2]DoanL,MandersT,WangJ.Neuroplasticityunderlyingthecomorbidityofpainanddepression[J].NeuralPlast, 2015, 2015: 504691.
[ 3 ]VogtBA,SikesRW.Themedialpainsystem,cingulatecortex,andparallelprocessingofnociceptiveinformation[J].ProgBrainRes, 2000, 122: 223-235.
[ 4 ]BerginkV,vanMegenHJ,WestenbergHG.Glutamateandanxiety[J].EurNeuropsychopharmacol, 2004, 14(3): 175-183.
[ 5 ]BarbierE,WangJB.Anti-depressantandanxiolyticlikebehaviorsinPKCI/HINT1knockoutmiceassociatedwithelevatedplasmacorticosteronelevel[J].BMCNeurosci, 2009, 10: 132.
[ 6 ]XiaoZ,JaiswalM,DengPY,etal.RequirementofphospholipaseCandproteinkinaseCincholecystokinin-mediatedfacilitationofNMDAchannelfunctionandanxiety-likebehavior[J].Hippocampus, 2012, 22(6): 1438-1450.
[ 7 ]ChaplanSR,BachFW,PogrelJW,etal.Quantitativeassessmentoftactileallodyniaintheratpaw[J].JNeurosciMethods, 1994, 53(1): 55-63.
[ 8 ]BorgesG,NetoF,MicoJA,etal.Reversalofmonoarthritis-inducedaffectivedisordersbydiclofenacinrats[J].Anesthesiology, 2014, 120(6): 1476-1490.
[ 9 ]GrégoireS,WattiezAS,EtienneM,etal.Monoarthritis-inducedemotionalandcognitiveimpairmentsinratsaresensitivetolowsystemicdosesorintra-amygdalainjectionsofmorphine[J].EurJPharmacol, 2014, 735: 1-9.
[10]ParentAJ,BeaudetN,BeaudryH,etal.Increasedanxiety-likebehaviorsinratsexperiencingchronicinflammatorypain[J].BehavBrainRes, 2012, 229(1): 160-167.
[11]NaritaM,KanekoC,MiyoshiK,etal.Chronicpaininducesanxietywithconcomitantchangesinopioidergicfunctionintheamygdala[J].Neuropsychopharmacology, 2006, 31(4): 739-750.
[12]FuchsPN,PengYB,Boyette-DavisJA,etal.Theanteriorcingulatecortexandpainprocessing[J].FrontIntegrNeurosci, 2014. 8: 35.
[13]CaoH,GaoYJ,RenWH,etal.Activationofextracellularsignal-regulatedkinaseintheanteriorcingulatecortexcontributestotheinductionandexpressionofaffectivepain[J].JNeurosci, 2009, 29(10): 3307-3321.
[ 14 ]YanN,CaoB,XuJ,etal.Glutamatergicactivationofanteriorcingulatecortexmediatestheaffectivecomponentofvisceralpainmemoryinrats[J].NeurobiolLearnMem, 2012, 97(1): 156-164.
[ 15 ]MacDonaldJF,KotechaSA,LuWY,etal.ConvergenceofPKC-dependentkinasesignalcascadesonNMDAreceptors[J].CurrDrugTargets, 2001, 2(3): 299-312.
[ 16 ]Mochly-RosenD,DasK,GrimesKV.ProteinkinaseC,anelusivetherapeutictarget? [J].NatRevDrugDiscov, 2012, 11(12): 937-957.
[ 17 ]SzaboST,Machado-VieiraR,YuanP,etal.GlutamatereceptorsastargetsofproteinkinaseCinthepathophysiologyandtreatmentofanimalmodelsofmania[J].Neuropharmacology, 2009, 56(1): 47-55.
[ 18 ]ArnstenA,ManjiH.Mania:arationalneurobiology[J].FutureNeurol, 2008, 3(2): 125-131.
[ 19 ]BowersBJ,CollinsAC,TrittoT,etal.MicelackingPKCgammaexhibitdecreasedanxiety[J].BehavGenet, 2000, 30(2): 111-121.
[ 20 ]VanKolenK,PullanS,NeefsJM,etal.NociceptiveandbehaviouralsensitisationbyproteinkinaseCepsilonsignallingintheCNS[J].JNeurochem, 2008, 104(1): 1-13.
The role of PKCzeta-expression in the anterior cingulatecortexinemotionalresponsetocompleteFreund’sadjuvant-inducedinflammatorypain
DUJun-yingΔ,WENCunΔ,SHAOXiao-mei,FANGJun-fan,LIANGYi,FANGJian-qiao*
(DepartmentofNeurobiologyandAcupunctureResearch,TheThirdClinicalMedicalCollege,ZhejiangChineseMedicalUniversity,Hangzhou310053,China)
ObjectiveToobservetheemotionalresponsetoinflammatorypainmodelinducedbycompleteFreund’sadjuvant(CFA),andtoexploretherelationshipbetweenphosphatekinaseC(PKC)zetaintheanteriorcingulatecortex(ACC)andemotionalresponsetoinflammatorypain.MethodsTwenty-fourhealthy2-montholdfemaleSprague-Dawleyratswererandomlydividedintocontrolgroupandmodelgroup.Theinflammatorypainmodelwasestablishedbyinjectingsubcutaneouslywith0.1mlCFAintotherighthindpawofrats.Toobservethedynamicalchangesofbodyweightandpainthresholdbeforemodeling,andat3, 7, 14, 21,and28daysaftermodeling.Detectingthetotaldistance,distanceinopenarm,thenumberoftimesenteringintotheopenarm,percentageoftimespentinopenarmofelevatedOmazeweremeasuredat14, 21, 28daysaftermodeling.Thetotaldistance,distanceinthecentralquadrat,thenumberoftimesenteringthecentralquadrat,andtimespentinthecentralquadratofopenfieldweredeterminedat14and29daysaftermodeling.TheproteinexpressionofPKCzetaintheACCwasdetectedbyWesternblotat14and29daysafterCFA-injection.ResultsTherewerenosignificantdifferencesofbodyweightbetweenthecontrolandmodelgroups(P>0.05).Comparedwiththecontrolgroup,thepainthresholdofratsinthemodelgroupwassignificantlydecreasedat1, 3, 7, 14, 21,and28daysafterCFA-injection(P<0.05),thedistanceinopenarmandpercentageoftimespentinopenarmofthemodelgroupweremarkedlyreducedat28daysaftermodeling(P<0.05),distanceinthecentralquadratandthenumberoftimesenteringintothecentralquadratofmodelgroupweresignificantlydecreasedat29daysaftermodeling(P<0.05),andtheproteinexpressionofPKCzetainipsilateralACCwasincreasedat29daysafterCFA-injection(P<0.05).TherewasanegativecorrelationbetweendistanceinopenarmandproteinexpressionofPKCzetaintheipsilateralACC,percentageoftimespentinopenarmandproteinexpressionofPKCzetaintheipsilateralACC,distanceinthecentralquadratandproteinexpressionofPKCzetainipsilateralACC,thenumberoftimesenteringintothecentralquadrantandproteinexpressionofPKCzetaintheipsilateralACC,respectively.ConclusionsAbnormalemotionalbehaviormayoccurinratswithchronicinflammationpaininducedbyCFA-injection.ThispainemotionbehaviormaybeassociatedwithhighproteinexpressionofPKCzetaintheipsilateralanteriorcingulatecortex.
Inflammatorypain;Emotion;Anteriorcingulatecortex;CompleteFreund’sadjuvant;PKCzeta;Rat
FANGJian-qiao.E-mail:fangjianqiao7532@163.com
浙江省自然科学基金项目(编号:LQ15H270003);浙江省医药卫生科技计划项目(编号:2015KYB281);浙江中医药大学校级科研基金项目(编号:2014ZR08);浙江中医药大学附属第三医院医药卫生科研计划项目(编号:ZS14ZA01)。
杜俊英(1984-),女,助理研究员,研究方向:针灸调控痛情绪的效应及机制研究。Email:dujunying0706@163.com。温存(1990-),女,硕士研究生,专业:针灸推拿学。Email:1140121113@qq.com。Δ共同第一作者
方剑乔(1961-),男,教授,研究方向:针灸镇痛与免疫调节的相关性研究。Email:fangjianqiao7532@163.com
研究报告
Q95-33
A
1005-4847(2016)04-0414-08
10.3969/j.issn.1005-4847.2016.04.015
2015-12-15
