早春类短命植物生物量研究(Ⅳ)
——荷青花生物量及其模型构建
2016-09-02范春楠郑金萍
范春楠,程 岩,郑金萍,刘 彬,赵 哲
(1.北华大学林学院,吉林吉林 132013;2.吉林省松花江三湖自然保护区管理局,吉林吉林 132013)
早春类短命植物生物量研究(Ⅳ)
——荷青花生物量及其模型构建
范春楠1,程岩2,郑金萍1,刘彬1,赵哲1
(1.北华大学林学院,吉林吉林132013;2.吉林省松花江三湖自然保护区管理局,吉林吉林132013)
基于荷青花在药用、观赏以及森林生态系统养分循环等方面存在的价值,利用全株收获法采样,经分选、烘干、称重和数据统计后,获得了荷青花生物量及分配特征,并以其易测生长指标为自变量构建了总生物量的一元和多元线性回归方程.结果表明:荷青花群丛在营养生长停止期,各生长指标及生物量分配在个体间有较大差异,但平均丛高和根冠比变化较小;地下部分所支持的丛幅冠、丛株数和花果数等存在一定的供给限制,且这种限制与地下生物量间存在着显著或极显著的正相关关系.虽然荷青花生物量随各生长指标的变化存在不同程度的差异,但根冠比的变化差异均不显著,这在一定程度上印证了荷青花存在权衡有限光合资源分配的能力.荷青花生物量模型以多元线性回归的拟合结果较好,但考虑自变量过多带来的系统误差,生物量预估模型可选用以丛株数和花果数为自变量的拟合方程.
早春类短命植物;荷青花;生物量;分配比例;模型构建
【引用格式】范春楠,程岩,郑金萍,等.早春类短命植物生物量研究(Ⅳ):荷青花生物量及其模型构建[J].北华大学学报(自然科学版),2016,17(3):308-314.
荷青花(Hylomecon japonica)别名鸡蛋黄花(东北)、刀豆三七、水菖兰七(浙江)、拐枣七、大叶老鼠七等,为罂粟科(Papaveraceae)荷青花属(Hylomecon)多年生早春类短命植物,高15~40 cm,根茎斜生,茎直立,花期4—5月,果期5—6月;产于我国东北至华中、华东地区,在朝鲜、日本及俄罗斯西伯利亚也有分布[1-2].荷青花多以根茎入药,其药用价值在《中药辞海》《中华本草》《浙江药用植物志》及《中国长白山药用植物彩色图志》等多部志书中均有记载[3-6].具有祛风除湿,舒筋通络,散瘀消肿,止血镇痛之功效,用于治疗风寒湿痹、风湿关节痛、跌打损伤、劳伤、四肢乏力、胃脘痛、痢疾等[7].同时,荷青花又因花期早,花冠大,色彩艳丽,被诸多学者公认为极具开发潜力的野生花卉品种[8-11],现已有北京天坛公园丁香林下的栽培应用实例[10].
本文鉴于荷青花的药用和观赏价值以及早春植物在森林养分循环中的特殊作用,利用全株收获法采样,从其生长状况、生物分配特征以及生物量模型构建方面进行研究.研究结果可在一定程度上揭示荷青花的生存策略,为摸清其资源储量及开发利用提供重要的理论依据,同时也可在明确早春植物在森林生态系统养分循环中的作用与地位方面提供重要参考.
1 研究地概况
研究地位于吉林市丰满区前二道乡王相村境内的磨盘山,气候为北温带大陆季风气候,年平均气温4. 5℃,1月平均气温-17.9℃,最低温度-40.2℃;7月平均气温22.9℃,最高温度33.6℃.平均降水量为668 mm,无霜期130 d左右.
森林植被多为次生落叶阔叶林,主要树种包括核桃楸(Juglans mandshurica)、花曲柳(Fraxinus rhynchophylla)、紫椴(Tilia amurensis)、蒙古栎(Quercusmongolica)等;灌木植物以卫矛(Euonymus alatus)、山楂叶悬钩子(Rubus crataegifolius)、东北山梅花(Philadelphus schrenkii)等为主;草本植物种类丰富,包括丝引苔草(Carex remotiuscula)、荨麻叶龙头草(Meehania urticifolia)、水杨梅(Geum aleppicum)、铃兰(Convallaria keiskei)等.本地的荷青花主要分布于落叶阔叶林下,在土壤湿润、肥沃的沟谷和溪流两岸较为集中.
2 研究方法
受荷青花生长周期短、物候期紧凑及群丛自身生长特性等因素制约,欲采集不同时期的荷青花个体比较困难.为此,本研究选择了营养生长基本停止且各器官并存的花果期进行采样.具体方法为选取荷青花健康群丛,测定其丛冠幅(Crown Width,CW)、平均丛高(Average Height,HA)以及丛株数(Individual Number,Ni)和花果数(Number of Flower and Fruit,Nff),利用收获法挖取全株,并经单丛标记、分装后带回.取回的样品以丛为单位,按器官分选为地下部分、茎叶、花果,置于80℃的烘箱中干燥至恒重后称量各器官质量,经统计获得花果生物量(Biomass of Flower and Fruit,Bff)、茎叶生物量(Biomass of Stem and Leaf,Bsl)、地上生物量(Aboveground Biomass,Ba)、地下生物量(Underground Biomass,Bu)、总生物量(Total Biomass,BT)和根冠比(Root-Shoot ratio,Rrs).本研究于2014年4月末至5月初进行,共计采集荷青花植株样品30丛,其生长指标和生物量分布状况见表1.
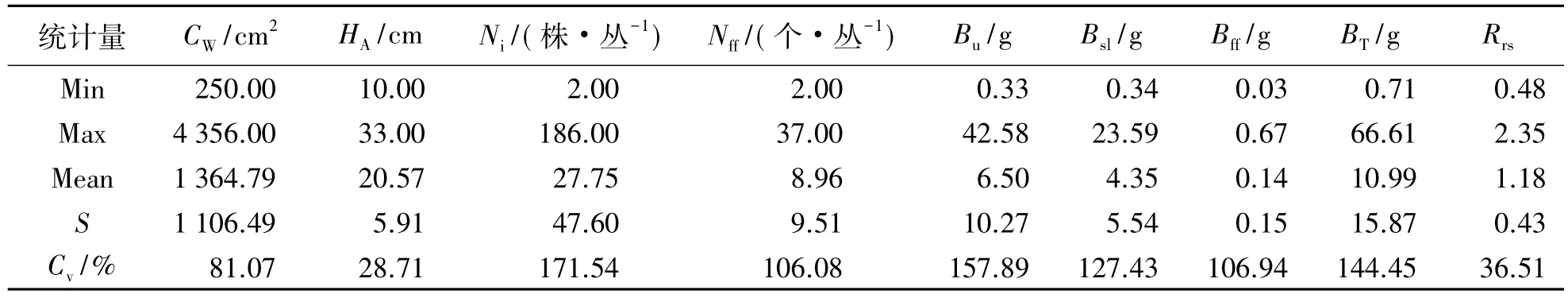
表1 荷青花群丛生长指标和生物量分布状况Tab.1 Grow th indexes and distribution of biomass of Hylomecon japonica
荷青花生物量模型的构建以丛冠幅、平均丛高、花果数、丛株数为自变量,利用SPSS软件进行相关分析,并构建一元和多元线性回归方程.
3 结果与分析
3.1生长状况
由表1可知:荷青花各生长指标在不同个体间存在较大差异,以丛株数和花果数的变幅相对最大,变异系数分别为171.54%和106.08%;其次为丛冠幅,变异系数为81.07%;平均丛高的变异相对较小,变异系数仅为28.71%.为进一步获得荷青花不同个体生长状况的分布特征,对样本的丛冠幅、平均丛高、丛株数和花果数的分布频数进行分别统计,结果见图1.
由图1可知:荷青花个体平均丛高的分布频数呈“单峰型”,在平均丛高16~20 cm时最大(0.43);随着丛冠幅、丛株数和花果数的增大、增多,荷青花个体的分布频数均呈明显下降的变化,其中丛冠幅≤1 000 cm2的样本个体占采集样本的比例略高于60%,丛株数≤10株/丛、花果数≤5个/丛的个体数量也均在50%以上,表明荷青花不同个体形态在丛冠幅面积、丛株数和花果数上存在一定的生长范围限制,且三者均与地下生物量存在显著或极显著的正相关关系(表2).
3.2生物量及其分配特征
由表1可知:荷青花各器官生物量和总生物量的变异系数均在100%以上,以地下生物量的变异最高(157.89%),其次为总生物量(144.45%)、茎叶生物量(127.43%)和花果生物量(106.94%);根冠比介于0. 48~2.35,平均为1.18,变幅相对较小,变异系数仅为36.51%.生物量及其分配比例随生长指标的变化趋势见图2和图3.
由图2可知:随丛冠幅的增加荷青花总生物量整体呈先增高后下降的趋势,在丛冠幅为2 001~3 000 cm2时最大,平均为39.33 g;茎叶生物量和地下部分生物量的变化趋势与总生物量相一致;花果生物量整体表现为随着丛冠幅的增加而增大.经方差分析发现,总生物量和各器官生物量均随丛冠幅的变化呈极显著差异(Sig.<0.000).
随平均丛高的增加总生物量、茎叶生物量和地下生物量整体也呈先增高后下降的变化趋势,均在平均丛高21~25 cm时最高,分别为23.00,14.37和8.42 g,但随株高的变化整体差异均不显著(Sig.分别为0.07 4,0.065,0.067);花果生物量随平均丛高的增加而增大,在丛高为30 cm以上时达0.32 g,随丛高的变化其生物量存在显著差异(Sig.=0.040).
随丛株数的增多总生物量、地下生物量和地上生物量呈总体增加趋势,在丛株数为60株以上时最大,分别为57.77,37.83,19.94 g,花果生物量则呈先增大后减少的变化,二者随丛株数变化的差异均极显著(Sig.<0.000).
随花果数的增加总生物量、地下生物量、茎叶生物量和花果生物量均呈明显的“单峰型”变化,且差异极显著(Sig.<0.000),最大值都出现在花果数为16~20个时,生物量分别为66.61,42.58,7.86和0.44 g.
从荷青花生物量分配比例随各生长指标的变化(图3)来看:生物量分配比例整体以地下部分略高,花果生物量平均不足总生物量的4%.其中,地下生物量的分配比例随平均丛高的增大整体增加,随花果数的增多呈明显的“单峰型”变化,但随丛冠幅和丛株数的增加变化趋势不明显;花果生物量的分配比例随丛冠幅的增大呈先下降后升高,随平均丛高和丛株数的增加整体下降,而随花果数的变化趋势不明显.经方差分析可知:根冠比的差异仅表现为随丛株数和花果数的变化差异显著(Sig.=0.030),而地下生物量、茎叶生物量和花果生物量的分配比例随各生长指标变化的差异均不显著.
3.3生物量模型构建
荷青花生物量与丛冠幅、丛株数、平均丛高及花果数的相关分析见表2.由表2可知:荷青花生物量与各生长指标的关系密切,除丛冠幅和平均丛高与根冠比的正相关关系不显著外,其余均表现为显著或极显著正相关关系.显著和极显著的相关系数介于0.424~0.951,以花果数与花果生物量、地上生物量以及丛株数与茎叶生物量、地下生物量和总生物量相对较高,均达到了0.9以上,这为荷青花生物量模型的构建提供了可能.
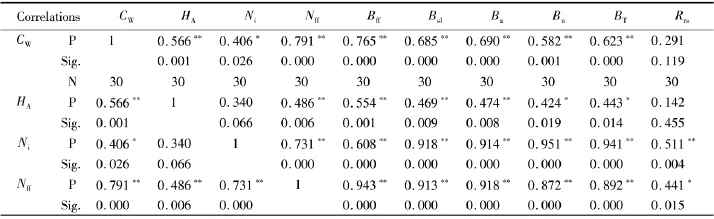
表2 荷青花生物量与各指标相关分析Tab.2 Correlations analysis am ong biom ass and grow th status of Hylomecon japonica
为此,分别以丛冠幅、平均丛高、丛株数和花果数为自变量,利用直线(y=ax+b)、指数(y=a ebx)、对数(y=a ln x+b)和幂函数(y=axb)对荷青花的总生物量进行拟合,结果见表3.
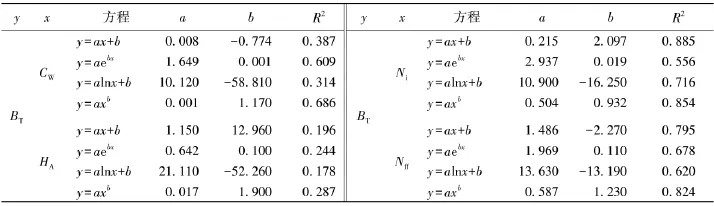
表3 荷青花总生物量与生长指标的一元线性回归Tab.3 The simple linear regression for total biomass and grow th indexes of Hylomecon japonica
由表3可知:荷青花生长指标与各总生物量的拟合方程R2介于0.178~0.885,拟合结果整体以丛株数和花果数为自变量的相对较好,拟合方程整体以幂函数较佳,但仅以丛株数为自变量的直线方程和幂函数以及以花果数为自变量的幂函数的拟合结果相关系数达到了0.8以上.
基于荷青花各生长指标均与生物量的相关关系显著以及一元线性回归方程拟合结果R2普遍较低的事实,本研究利用SPSS软件构建总生物量与生长指标的多元线性回归方程,结果见表4.
由表4可知:多元线性回归拟合方程R2在0.400~0.977,明显高于一元线性回归结果.其中,二元线性回归方程的R2以丛株数和花果数为自变量时最大,为0.975,以丛冠幅、平均丛高为自变量时最低,为0. 400;三元线性回归方程的R2介于0.817~0.977,以丛冠幅、丛株数、花果数为自变量时较高,其次为以平均丛高、丛株数、花果数和丛冠幅、丛平均高、丛株数为自变量的拟合方程,R2分别为0.975和0.955,而以丛冠幅、丛平均高和花果数为自变量的R2明显偏低;四元回归方程的拟合结果R2最大,为0.977,拟合结果较好.
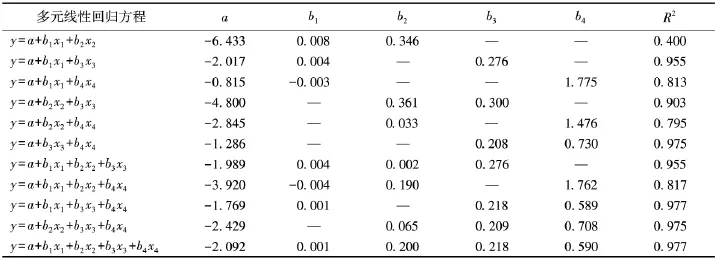
表4 荷青花总生物量与生长指标的多元线性回归方程Tab.4 M ultiple linear regression for total biomass and grow th indexes of Hylomecon japonica
4 结论与讨论
生物量作为植物群落能量基础和营养物质的重要组成部分,是生态系统生产力的重要体现,特别是在评价森林生态系统碳循环过程中具有不可替代的作用[12].受植物生长发育不同阶段个体光合作用能力差异及有限性的影响,其生长、繁殖和防御等各种功能间始终存在着资源竞争,因此植物必需权衡有限的光合资源分配问题[13-15],生活史对策正是这种不同功能间权衡资源分配的综合结果[16].由本次研究可知:荷青花群丛的生长指标及其生物量分配状况在不同群丛间存在较大差异,但在其营养生长停止期却有着相对稳定的根冠分配比例.荷青花个体数量的分布频数,除随平均丛高的增大呈“单峰型”变化外,随丛冠幅、丛株数和花果数的增大或增多均明显下降,表明荷青花地下部分所支持的地上营养面积、群丛个体数和花果数等存在一定的供给限制,且这种限制与地下生物量间存在着显著或极显著的正相关关系.
荷青花生物量随各生长指标的变化趋势:总生物量、茎叶生物量和地下生物量较为一致,均表现出整体随丛冠幅、平均丛高和花果数的增加呈先增高后下降的趋势,随丛株数的增多呈总体上升变化;花果生物量则表现为随丛冠幅、平均丛高的增加而增大,随丛株数和花果数的增加呈先增大后减小的变化.方差分析表明,除总生物量、茎叶生物量和地下生物量随平均丛高的变化差异不显著外,其余均存在显著或极显著的差异.荷青花生物量的分配比例整体以地下部分略高于地上,花果生物量不足总生物量的4%.随各生长指标的变化生物量分配比例变化趋势不一.经方差分析可知,根冠比仅随丛株数和花果数的变化差异显著,而地下生物量、茎叶生物量和花果生物量的分配比例随各生长指标的变化差异均不显著.由此表明荷青花个体生物量与各生长指标的关系密切,但生物量的分配比例却并不随生长指标的变化而表现出显著的差异,这在一定程度上印证了荷青花存在权衡有限光合资源分配的能力.
生物量模型的构建一直以来都是生物量研究的重点内容之一,现有研究结果以乔木植物居多[17-19],林下植被的模型构建还相对薄弱[20-24].本次基于荷青花生物量与各生长指标间的密切关系,分别构建了4种常用指数的一元线性回归方程,但仅以丛株数为自变量的直线方程和幂函数以及以花果数为自变量的幂函数R2达到了0.8以上.多元线性回归方程的R2明显高于一元线性回归结果;除以丛冠幅和平均丛高、丛冠幅和花果数、平均丛高和花果数为自变量的二元回归方程以及以丛冠幅、平均丛高和花果数为自变量的三元回归方程R2略低外,其余拟合结果均在0.9以上,拟合结果较好.然而在最优模型选择中,一方面要通过更多的变量得到最好的相关模型;另一方面又希望减少相关变量来避免更多的系统误差[24-25].根据以上原则和生物量模型拟合结果,虽以丛冠幅、丛株数、花果数或丛冠幅、平均丛高、丛株数、花果数为自变量的拟合最好,但因变量数量较多,亦可选用拟合结果次之的以丛株数和花果数为自变量的拟合方程.
[1]中国科学院中国植物志编辑委员会.中国植物志:第32卷[M].北京:科学出版社,1999:72.
[2]郑金萍,杨晶,郭忠玲,等.东北温带森林早春类短命植物种类及区系分布[J].北华大学学报(自然科学版),2014,15(4):516-521.
[3]严永清,余传隆.中药辞海:第二卷[M].北京:中国医药科技出版社,1996:1946.
[4]国家中医药管理局《中华本草》编委会,国家中医药管理局.中华本草3[M].上海:上海科学技术出版社,1998: 2269-2270.
[5]《浙江药用植物志》编写组.浙江药用植物志:上册[M].杭州:浙江科学技术出版社,1980:413-414.
[6]严仲铠,李万林.中国长白山药用植物彩色图志[M].北京:人民卫生出版社,1997.
[7]程虎印,程江雪,卫昊,等.太白七药拐枣七的研究进展[J].陕西中医学院学报,2011,34(4):94-95.
[8]赫丽丰,纪汉文,康洪学.黑龙江省早春野生花卉种质资源[J].国土与自然资源研究,1991(4):64-70.
[9]韩梅,杨利民,刘敏莉.吉林省野生早春草本花卉资源的筛选[J].中国野生植物资源,1997,16(4):22-23.
[10]胡晋燕,严鹰.耐荫地被及宿根花卉在公园中的应用研究[J].北京园林,2012,28(4):58-63.
[11]肖智,朱俊义,周繇,等.长白山草本观赏植物资源在园林景观中的应用[J].通化师范学院学报,2013,34(3):41-43.
[12]范春楠,庞圣江,郑金萍,等.长白山林区14种幼树生物量估测模型[J].北京林业大学学报,2013,35(2):1-9.
[13]Sutherland S,Vickery Jr R K.Trade-offs between sexual and asexual reproduction in the genus Mimulus[J].Oecologia,1988,76:330-335.
[14]Cheplick G P.Life history trade-offs in Amphibromus scabrivalvis(Poaceae):Allocation to clonal growth,storage,and cleistogamous reproduction[J].American Journal of Botany,1995,82(5):621-629.
[15]张大勇,姜新华,雷光春,等.理论生态学研究[M].北京:高等教育出版社,海德堡:施普林格出版社,2000.
[16]张景光,王新平,李新荣,等.荒漠植物生活史对策研究进展与展望[J].中国沙漠,2005,25(3):306-314.
[17]Wang C K.Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J].Forest Ecology and Management,2006,222:9-16.
[18]Chave J,Réjou-Méchain M,Búrquez A,et al.Improved allometric models to estimate the aboveground biomass of tropical trees[J]. Global Change Biology,2014,20(10):3177-3190.
[19]Goodman R C,Phillips O L,Baker TR.The importance of crown dimensions to improve tropical tree biomass estimates[J]. Ecological Applications,2014,24(4):680-698.
[20]Návar J,Méndez E,Nájera A,et al.Biomass equations for shrub species of Tamaulipan thorn scrub of Northeastern Mexico[J]. Journal of Arid Environments,2004,59(4):657-674.
[21]李晓娜,国庆喜,王兴昌,等.东北天然次生林下木树种生物量的相对生长[J].林业科学,2010,46(8):22-32.
[22]黄劲松,邸雪颖.帽儿山地区6种灌木地上生物量估算模型[J].东北林业大学学报,2011,39(5):54-57.
[23]Paton D,Nunez J,Bao D,et al.Forage biomass of 22 shrub species from Monfragüe Natural Park(SW Spain)assessed by loglog regression models[J].Journal of Arid Environments,2002,52(2):223-231.
[24]范春楠.吉林省森林植被碳估算及其分布特征[D].哈尔滨:东北林业大学,2014.
[25]唐守正,张会儒,胥辉.相容性生物量模型的建立及其估计方法研究[J].林业科学,2000,36(专刊1):19-27.
【责任编辑:郭伟】
On Spring Ephemeral Plants Biomass(Ⅳ)——Biomass and Its Model Building of Hylomecon japonica
Fan Chunnan1,Cheng Yan2,Zheng Jinping1,Liu Bin1,Zhao Zhe1
( 1.Forestry College of Beihua University,Jilin 132013,China; 2.Administration Bureau of Sanhu Nature Reserve of Songhua River in Jilin Province,Jilin 132013,China)
Based on the value of Hylomecon japonica in the medicine,ornamental and nutrient cycling of forest ecological system,the research obtained its biomass,distribution characteristics,and constructed the singleelement and multiple regression model for its total biomass with the growth indexes as the independent variable, by taking the whole plant harvest sampling method after sorting,drying,weighing and data statistics.The results showed that its growth indexes and biomass allocation varied remarkably from individuals,but its average height and root-shoot ratio changed smaller,when Hylomecon japonica association came to the withholding vegetative growth period.There was a certain supply constraint in crown width,the individual number and number of flowers and fruit of cluster supported by underground part,which had a significant positive correlation with the underground biomass.Although growth indexes of Hylomecon japonica were different with various degrees,but its root-shoot ratio difference was insignificant,confirming the Hylomecon japonica had the ability of resource allocation in weighing limited photosynthetic.It was relatively better to build the multiple linear regressions than singe-element regression; however,considering too many independent variables could cause some system errors, we selected a better fitting equation with individual number and number of flowers and fruit of cluster as independent variables.
spring ephemeral plants; Hylomecon japonica; biomass; distribution proportion; model building
Q948;S718.52
A
1009-4822(2016)03-0308-07
10.11713/j.issn.1009-4822.2016.03.006
2015-09-07
国家自然科学基金项目(31400387).
范春楠(1980-),男,博士,助理研究员,主要从事森林生态系统结构、功能研究,E-mail:fanchunnan@126.com;通信作者:郑金萍(1979-),女,博士,副教授,主要从事森林生态系统结构、功能及植被恢复研究,E-mail:zhengjinping80@126.com.
