基于生物位置细胞放电机理的空间位置表征方法
2016-08-30李伟龙吴德伟空军工程大学信息与导航学院西安710077西安通信学院西安710106
李伟龙 吴德伟 周 阳 杜 佳(空军工程大学信息与导航学院西安710077)(西安通信学院西安710106)
基于生物位置细胞放电机理的空间位置表征方法
李伟龙*①吴德伟①周阳①杜佳①②①
①(空军工程大学信息与导航学院西安710077)
②(西安通信学院西安710106)
为实现运行体智能自主定位,该文提出基于生物位置细胞放电机理的空间位置表征方法。首先建立运行体空间位置与运行体和不同路标之间距离的对应关系,以两种不同的空间覆盖方式构建位置细胞图,然后利用感知的距离信息激发位置细胞放电估计运行体位置,并分析了模型中各参数对空间位置表征和定位性能的影响。仿真结果表明,两种方法均能通过构建的位置细胞图实现空间位置表征和自主定位,但等间隔离散空间构建位置细胞图方法受距离间隔影响较大,而任意探索构建位置细胞图方法从生物自主感知空间位置的角度出发,通过选择合适的距离阈值和运动训练时间构建位置细胞图,能够更好地完成运行体空间位置表征并且定位精度也较高。
自主导航;仿生定位;空间表征;位置细胞
1 引言
人类是怎么知道自己身在何处,又是如何找到从一个地方到另一个地方路线,并且把这些信息存储下来,让自己重返故地的时候能立刻找到正确的路呢?2014年诺贝尔生理学奖的研究成果给出了答案。获奖者发现了大脑里精巧的“导航系统”,它像是一台精密的GPS,实时给出空间导航信息,指导人们向计划的目标前进[1,2]。其中,获奖者之一约翰⋅奥基夫通过实验发现,大脑海马体里存在一种神经细胞,每当大鼠身处屋子的某个特定地点的时候,这种细胞总是会被激活,而其他神经细胞则在大鼠身处其他地方的时候被激活,奥基夫称之为“位置细胞”(Place Cell,PC)[3],并认为如果每个地方都由对应的位置细胞来表示,大脑就生成了一张“认知地图”,这样就可以简单地把客观世界映射到了大脑里,实时给出人们导航信息。
上述研究是从生物学的角度出发的,而传统导航研究者往往关注的是如何从物理实现层面解释导航过程,那么如何利用生物学研究成果促进工学导航研究的发展呢?答案自然是建立合理的数学模型模拟生物学上的研究发现,并通过不断深入研究和改进应用于民用或军事方面。而目前关于生物位置细胞的研究资料[48]-,基本都是生命科学和脑科学相关专业的研究人员从实验的角度提出一些新的发现、新的机理,关于如何构建位置细胞图实现导航过程的资料很少。文献[9]通过大量的实验对海马区位置细胞的特性和功能进行了详细分析和说明;文献[10]重点分析了神经系统是如何告诉生物自己在空间环境的位置和方向的;文献[11]分析了各种导航细胞的特性,并通过建立模型解释了自我定位的神经机制。这些研究主要是通过生物实验上的重大发现阐述生物导航机理的,未见物理层面上的仿真实现。文献[12]从全景图像中提取局部视图作为路标,并利用磁罗盘量测路标的方位角,根据局部视图的相似程度定义位置细胞放电。该方法依赖于磁罗盘的测量精度同时对目标识别的要求较高,计算复杂度高且主要应用于室内小环境范围。文献[13]将自运动感知信息与视觉位置细胞信息相结合,构建多模位置细胞表示的仿生导航模型并用于机器人导航,仿真和实验结果验证了该模型具有一定的智能性。文献[14,15]利用头朝向细胞感知的方向和速度信息构建多层位置细胞,提出一种生物启发的目标导向导航模型并通过仿真实验证明了该方法的有效性。从这些研究中可以看出,脑科学的不断进步正在影响着导航技术的发展方向,利用脑科学领域的发现研究导航问题具有一定的可行性。但是,针对空间定位问题时,研究者很少将重点放在细胞放电情况与空间信息的对应关系上,在模型的实现中也缺少定位性能的分析。
于是,本文结合位置细胞的特点与功能,从传统导航研究者角度出发,提出基于生物位置细胞放电机理的空间位置表征方法,旨在说明该方法用于运行体自主定位的有效性和可实现性。文章接下来的具体安排是,先介绍位置细胞的特点及放电模型,然后对空间位置表征方法进行具体说明,并通过仿真实验对提出方法的定位性能进行详细分析,最后对全文进行总结。
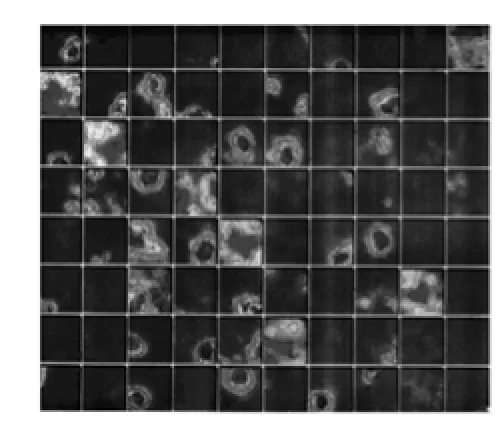
图1 位置细胞放电响应[16]
2 位置细胞特点及放电模型
文献[3]研究发现,自由活动的大鼠海马内某些神经元电活动具有明显的位置选择性,它与大鼠活动过程中所处的位置密切相关。当动物处在相对狭小的范围时,如果海马内某个细胞呈现最大频率放电,而在其他地方很少或没有放电,那么该细胞即称为位置细胞,其放电所对应的环境中的范围称为该细胞的“位置野”(Place Field)或“放电野”(Firing Field)[16]。这种特异性放电活动可以帮助动物判断自身在当前环境中所处的位置,单个位置细胞对应环境中的某一位置,特定的环境就可以用许多位置细胞的放电活动进行表征。图1给出了一次实验中大鼠CA 3和CA 1区的位置细胞放电响应图,在大鼠经历的区域都会有对应的位置细胞放电,放电野形状近似圆形,并且中间区域放电响应最大,依次向外递减,圆形外部深色区域放电强度很低,可认为没有放电。
根据实验中位置细胞放电响应图的形状,可以采用2维高斯函数进行模拟。设运行体所处的空间位置为p,则第i个海马位置细胞的放电率PCi(p)可以表示为

其中,is为第i个位置细胞的参考位置,σ为放电率调整参数。图2给出了250σ=时位置细胞的放电率分布情况,可以看出,模拟的位置细胞中间区域放电强度最大,并且呈圆形依次向外递减,正中心放电率为1,圆形外部深色区域放电率几乎为0。因此式(1)能够一定程度上模拟海马位置细胞特性。

图2 位置细胞放电模拟
3 空间位置表征方法
通过上一节的介绍可知,生物通过海马位置细胞描述所经历的空间位置,借鉴生物海马导航原理,提出基于位置细胞放电机理的空间位置表征方法。在构建位置细胞图时需要解决两个问题:(1)通过何种方式感知环境获取有用信息;(2)如何利用获取的有用信息形成位置细胞用于定位。针对问题(1),采用计算机视觉技术通过运行体搭载的视觉传感器感知环境,提取出识别率高、空间分布均匀的路标,为下一步构建位置细胞奠定基础,具体方法可参考作者之前的研究成果[17],这里不再展开;针对问题(2),利用提取出的路标,通过计算运行体与不同路标之间的距离构建位置细胞,编码运行体的空间位置。下面重点对问题(2)进行分析。
卫星导航通过空间位置坐标已知的卫星以及测量得到的伪距信息实现对用户的定位,其几何思想是三球交汇原理,即至少需要3颗(不考虑钟差)空间位置不同的卫星才能实现定位。受其启发,考虑通过建立运行体空间位置与运行体和不同路标之间距离的对应关系,构建位置细胞表征运行体在空间中的位置,即某一时刻运行体的位置可以由其与不同路标之间距离关系进行标定。具体步骤如下:
步骤1获取路标,建立空间对应关系。
首先通过视觉传感器和计算机系统在空间环境中选取N个稳定路标(其中3N≥,且至少有3个路标不在同一直线上),然后利用运行体与不同路标之间的距离关系,对运行体在空间环境中的不同位置进行编码。(例如运行体某一时刻在空间中的位置坐标为与N个路标之间的距离分别为建立空间对应关系
步骤2定义位置细胞放电。
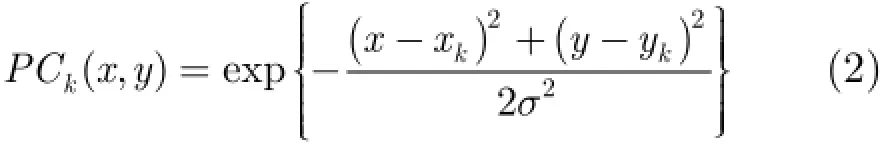
步骤3构建位置细胞图,表征空间位置。采用两种方式构建位置细胞图,一种采用等间隔离散空间构建位置细胞图方法;另一种采用空间任意探索构建位置细胞图方法。通过构建位置细胞图,使得空间中的任一位置都有对应的位置细胞放电,即于是,就能建立起位置细胞放电率与运行体和不同路标之间距离的对应关系,即当运行体再次到达空间中某一位置时,只要通过传感器获得与不同路标之间的距离关系,即可根据位置细胞的放电活跃度估计运行体的空间位置。
(1)等间隔离散空间构建位置细胞图:假定空间环境为一正方形区域大小为a a×,采用均匀分割的准则对空间区域离散化,分别沿横向和纵向以一定的间隔B将空间等分为m m×个小正方形区域,在每个小正方形中心点依次构建位置细胞,这样通过调整间隔B最终使位置细胞能够覆盖整个环境空间,即运行体在空间环境中任一位置都能找到相应的位置细胞放电,从而表征运行体的空间位置。
图3,图4给出了不同空间间隔B和不同放电野半径r位置细胞构建效果图(图中圆圈表示运行体对空间环境某个区域的位置响应,进而生成类似于生物脑的位置细胞,而并不是对运行体空间位置坐标的准确描述)。从图中可以看出,间隔B固定时,随着放电野半径r的增大,空间覆盖越来越多,不同位置细胞间重叠区域也越来越多,但构建的细胞数量不变,相应的位置野中心也不变;放电野半径r固定时,随着间隔B的增大,构建的位置细胞数量逐渐减少,空间覆盖范围和不同位置细胞之间的重叠区域也随之减少,进而位置描述的准确程度降低。因此,需要根据运行体定位精度要求选择合适的间隔B和放电野半径r构建位置细胞。
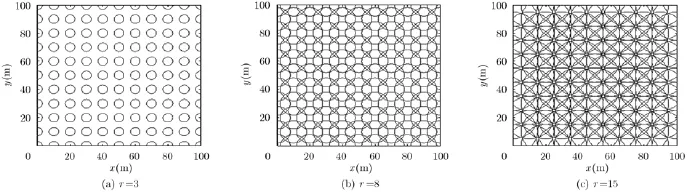
图3 不同放电野半径r下位置细胞构建效果图(B=10)
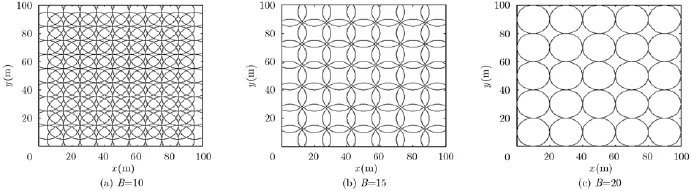
图4 不同间隔B下位置细胞构建效果图(r=10)
(2)任意探索构建位置细胞图:从生物自主感知环境的角度出发,根据生物在运动过程中对环境的认识和理解描述自己在空间中的位置。假定运行体在某一固定环境中任意运动,每一个时间间隔运行体都会根据自身当前位置与不同路标的距离关系构建一个位置细胞。这样经过一段时间构建的位置细胞会覆盖整个空间环境,进而运行体在空间中的任一位置都能用对应的位置细胞放电来表征。图5给出了一段时间内运行体在给定环境中通过任意探索构建的位置细胞图,其中黑线代表运行体的运动轨迹,圆圈表示构建的位置细胞。仿真参数设置如下:运动空间区域大小为100 m×100 m,定位周期为1 s,假设运行体在每个定位周期内速度保持不变,不同定位周期内速度在5~25 m/s区间内随机取值,方向任意选择,位置细胞放电野半径为5 m。
从图5中可以看出,构建的位置细胞数量与运行体在空间中的运动训练时间呈正比,运动训练时间越长,构建的位置细胞数越多,相应的空间覆盖率也越高。运动训练时间为500 s时,空间环境中有个别区域没有被位置细胞覆盖,而运动训练时间为1000 s时,构建的位置细胞能够覆盖整个空间环境,但同时可以观察到构建的不同位置细胞空间覆盖重叠区域所占比例很大,部分位置细胞的重叠区域甚至在90%以上,会带来不必要的存储资源占用,并且随着环境空间的进一步增大,这种资源占用消耗也会迅速增加,因此,应该采取一定措施减少空间覆盖重叠区域较大的位置细胞,使得能够以较少数量低重叠率的位置细胞达到覆盖空间环境的目的,从而减少占用的存储资源。
图6给出了相同仿真条件下筛选前和筛选后构建的位置细胞图,仿真参数设置如下:运动空间区域大小为100 m×100 m,定位周期为1 s,运行体在每个定位周期内速度保持不变,不同定位周期内速度在5~25m/s区间内随机取值,方向任意选择,运动训练时间1000 s,放电野半径5 m。筛选原则为设置一个距离阈值,当运行体当前位置与之前构建的位置细胞中心的距离大于阈值时,则在当前位置构建新的位置细胞;反之若小于阈值,则不在当前定位周期内构建位置细胞,继续下一时刻运动。从图6中可以看出,筛选前构建的位置细胞数量为1000个,筛选后位置细胞数量为315个,数量上大大减少,但对环境的覆盖程度几乎不变,因此该方法在减小计算复杂度的同时依然可以较好地表征空间环境。
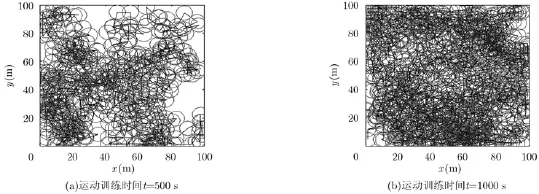
图5 不同运动训练时间下构建的位置细胞图
4 定位性能分析
针对上述两种位置细胞图构建方法,利用仿真实验分析其定位性能。让运行体在已构建好位置细胞图的空间环境中任意运动,记录运动轨迹,并利用运行体与不同路标之间的距离和构建的位置细胞图的对应关系实时估计运行体在空间中的位置。当运行体在空间中某一位置有位置细胞放电时,即落在某一位置细胞区域内时,将对应位置细胞的圆心作为当前估计位置。如果有多个位置细胞同时放电,选择放电率最大,即最靠近运行体的位置细胞。实验中仿真参数设置如下:运动空间区域大小为100 m×100 m,4个路标的位置分别为(10,23),(65,40),(35,70),(72,63),定位周期为1 s,运行体在每个定位周期内速度保持不变,不同定位周期内速度在5~25m/s区间内随机取值,方向任意选择。为分析定位误差,定义定位精度PP为运行体与不同路标之间估计距离与真实距离的偏差。
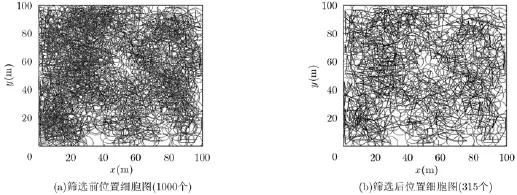
图6 筛选前后位置细胞图(距离阈值R=3m)
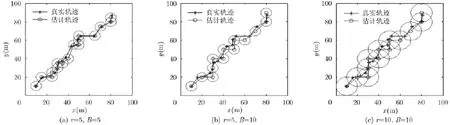
图7 不同间隔B和放电野半径r下的定位结果
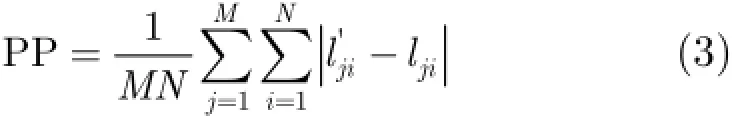
其中,M为位置数,N为路标的个数。
4.1方法1定位性能分析
通过改变仿真参数间隔B和放电野半径r,分析定位效果,图7给出了不同间隔B和放电野半径r情况下利用构建的位置细胞图进行位置估计的结果。从图7(a)和图7(b)对比可以看出,间隔B越小,构建的位置细胞数量就越多,定位精度也就越高;从图7(b)和图7(c)对比可以看出,放电野半径r的变化对定位精度并没有造成影响,这是因为以对应位置细胞圆心作为当前位置估计的方法只要间隔B相同,构建的位置细胞排列方式是相同的,所以最终位置估计的结果也是相同的。因此,这里主要分析间隔B对定位性能的影响。
图8给出了不同间隔B对定位误差的影响情况。对于每个间隔B都采用100组随机运动实验计算定位误差,最终取平均值作为输出结果。从图8中可以看出,随着间隔B的增大,定位平均误差逐渐增大,这是因为间隔B越大,相同空间区域构建的位置细胞数量就越少,对运行体实时位置估计的偏差就会增大。
4.2方法2定位性能分析
从前文分析可知,通过任意探索构建位置细胞图进行定位的方法主要与运动训练时间t和阈值R有关。图9给出了不同运动训练时间t和阈值R情况下利用构建的位置细胞图进行位置估计的结果。从图9(a)和图9(b)对比可以看出,运动训练时间相同的情况下,阈值R=3和R=5时都能对运行体的位置进行估计,阈值小的构建的位置细胞图比阈值大的构建的位置细胞图估计的位置更加靠近运行体的真值;从图9(b)和图9(c)对比可以看出,阈值R相同的情况下,运动训练时间越长,估计的位置越靠近运行体的真值。
图8 间隔B对定位误差的影响
同样利用式(3)给出的定位精度对该方法构建的位置细胞图用于运行体定位的效果进行分析。对于每个阈值和运动训练时间都进行100次仿真,取平均值作为最后定位误差的输出结果。表1和表2分别给出了定位精度随阈值R和运动训练时间t的变化规律。分析表中数据可以知道,运动训练时间t相同时,随着阈值R的增大,构建的位置细胞数量越少,并且减少的数量很大,定位误差也随之增大,同时阈值小于3m后,需要构建的位置细胞数量增长速度极快,这会占用过大的存储资源并且加大计算负担;阈值R相同时,随着运动训练时间t的增大,构建的位置细胞数量越多,定位精度也逐渐提高,但当运动训练时间大于1250 s后,构建的位置细胞数量仅有少量增长,并且定位精度提高的程度也不明显。图10进一步给出了定位误差受距离阈值和运动训练时间同时变化的影响。从图10中可以看出,定位误差受距离阈值影响程度较大,阈值过大导致位置细胞数量减少,构建的位置细胞图不能准确表征运行体的空间位置,阈值过小导致位置细胞数量大幅度增加,计算资源消耗迅速加大。因此,在处理实际问题中,应根据空间环境范围、位置细胞放电野大小以及运行体运动状态等条件大致设定距离阈值和运动训练时间等参数,进而综合考虑定位精度要求和计算复杂度,选择合适的距离阈值和运动训练时间构建位置细胞图,使系统能够在充分利用计算资源的情况下提高定位精度。
4.3对比分析
通过对两种位置细胞图构建方法进行对比分析可以发现,方法1构建思路比较简单,运算量小,但受位置细胞排列间隔影响较大,当排列间隔较大时定位误差较大,另外该方法不能自主生成位置细胞地图,不符合生物定位机理。方法2从生物自主感知环境的角度出发,通过训练使运行体不断加深对空间环境的认识和理解,逐渐构建和完善位置细胞地图,能较好地完成运行体空间位置表征,并且定位过程符合生物导航机理。从仿真分析中可以看出,该方法通过选择合适的距离阈值和运动训练时间,能够使运行体自主生成位置细胞图并完成自我定位,具有一定的有效性和可实现性。
图9 不同阈值R和运动训练时间t下的定位结果
表1 定位精度随距离阈值的变化情况(运动训练时间t=1000 s)

表2 定位精度随运动训练时间的变化情况(阈值R=3m)
图10 定位误差变化规律
5 结束语
生物位置细胞的发现及其在生物导航过程中表现出的位置特异性,使研究者一直在寻找具备生物位置细胞功能的模型,以将其应用于机器导航中。本文从导航定位需求角度提出两种通过构建位置细胞图表征运行体空间位置的方法,并进行了仿真分析。从仿真结果上看,两种方法均能实现自我定位,但基于任意探索构建位置细胞图的方法2更好地模拟了生物位置细胞功能,并且也更符合生物导航机理。同时,作者在整个研究过程中也受到一些启示。在进行与导航有关的研究中,生物学领域的研究者往往侧重于采用一定的技术手段通过进行大量的实验追求与导航相关的生物特性,注重证明发现过程的可信度。而工学领域的导航研究者往往更加关注如何从物理层面上解释导航相关问题,并努力通过各种技术方法实现导航过程。侧重点的不同导致很难将两者联系起来。现有的导航手段都是通过数学推导、方程解算等方式得到运行体具体的位置信息,而生物往往是通过多种感知方式认识和理解环境,并利用积累的经验知识获取自身在空间中的相对位置。因此,如何将生物对导航信息的处理机制与现有导航方式相结合是需要进一步深入研究的问题,作者认为生物导航机理的研究和进步必将以其独特的方式促进智能导航的研究发展。
参考考文
[1]BURGESSN.The 2014 nobel prize in physiology ormedicine: A spatialmodel for cognitive neuroscience[J].Neuron,2014,84(6):1120-1125.
[2]DERDIKMAN D and MOSER E I.A manifold of spatial m aps in the brain[J].Trends in Cogn itive Sciences,2010,14(12):561-569.
[3]O’KEEFE J and DOSLROVSKV J.The hippocampus as a spatialmap[J].Brain Research,1971,34(1):171-175.
[4]PFEIFFER B E and FOSTER D J.Hippocampal p lace-cell sequences depict future paths to remembered goals[J].Nature,2013,497:74-79.
[5]DAVACHI L and DUBROW S.How the hippocam pus preserves order:the role of prediction and context[J].Trends in Cognitive Sciences,2015,19(2):92-99.
[6]W ILLS T J and CACUCCI F.The development of the hippocam pal neural representation of space[J].Current Opin ion in Neurobiology,2014,24:111-119.
[7]MADL T,CHEN K,MONTALDI D,et al.Com putational cognitive models of spatial memory in navigation space:a review[J].Neural Networks,2015,65:18-43.
[8]HOWARD LR,JAVADIA H,YU Y,etal.Thehippocam pusand entorhinal cortex encode the path and Euclidean distances to goals during navigation[J].Current Biology,2014,24(12):1331-1340.
[9]RAYMOND P K and EDMUND T R.A com putational theory of hippocam pal function,and tests of the theory:new developments[J].Neuroscience and Biobehavioral Reviews,2015,48:92-147.
[10]HUGO J S and BARRY C.Neural systems supporting navigation[J].Current Opinion in Behavioral Sciences,2015,1:47-55.
[11]BARRY C and BURGESS N.Neural mechanism s of selflocation[J].Current Biology,2014,24:330-339.
[12]GIOVANNANGELI C and GAUSSIER P.Autonom ous vision-based navigation:goal-oriented action p lanning by transient states prediction,cognitive map building,and sensory-motor learning[C].2008 IEEE/RSJ International Conference on Intelligent Robots and System s,Nice,2008: 22-26.
[13]JAUFFRET A,CUPERLIER N,GAUSSIER P,et al. Multimodal integration of visual p lace cells and grid cells for navigation tasks of a real robot[C].12th International Conference on Sim ulation of Adap tive Behavior(SAB 2012),Odense,2012:136-145.
[14]ERDEM U M,M ILFORD M J,and HASSELMO M E.A hierarchical model of goal directed navigation selects trajectories in a visual environment[J].Neurobiology of Learning and Mem ory,2015,117:109-121.
[15]ERDEM U M and HASSELMO M E.A biologically inspired hierarch ical goal directed navigation m odel[J].Journal of Physiology-Paris,2014,108:28-37.
[16]O’KEEFE J and BURGESS N.Geom etrical determ inants of the place fieldsof hippocam pal neurons[J].Nature,1996,381: 425-428.
[17]李伟龙,吴德伟,杜佳,等.用于无人机自主定位的仿生视觉路标获取方法[J].仪器仪表学报,2015,36(6):1207-1214. LI W eilong,WU Dewei,DU Jia,et al.The landm ark acquisition m ethod based on bionic vision for UAV autonomous positioning[J].Chinese Journal of Scientific Instrument,2015,36(6):1207-1214.
李伟龙:男,1989年生,博士生,研究方向为智能自主导航.
吴德伟:男,1963年生,教授,博士生导师,研究方向为导航定位理论、技术及应用、智能自主导航、量子导航.
A Method of Spatial Place Representation Based on Biological Place Cells Firing
LIWeilong①WU Dewei①ZHOU Yang①DU Jia①②①
①(Information and Navigation College,Air Force Engineering University,Xi’an 710077,China)
②(Xi’an Communications Institute,Xi’an 710106,China)
In order to realize intelligent and au tonom ous navigation of vehicles,a m ethod of spatial p lace representation based on biological p lace cells firing is proposed.A relationship is built that the spatial location of vehicle is corresponding with the distances between the vehicle and different landmarks,and amap of place cells is constructed in two different ways of space coverage.Then using the real-time distances sensed inspires p lace cells firing so as to estimate the location of the vehicle.An analysis is carried out about different parameters in the model in fluence on spatial location exp ression and the performance of positioning.The simulation results show that both two ways can realize location exp ression and au tonom ous positioning th rough using them ap of p lace cells. Theway that the space is separated equally is influenced by the distance interval greatly,and the way of building themap of place cells through exploring random ly,starting off with the perspective of sensing spatial location autonomously,can finish spatial location expression and autonomous positioning better through choosing appropriate interval and trained time.
Autonomous navigation;Biological positioning;Spatial representation;Place cell
The National Natural Science Foundation of China(61273048)
TP391
A
1009-5896(2016)08-2040-07
10.11999/JEIT 151331
2015-11-26;改回日期:2016-04-18;网络出版:2016-06-16
李伟龙weilongli2008@126.com
国家自然科学基金(61273048)
