不同植物种类及棉花品种对扶桑绵粉蚧生物学影响研究进展
2016-08-23鹿东东商晗武
鹿东东,商晗武
(中国计量学院生命科学学院,杭州 310018)

不同植物种类及棉花品种对扶桑绵粉蚧生物学影响研究进展
鹿东东,商晗武*
(中国计量学院生命科学学院,杭州 310018)
扶桑绵粉蚧Phenacoccussolenopsis是一种多食性入侵害虫,其寄主范围广、适应性强,已入侵多个国家和地区,对农业生产造成了严重危害。本文通过分析不同寄主植物上扶桑绵粉蚧研究,找出相对适宜与不适宜寄主植物,以便指导作物种植,并为培育抗扶桑绵粉蚧作物提供一定参考。本文通过收集分析相关文献,运用数理统计方法,综述和分析了不同寄主植物上扶桑绵粉蚧生长发育、生殖、抗逆等方面研究进展及其差异。发现马铃薯、豚草是极有利于扶桑绵粉蚧繁衍的植物,其次是扶桑与棉花。对10个品种棉花的分析发现,中棉所59对扶桑绵粉蚧具有较强的抗性,而抗虫棉RCH-134不具有抗性。对扶桑绵粉蚧生物学尤其是生长发育、繁殖能力等研究已经比较多,今后与该虫相关的抗虫育种、生物防治、分子生物学等研究可望成为重点。
扶桑绵粉蚧;寄主植物;棉花
扶桑绵粉蚧PhenacoccussolenopsisTinsley,最早在美国新墨西哥州被发现并命名(Fuchsetal., 1991),其后一直在美国的中部、加勒比群岛和厄尔多瓜为害蔬菜和花卉作物(Williams and Willink, 1992)。随后,该虫在智利和巴西的番茄和几种杂草上被发现(Culik and Gullan, 2005)。我国于2008年在广州首次发现了该虫(马骏等, 2009; 武三安和张润志, 2009),2009年调查结果显示,该虫已在我国的8个省和自治区有零散分布(张润志等, 2010),其已被列为我国第436种检疫性有害生物(刘刚, 2009)。扶桑绵粉蚧是一种入侵性害虫,在过去的一个世纪里,它入侵了超过23个国家(Ahmedetal., 2015)。该虫能严重危害棉花,在巴基斯坦和印度造成棉花大面积减产;为防治该虫,仅在旁遮普地区2个月内农药的使用费用就达1.2亿美元(Hodgsonetal., 2008;Nagrareetal., 2009),由此可见其对农业生产的为害性。为有效地防治该虫,已对其进行了多方面的研究(Luetal., 2011;Guanetal.,2012; Muhammadetal., 2015),对这些研究结果进行的综述也已有报道(胡熙熹等, 2012),但尚未见应用数理统计方法对这些资料进行综述分析。本论文应用数理分析方法对寄主植物与扶桑绵粉蚧关系研究结果进行综合剖析,以期望找出相对适宜与不适宜寄主植物,以便以此为依据对作物种植方式进行指导,有效地防止该虫蔓延暴发,并为培育抗虫作物提供参考。
1 材料与方法
1.1研究材料
供综析的文献是至今国内外已公开发表的45篇研究报告,文献来源于CNKI—中国学术期刊(网络版)(中国期刊网)、万方数据资源、维普中文科技期刊数据库、Web of Science(SCI、SSCI、CPCI)数据库、Elsevier ScienceDirect 数据库、IEL(IEEE/IET Electronic Library)数据库、Springer Link电子期刊全文数据库、 EI数据库等中外文文献数据库。
1.2分析方法
通过查阅相关文献,运用多重比较差异分析方法(黄雨舜,2005;钱俊和陈平雁,2008),对扶桑绵粉蚧的有关研究进行综合分析。
2 结果及分析
扶桑绵粉蚧是一种多食性害虫,它通过吸食植物汁液,致使植物发育迟缓,甚至失水干枯;分泌的蜜露能吸引蚂蚁,诱发煤污病,降低光合作用,严重时致使植物大片死亡(Arifetal., 2009);最近的研究还发现,该虫还能抑制植物的防御反应,使其转化为有利自己的方向,方便其自身取食和生长发育(Zhangetal., 2015)。其寄主范围广,繁殖能力强,并兼有有性生殖和孤雌生殖两种繁殖方式,极易转移扩散,使得多种作物严重减产。所以该虫入侵地的科学家均对其十分重视,并对其进行各方面的研究。
相对其他方面来说,对扶桑绵粉蚧生物学方面的研究较多。在形态学方面已有较多研究(Guanetal., 2012),其发育、繁殖特性也已基本清楚(关鑫和陆永跃, 2012; 王超等, 2014; 王超等, 2016)。扶桑绵粉蚧为卵胎生,卵产在卵囊内,产卵量大,单雌产卵数为128-812粒,平均为344粒(Vennilaetal., 2010; 朱艺勇等, 2011),其生活史为:(1) 雌虫分3个虫态:卵→若虫(1龄若虫—2龄若虫—3龄若虫)→雌成虫;(2) 雄虫分4个虫态:卵→若虫(1龄若虫—2龄若虫)→预蛹→蛹→雄成虫(朱艺勇等, 2011)。在寒冷地区扶桑绵粉蚧以卵或其他虫态在植物或土壤中越冬;在热带地区扶桑绵粉蚧可终年繁殖,种群增长迅速,世代重叠严重(Hodgsonetal., 2008)。在生育特性方面研究较多的是扶桑绵粉蚧生长发育与温度的关系,其生长发育的最低、最适、最高温度基本清楚,一般在18℃及其以下、35℃及其以上均不利其生长,而26℃-30℃是其最适生长温度(Amarasekareetal., 2008;Hodgsonetal., 2008; Fandetal., 2014),过冷却点最低为-24.02℃(关鑫等, 2009),故其能够较好的越冬。在防治方面化学防治研究较多。农业防治基本上没有研究,还未见具有抗扶桑绵粉蚧的棉花品种育成。生物防治应是防治扶桑绵粉蚧的良好方法(Ahmadetal., 2011),已开展了不少基础工作,已报道捕食天敌、寄生性天敌14种、12种(Ashfaqetal., 2010; Tanwaretal., 2011; Arifetal., 2012),也研究了微生物和植物源杀虫剂,但还属于初步的研究(黄芳等, 2015)。已经开展了扶桑绵粉蚧在不良生态环境中抗逆性的研究如饥饿胁迫(郑婷等, 2011)、高温胁迫(关鑫等, 2011;Sreedevietal., 2013)、低温胁迫(Sreedevietal., 2013)与干旱胁迫(Kumaretal., 2013)等。对该虫在棉花、番茄、茄子、扶桑、秋葵、豚草等多种寄主植物上的生物学已进行了研究(黄芳等, 2011; 关鑫等, 2012),对不同品种棉花上生长发育等也进行了研究(赵新宁, 2012;赵新宁等, 2012)。同时,吕茂翠(2013)研究了锦葵科、旋花科、茄科、胡麻科、葫芦科的5种寄主植物对扶桑绵粉蚧的适应性。
2.1不同植物上扶桑绵粉蚧生物学差异
昆虫的生长发育和种群动态与寄主植物息息相关。寄主不同,昆虫的生长、繁殖、寿命等也会有差异(Nagrareetal., 2009)。研究表明寄主植物是影响扶桑绵粉蚧生长发育和繁殖力的最重要因素。扶桑绵粉蚧的寄主范围极广,包括作物、蔬菜、观赏植物和杂草等,共57科149属207种,其中对禾本科、锦葵科、茄科、葫芦科、大戟科、胡麻科、苑科等的为害较严重(张润志等, 2010)。已有研究表明,对于扶桑绵粉蚧在不同植物上为害时,其个体的发育特征,随着寄主植物不同,该虫生长发育参数也有所不同。Akintola等(2008)研究发现扶桑绵粉蚧在扶桑上的若虫期1龄<2龄<3龄。黄芳等(2011)研究了扶桑绵粉蚧在棉花、番茄和茄子上的个体发育情况,其成虫期>若虫期,若虫期1龄>3龄>2龄;且发现扶桑绵粉蚧的存活率和产卵量高低依次为棉花>茄子>番茄;在烟草、扶桑、棉花、马铃薯4种植物上扶桑绵粉蚧的生物学差异(关鑫等, 2012),其若虫期与黄芳等(2011)研究所获规律相同。王莹莹等(2012)研究了南瓜上扶桑绵粉蚧在不同温度条件下的发育历期,发现该虫在南瓜上的若虫期显著高于其他植物,且在各温度条件下若虫期依次为3龄>1龄>2龄。Mamoon-ur-Rashid 等(2012)研究了在棉花、扶桑和秋葵上扶桑绵粉蚧的发育历期,若虫期依次为棉花<扶桑<秋葵。
2.1.1若虫(蛹)期
由于扶桑绵粉蚧的雌虫的若虫有1、2、3龄之分,而雄虫有1、2龄,相应的雌虫的3龄属蛹期。为了利于分析,本文将该虫雌虫1-3龄的历期之和称若虫期,而雄虫的1-2龄期加蛹期简称为若蛹期。
表1、2是根据有关文献资料统计分析扶桑绵粉蚧雌虫若虫期与雄虫若蛹期的结果。从表1、2可见,10种植物,无论是雌虫或是雄虫都是南瓜的若虫期、若蛹期最长,并均显著长于其他9种植物。在这10种植物中,若虫(蛹)期相对较长的是南瓜、番茄与马铃薯;相对较短的为棉花、扶桑与豚草。从表1、2可知,经至今所获研究资料的统计,在南瓜上扶桑绵粉蚧若虫期与若蛹期的均值分别为26.71 d、23.20 d;在番茄上为17.63 d、19.87 d;在马铃薯上为17.64 d、18.24 d;在棉花上为16.02 d、16.50 d;在扶桑上为15.68 d、16.21 d;在豚草上为14.81 d、15.30 d。番茄、马铃薯的若蛹期与棉花、扶桑、豚草的差异同样都达显著水平,但若虫期在它们之间的差异不显著。而从不同龄期供析的10种植物的平均值来看,不论雌、雄虫都是1龄若虫的龄期最长。雌虫1龄与2龄差异显著;1龄与3龄,2龄与3龄差异不显著;雄虫1龄与2龄、3龄差异显著。

表1 在不同植物上的扶桑绵粉蚧雌虫若虫期分析
续上表

寄主植物Host-plants1龄(d)1stinstar2龄(d)2ndinstar3龄(d)3rdinstar若虫期(d)Nymphperiod棉花Gossypiumspp.(同上)6.39±0.094.38±0.145.25±0.0916.02±0.11b扶桑Hibiscusrosa-sinensis(同上)6.37±0.114.20±0.145.11±0.1115.68±0.10b豚草Ambrosiaartemisiifolia(同上)5.74±0.104.16±0.124.91±0.1214.81±0.11b平均值Mean6.72±0.50a5.13±0.54a5.80±0.42ab-
注:表中数据,同列或同行(仅平均值)字母相同时,表示处理间在P<0.05水平差异性不显著。下表同。Note: Date in the table in the same column or peer(only mean) followed by same letters are not significantly different atP< 0. 05.The same below.
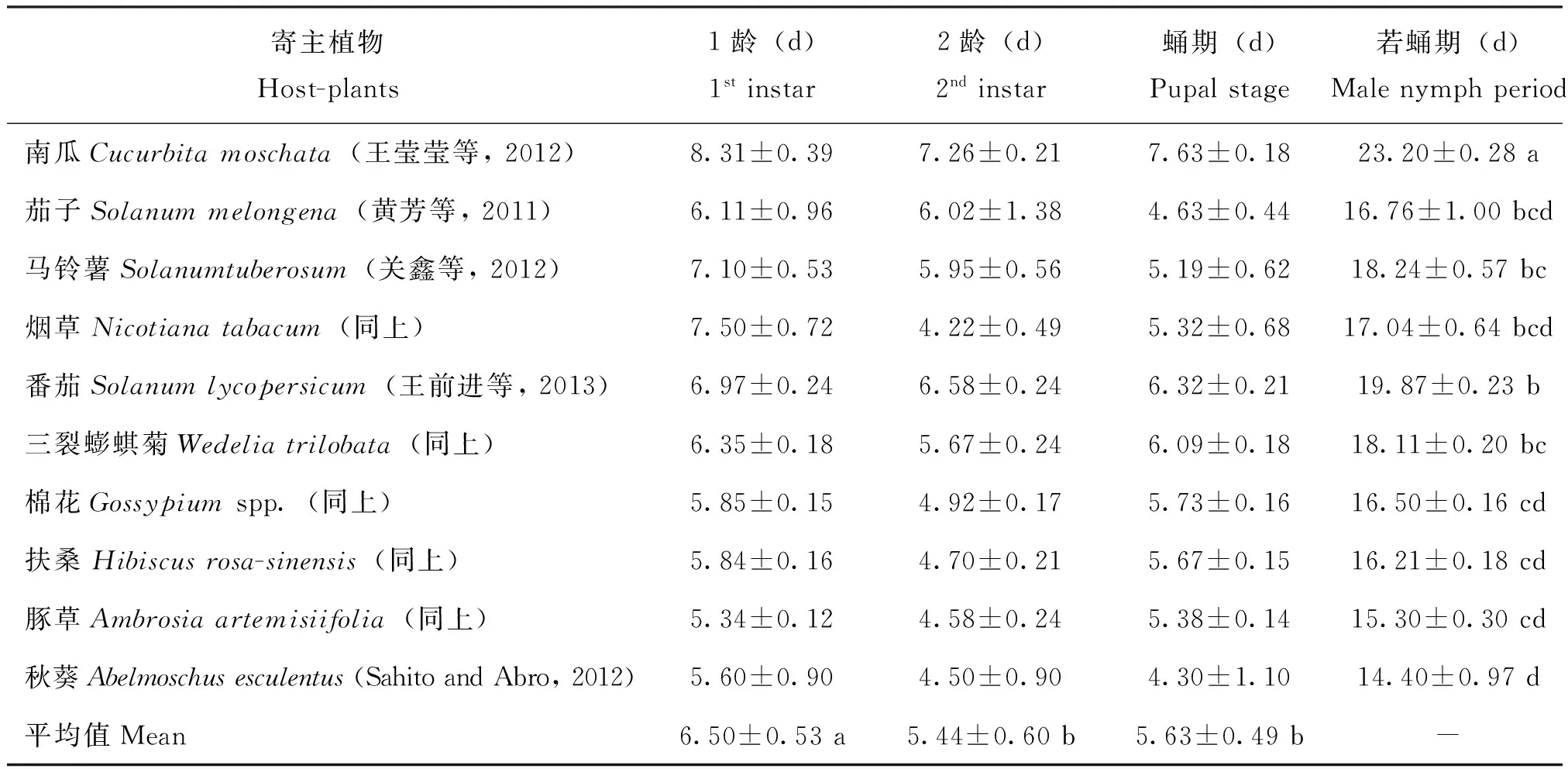
表2 在不同植物上的扶桑绵粉蚧雄虫若蛹期分析
2.1.2成虫期、若虫-成虫期
经8种植物的综析结果,见表3。雌、雄成虫期均是番茄最长;最短的,雌虫为南瓜,雄虫为豚草,南瓜倒数第二。经差异显著性分析,雌虫的成虫期在番茄、蟛蜞菊、棉花、扶桑之间以及在豚草、茄子之间差异不显著,在南瓜与秋葵之间差异显著;南瓜、秋葵与其它植物,豚草、茄子与其它植物之间差异显著;雄虫成虫期是在番茄、蟛蜞菊、茄子、秋葵之间及在棉花、扶桑、南瓜之间差异不显著;而豚草与其它植物,棉花、扶桑、南瓜与其它植物差异显著。总之,对成虫期的综析发现,雌、雄虫均是番茄、蟛蜞菊的成虫期较长,而南瓜、豚草相对较短,且番茄、蟛蜞菊的成虫期均显著长于南瓜与豚草。而若虫-成虫期(寿命),综析结果也是番茄与蟛蜞菊的历期较长,但相对较短仅为豚草;南瓜,在雌虫为最短,在雄虫为最长。这4种植物之间的若虫-成虫期,除雌虫的南瓜与豚草间差异不显著,其余差异均显著。
2.1.3产卵前期、产卵期与产卵量
表4是经8种植物分析了产卵前期、产卵期及产卵量在8种植物上的表现。从表4可知,雌虫的产卵前期及产卵期均是马铃薯最长,扶桑与豚草相对较短,且马铃薯与其它7种植物间的差异均达显著水平。而茄子的表现较特殊:其产卵前期较长,并与蟛蜞菊、扶桑、棉花及豚草的差异均显著,但其产卵期却是最短的。产卵量的分析,缺少茄子,共有植物7种,其结果,马铃薯的产卵量最高,平均达552.50粒,并除豚草均与其它五种植物的差异达显著标准,而番茄、烟草的产卵量相对较低,平均为342.63粒与417.20粒,且两者之间的差异也达显著水平。比较番茄、烟草两种植物,扶桑绵粉蚧的产卵前期、产卵期均相对较长;豚草的产卵前期及产卵期的表现与它的相比恰恰相反,产卵前期最短,为10.85 d,产卵期倒数第二位为15.84 d。豚草的产卵量为第二位,达511.12粒,与马铃薯差异不显著。综析结果,在供析的植物中仅马铃薯的产卵前期、产卵期及其产卵量,3者均居第一位。

表3 扶桑绵粉蚧在不同植物上的雌、雄虫成虫和若虫-成虫期的分析
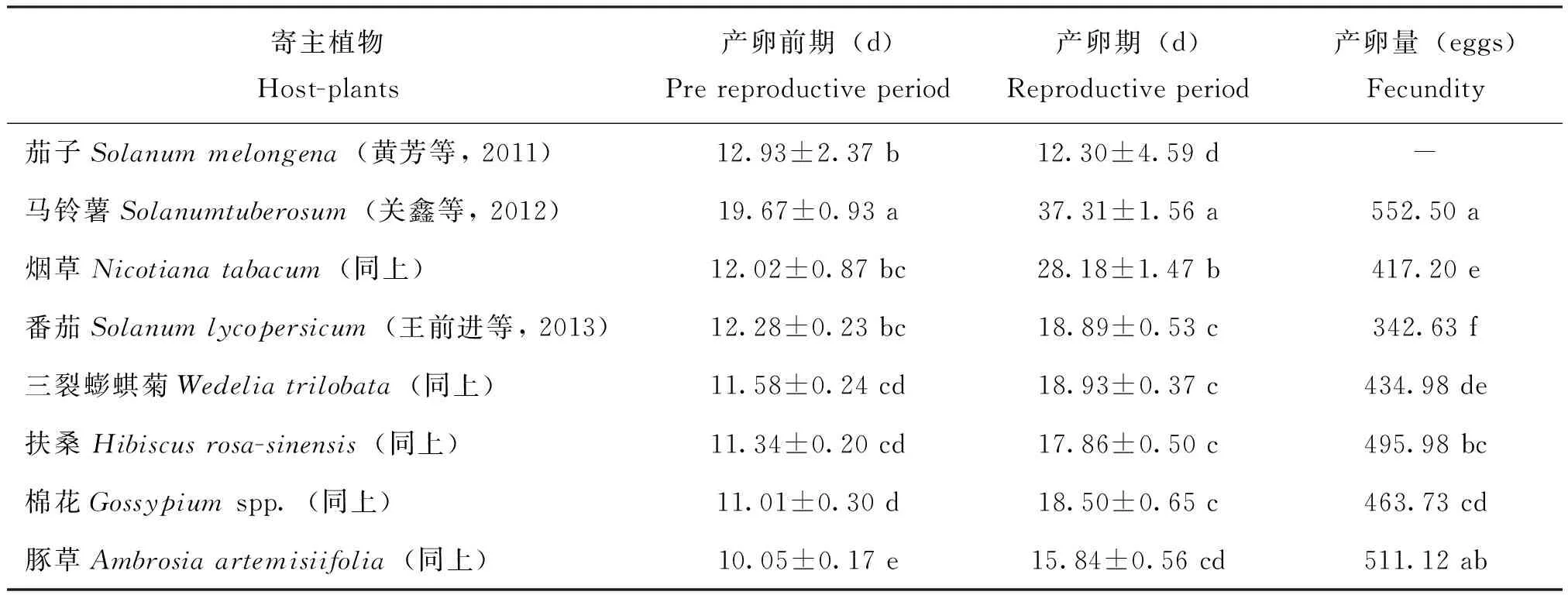
表4 不同植物上扶桑绵粉蚧产卵情况的分析
2.2对不同棉花品种的综析结果
至今,国内外对扶桑绵粉蚧为害棉花时生物学特性的表现,已有不少报道。Vennila等(2010)在实验室条件(8月和10月平均气温和湿度条件)下研究了扶桑绵粉蚧在转基因棉花上的生育历期,并分析了其繁殖特点,以便采取有效措施来防治扶桑绵粉蚧;Prasad等(2012)用不同温度对扶桑绵粉蚧在转Bt基因棉NCS-207上的生育历期作了统计分析;Kumar等(2013)对转Bt基因棉RCH-134上的扶桑绵粉蚧就不同温度和不同湿度的生育历期作了分析;王飞飞等(2014)统计了不同温度下扶桑绵粉蚧在浙凤棉1号的生育历期情况;赵新宁等(2012)比较了扶桑绵粉蚧在转基因棉和非转基因棉上的生长历期,发现其1龄若虫期都显著长于2、3龄若虫期,但若虫期在中棉所 59上最短,为15.6 d,在中棉所48上最长,为17.1 d;产卵期在中棉所41上最短,为13.6 d,在湘杂棉上最长,为17.9 d;产卵期在中棉所41上最长,为15.7 d,在湘杂棉上最短,为12.4 d;其寿命在中棉所41上最长,为48.1d,在中棉所59上最短,为43.6 d。并通过测定不同棉花品种叶片中可溶性糖、可溶性蛋白与棉酚含量,将不同品种生化物质含量与相应危害扶桑绵粉蚧的生物学参数作相关分析,结果发现,棉叶较高可溶性糖含量会降低扶桑绵粉蚧死亡率,延长产卵前期,有利于扶桑绵粉蚧生长发育与繁殖;而较高的棉酚含量会影响扶桑绵粉蚧正常生长发育,抑制其繁殖,提高死亡率。
2.2.1若虫期
表5是对扶桑绵粉蚧在10个棉花品种上的龄期及若虫期的多重比较分析结果。10个棉花品种扶桑绵粉蚧的不同龄期的平均值1龄最长为6.18 d,2龄最短,为5.05 d。1龄与2龄、3龄差异显著,2龄与3龄差异不显著。10种品种的平均若虫期,NCS-207最长为19.9 d,RCH-134最短为13.20 d,差异达显著水平。NCS-207、浙凤棉1号、中棉所48这3个品种的若虫期相对较长,且三者间差异不显著,中棉所59、RCH-134相对较短,二者间差异也不显著,但RCH-134与浙凤棉1号、中棉所48差异达显著水平。此外,其它5个品种的若虫期差异均未达显著标准。

表5 扶桑绵粉蚧在10个棉花品种上的若虫期分析
2.2.2成虫期、若虫-成虫期
经9个品种的综析结果,见表6。成虫期,中棉所59最短,为27.88 d,而RCH-134最长,为42.40 d,且均显著长于其它8个品种。居第二位是浙凤棉1号为34.67 d,其与中棉所44、47、59及湘杂棉2号差异达显著水平,而后4个品种的成虫期差异不显著。若虫-成虫期同样是RCH-134与浙凤棉1号较长,分别为55.60 d与52.11 d;中棉所59最短,为43.55 d,且供析的8个品种的差异显著性也与成虫期较相类同,所不同的是,RCH-134与浙凤棉1号还与中棉所48、湘杂棉9号的差异达显著水平。
2.2.3产卵前期、产卵期与产卵量
表7是9个品种的产卵前期与产卵期的分析结果。结果,产卵前期浙凤棉1号居中为14.56 d,湘杂棉2号最长为17.91 d,与中棉所44、41、59差异不显著,但这4个品种均与中棉所48、47,湘杂棉9号及RCH-134这后4个品种差异达显著水平。中棉所48与47差异不显著,但与湘杂棉9号、RCH-134差异均显著。产卵期RCH-134、湘杂棉9号与湘杂棉2号3个品种完全呈现了与产卵前期相反的特性,RCH-134 产卵期居第一为30.2 d,并显著长于其它8个品种,湘杂棉9号居第二为18.5 d,与中棉所44、59及湘杂棉2号差异显著,湘杂棉2号最短为12.37 d。产卵量的供析资料缺浙凤棉1号,据8个材料的分析结果,产卵量居首的为湘杂棉9号,卵量高达463.73粒,第二位的是RCH-134,卵量达334.40粒,最低的为中棉所59,卵量仅105.01粒,3个品种间的产卵量差异均达显著水平。

表6 扶桑绵粉蚧在10个棉花品种上的成虫期及若虫-成虫期分析

表7 扶桑绵粉蚧在10个棉花品种上的产卵情况分析
3 结论与讨论
经综合分析发现,马铃薯与豚草是极有利于扶桑绵粉蚧扩散繁衍的植物,其次是扶桑与棉花。而经10个品种的棉花综析发现,中棉所59具有较好的抗扶桑绵粉蚧的作用,而抗虫棉RCH-134对扶桑绵粉蚧不具有抗性。可具体归纳叙述成以下3点。
3.1不同龄期的表现
无论是雌虫还是雄虫,或是在不同的植物上及其在不同的棉花品种上,扶桑绵粉蚧综析结果的龄期平均值均是1龄的历期最长,2龄的最短,且除RCH-134以外,1龄历期都是显著长于2龄、3龄或蛹历期,而2龄与3龄或蛹历期的差异不显著。分品种看龄期,10个供析棉花材料有8个品种的1龄历期最长,仅RCH-134的1龄历期特短为3.9 d,其寿命最长为42.4 d,产卵量居第二位。除RCH-134外,其它9个供析材料,浙凤棉1号的1龄历期特长为7.33 d,余下8个品种,即在大多数棉花品种上1龄历期约在6-6.5 d之间。
3.2在不同植物上的表现
从综析结果可知,雄虫在多种植物上表现的若蛹期、成虫期及其寿命较其雌虫有规律性,尤其是若蛹期与寿命之间,不同植物间的历期具有显著正相关特性(r=0.969,P=0.000),而雌虫就没有此特性,这可能与雌虫需产卵有关。经归纳综析结果可以发现:雌虫若虫期居于极端位的南瓜与豚草,其寿命均居末位。如果“产卵量”是反映世代转换能力的一种指标,即物种能繁衍生存的一种能力,由于居一二位的南瓜、秋葵无产卵量资料数据,以居第四位的番茄观之,其成虫期居一,寿命居一,产卵前期居三,产卵期居四,但产卵量却居末位。这是否是番茄对扶桑绵粉蚧来讲,是不利于该虫繁衍,而利于其个体生存?南瓜与秋葵是否也是如此,至今还尚缺数据可定。豚草各生育阶段的历期均较短,若虫期居末位,成虫期倒数第四,寿命倒数第二,产卵前期居末位,产卵期倒数第二,然而其产卵量却居第二,与马铃薯差异不显著,结果豚草似乎是一种有利其世代繁衍的植物。马铃薯若虫期居第三位,与番茄相比仅长0.01 d,因此,这两种植物的若虫期应该是相同的,但产卵量马铃薯居首,据其产卵前期、产卵期也居首,故可推测其寿命也不短,即马铃薯是一种使扶桑绵粉蚧长寿且产卵量高的植物。故从反映世代转换的能力来看,马铃薯与豚草是利于扶桑绵粉蚧扩散繁衍的植物,其次是扶桑与棉花。
3.3在不同棉花品种上的表现
经综析结果,从表6可知棉花成虫期与寿命的长短,在不同品种间具有良好的相关性,经相关性分析呈极显著正相关(r=0.938,P=0.002)。而供析的10种棉花品种,若虫历期居首的是NCS-207,但其无相应其它生育阶段的数据资料,而若虫历期居第2位的浙凤棉1号无相应产卵量数据。然尽管如此,其它8个品种的若虫历期与产卵量的关系明显可分为两种情况:(1)中棉所的48、44、41、47的若虫期与产卵量具有互为相反相成的特点;(2)另4个若虫期居后四位的品种,其产卵量湘杂棉2号与中棉所59居末尾;而湘杂棉9号与RCH-134却依次居第一、二位,并且产卵量显著高于湘杂棉2号与中棉所59。换言之,不同品种棉花经综合分析结果,8个品种有6个品种呈现了若虫历期长短与产卵量大小呈负相关规律;两个品种呈现了正相关特性。尤其是中棉所59,其若虫期、产卵期均居末二位,成虫期、寿命、产卵量均居末位,是一个明显不利扶桑绵粉蚧生育与繁殖的品种,而RCH-134与湘杂棉9号若虫期分别为末位与末三位,但其产卵量分别居第二与第三位,且产卵量显著高于中棉所59,由此可见,这两个是极利于扶桑绵粉蚧生存扩散的品种。故经综合分析表明,扶桑绵粉蚧的若虫期短,可能有两种截然相反的原因导致:(1)寄主植物有利于粉蚧取食消化而促进个体发育加快的缘故;(2)寄主植物被粉蚧取食而难以消化,使个体难以正常发育的缘故。后者明显表现了品种对粉蚧具有一定抗性作用,这已被赵新宁(2012)研究证明。当扶桑绵粉蚧取食中棉所59以后,粉蚧体内解毒酶明显增加,以保其能安全生存繁衍,与湘杂棉9号相比,中棉所59产卵量相对要少4.42倍;这说明湘杂棉9号对粉蚧无抗性作用,而虽然供析材料RCH-134为转基因抗虫棉,但它也并不具有明显抗扶桑绵粉蚧作用,其产卵量仅次于湘杂棉9号,居第二位,有334.40粒。
有关扶桑绵粉蚧在生物化学及分子遗传学方面的研究,目前国内外还较少,尤其是生物化学方面的研究更少。赵新宁(2012)通过测定不同棉花品种叶片中的生化物质含量与其相应危害扶桑绵粉蚧的生物学参数作了相关分析。王伟兰(2014)对扶桑绵粉蚧对不同寄主的取食偏好性作了研究,测定了不同寄主植物上可溶性糖、可溶性蛋白及单宁含量,发现扶桑绵粉蚧的取食选择性与这些物质无显著相关性。Singh等(2012)采用PAPD分子标记技术对5种寄主植物上20个扶桑绵粉蚧种群进行遗传多样性分析,结果可将这20个种群分为2个分支,这说明扶桑绵粉蚧的生长发育可能与不同植物的抗性和寄主专化性有关。田虎等(2013)采用mtDNA COI 基因序列的特异性引物(SS-COI)进行PCR扩增,鉴别扶桑绵粉蚧;何衍彪等(2011)通过对12种粉蚧的 18S rDNA,28S rDNA 和 COI 序列的多重比对和聚类分析,探讨了其用于粉蚧系统进化分析及种类鉴定的可行性,结果表明 COI 基因的序列变异率相对较高,完全可作为近缘属种粉蚧类昆虫识别的依据。罗梅等(2012)首次克隆了扶桑绵粉蚧的组织蛋白酶B基因(PsCB)的cDNA 序列,随后对扶桑绵粉蚧的环指蛋白基因(PsRing3)(罗梅等, 2014)、钙调蛋白基因(PsCam)(罗梅等, 2013)和谷胱甘肽-S-转移酶基因(PsGSTzl)(罗梅等, 2012)序列也进行了克隆分析,为进一步研究这些基因对扶桑绵粉蚧的生长发育及其代谢的影响提供了材料。罗梅等(2014)也利用高通量测序的方法对扶桑绵粉蚧转录组的数据进行搜索,共设计出 1228对引物,为扶桑绵粉蚧遗传多样性分析、进化分析及入侵生物学等奠定基础。Thomas等(2014)使用线粒体细胞色素氧化酶1(mtCO-1)基因对扶桑绵粉蚧进行遗传多样性分析,结果显示,来自于亚洲不同寄主植物的扶桑绵粉蚧mtCO-1基因序列无明显变化,但用NCBI基因库中可用序列构建系统发育树,显示亚洲和美国之间具有明显的种群隔,进一步了解了扶桑绵粉蚧与寄主相关的形态变异和种群间的遗传多样性问题。
因此,今后有关扶桑绵粉蚧的抗虫育种、生物防治及生物化学和分子遗传学方面的研究可望成为重点,所以以后的研究应(1)对扶桑绵粉蚧的基因组进行组装与注释,基因组测序,丰富数据库,并对扶桑绵粉蚧多种基因功能进行研究,从分子水平初步解释其生物学特性;(2)从基因和蛋白水平研究扶桑绵粉蚧与寄主植物之间的互作关系,以进一步分析研究扶桑绵粉蚧的寄主适应性;(3)对不同寄主植物上的抗扶桑绵粉蚧基因进行探索与筛选,为转基因抗虫育种提供理论基础;(4)多方面开展对扶桑绵粉蚧的生物防治研究。
References)
Ahmad F, Akram W, Sajjad A,etal. Management practices against cotton mealybug,Phenacoccussolenopsis(Hemiptera: Pseudococcidae) [J].InternationalJournalofAgricultureandBiology, 2011, 13(4): 547-552.
Ahmed M Z, Ma J, Qiu B L,etal. Genetic record for a recent invasion ofPhenacoccussolenopsis(Hemiptera: Pseudococcidae) in Asia [J].Environmentalentomology, 2015, 44(3): 907-918.
Akintoln A, Ande A. First record ofPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) onHibiscusrosa-sinensisin Nigeria [J].AgriculturalJournal, 2008, 3(1): 1-3.
Amarasekare KG, Chong JH, Epsky ND,etal. Effect of temperature on the life history of the mealybugParacoccusmarginatus(Hemiptera: Pseudococcidae) [J].JournalofEconomicEntomology, 2008, 101(6): 1798-1804.
Arif MI, Rafiq M, Ghaffar A. Host plants of cotton mealybug (Phenacoccussolenopsis): A new menace to cotton agroecosystem of Punjab, Pakistan [J].InternationalJournalofAgricultureAndBiology, 2009, 11(2): 163-167.
Arif MI, Rafiq M, Wazir S,etal. Studies on cotton mealybug,Phenacoccussolenopsis(Pseudococcidae: Homoptera), and its natural enemies in Punjab, Pakistan [J].InternationalJournalofAgricultureandBiology, 2012, 14(4): 557-562.
Ashfaq M, Shah GS, Noor AR,etal. Report of a parasitic wasp (Hymenoptera: Encyrtidae) parasitizing cotton mealybug (Hemiptera: Pseudococcidae) in Pakistan and use of PCR for estimating parasitism levels [J]. Biocontrol Sci Techn, 2010, 20(6): 625-630.
Chen F, Li SL, Lu YY. Major biological parameters of high and low temperature strains of cotton mealybugPhenacoccussolenopsisTinsley[J].GuangdongAgriculturalSciences, 2014,41(24): 83-87. [陈芳, 李慎磊, 陆永跃. 棉花粉蚧高温和低温品系的主要生物学参数研究[J]. 广东农业科学, 2014, 41(24): 83-87]
Culik MP, Gullan PJ. A new pest of tomato and other records of mealybugs (Hemiptera: Pseudococcidae) from Espírito Santo, Brazil [J].Zootaxa, 2005, 964:1-8.
Fand BB, Tonnang HEZ, Kumar M,etal. A temperature-based phenology model for predicting development, survival and population growth potential of the mealybug,PhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) [J].CropProtection, 2014, 55:98-108.
Fuchs T, Stewart J, Minzenmayer R,etal. First record ofPhenacoccussolenopsisTinsley in cultivated cotton in the United States [J].TheSouthwesternEntomologist(USA), 1991, 16(3): 215-21.
Guan X, Lu YY, Zeng L,etal. Supercooling points and freezing points of mealybugPhenacoccussolenopsisTinsley[J].JournalofEnvironmentalEntomology, 2009, 31(4): 381-384. [关鑫, 陆永跃, 曾玲,等. 扶桑绵粉蚧的过冷却点和体液结冰点测定[J]. 环境昆虫学报, 2009, 31(4): 381-384, 394]
Guan X, LU YY, Zeng L,etal. Life tables ofPhenacoccussolenopsisTinsley experimental population at different temperatures[J].AgriculturalScience&Technology, 2012, 13(4): 792-797,814.
Guan X, Lu YY, Zeng L. Study on developmental durations and fecundity ofPhenacoccussolenopsisTinsley on four species of hosts[J].AgriculturalScience&Technology, 2012, 2: 408-411.
Guan X, Lu YY. Study on body length, body width and morphological characteristics ofPhenacoccussolenopsisTinsley fed on cotton[J].JournalofAnhuiAgriculturalSciences, 2012,40(1): 261-262,298. [关鑫, 陆永跃. 棉花上扶桑绵粉蚧各虫态体长、体宽与形态特征[J]. 安徽农业科学, 2012, 40(1): 261-262,298]
Guan X, Zeng L, Hu XN,etal. Lethal effect of hot water on the mealybug,PhenacoccussolenopsisTinsley[J].PlantQuarantine, 2011, 25(1): 1-5. [关鑫, 曾玲, 胡学难, 等. 热水处理对扶桑棉粉蚧的致死作用[J]. 植物检疫, 2011, 25(1): 1-5]
He YB, Wan XW, Zhan RL,etal. Genetic relationship of 12 species of mealybugs (Hemiptera: Pseudococcidae) based on DNA squences[J].ChineseJournalofTropicalCrops, 2011, 32(12): 2324-2330. [何衍彪, 万宣伍, 詹儒林, 等. 基于DNA序列的12种粉蚧亲缘关系分析 [J]. 热带作物学报, 2011, 32(12): 2324-2330]
Hodgson C, Abbas G, Arif MJ,etal.PhenacoccussolenopsisTinsley (Sternorrhyncha: Coccoidea: Pseudococcidae), an invasive mealybug damaging cotton in Pakistan and India, with a discussion on seasonal morphological variation [J].Zootaxa, 2008, 1913: 1-35.
Hu XX, Xiao TG. Review ofPhenacoccussolenopsisTinsley[A].HuangzhongEntomologist, 2012, 8. [胡熙熹, 肖铁光. 扶桑绵粉蚧研究进展[A]. 华中昆虫研究, 2012, 8]
Huang F, Zhang PJ, Zhang JM,etal. Effect of three host plants on the development and reproduction ofPhenacoccussolenopsis[J].PlantProtection, 2011, 37(4): 58-62. [黄芳, 张蓬军, 章金明, 等. 三种寄主植物对扶桑绵粉蚧发育和繁殖的影响[J]. 植物保护, 2011, 37(4): 58-62]
Huang F, Zhao CL, Lu YB. Dynamics ofTremblayaprincepsduring individual development of its hostPhenacoccussolenopsis(Hemiptera: Pseudococcidae)[J].ActaEntomologicaSinica, 2015, 58(10): 1140-1145. [黄芳, 赵春玲, 吕要斌. 共生菌Tremblayaprinceps在扶桑绵粉蚧个体发育中的动态变化[J]. 昆虫学报, 2015, 58(10): 1140-1145.]
Huang YS. Multiple conlparison of the ratio [J].ShanghaiJournalofPreventiveMedicine, 2005, 17(7): 349-350. [黄雨舜.率的多重比较[J]. 上海预防医学, 2005, 17(7): 349-350]
Kumar S, Sidhu JK, Hamm JC,etal. Effects of temperature and relative humidity on the life table ofPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) on cotton [J].FloridaEntomologist, 2013, 96(1): 19-28.
Kumar S, Sidhu JK, Hamm JC,etal. Effects of temperature and relative humidity on the life table ofPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) on cotton [J].FloridaEntomologist, 2013, 96(1): 19-28.
Liu G.Phenacoccussolenopsisis included in the Chinese plant quarantine pest list[J].BeijingAgriculture:PracticalTechnology, 2009, 4:56. [刘刚. 我国将扶桑绵粉蚧列入进境植物检疫性有害生物名录[J]. 北京农业: 实用技术, 2009, 4: 56]
Lu MC. Effect of Different Host Plants on the Development and Reproduction ofPhenacoccussolenopsisTinsley, A New Invasive of China[D]. Zhejiang Normol Universicity. 2013. [吕茂翠. 不同寄主植物对扶桑绵粉蚧生长发育和繁殖的影响[D]. 浙江师范大学, 2013]
Lu YY, Guan X, Zeng L. Effect of temperature on the development of the mealybug,PhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) [J].ScientificResearchandEssays, 2011, 6(31): 6459-6464.
Luo M, Bin SY, Lin JT. Prokaryotic expression and expression profiling of calmodulin genes in mealybugPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae)[J].JournalofNorthwestA&FUniversity(Nat.Sci.Ed.), 2013, 12: 138-142. [罗梅, 宾淑英, 林进添. 扶桑绵粉蚧钙调蛋白基因的原核表达与表达谱分析[J]. 西北农林科技大学学报(自然科学版), 2013, 12: 138-142.]
Luo M, Dong ZY, Bin SY,etal. Molecular cloning and bioinformatics analysis of calmodulin genes inPhenacoccussolenopsisTinsley[J].JournalofHuazhongAgriculturalUniversity, 2012, 31(3): 320-324. [罗梅, 董章勇, 宾淑英, 等. 扶桑绵粉蚧钙调蛋白基因的克隆与生物信息学分析 [J]. 华中农业大学学报, 2012, 31(3): 320-324]
Luo M, Dong ZY, Bin SY,etal. Molecular cloning and expression of different developmental stages of glutathione -S-transfeses gene in mealybugPhenacoccussolenopsisTinsley (Hemiptera:Pseudococcidae)[J].JournalofNorthwestA&FUniversity(Nat.Sci.Ed.), 2012, 9: 83-89. [罗梅, 董章勇, 宾淑英, 等. 扶桑绵粉蚧谷胱甘肽-S-转移酶基因的克隆及不同发育阶段的表达分析 [J]. 西北农林科技大学学报(自然科学版), 2012, 9: 83-89]
Luo M, Dong ZY, Bin SY,etal. Molecular cloning, prokaryotic expression and expression at different developmental stages of cathepsin B gene in mealybugPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) [J].ActaEntomologicaSinica, 2012, 55(3): 276-283. [罗梅, 董章勇, 宾淑英, 等. 扶桑绵粉蚧组织蛋白酶B基因的克隆、原核表达和不同发育阶段表达分析 [J]. 昆虫学报, 2012, 55(3): 276-283]
Luo M, Lin JT, Bin SY. Molecular cloning, sequence characteristics and expression of ring finger protein gene ofPhenacoccussolenopsisTinsley[J].JournalofNorthwestA&FUniversity(Nat. Sci. Ed.), 2014, 4: 125-131. [罗梅, 林进添, 宾淑英. 扶桑绵粉蚧环指蛋白基因的克隆及其序列特征与表达谱分析[J].西北农林科技大学学报(自然科学版), 2014, 4: 125-131]
Luo M, Zhang H, Bin SY,etal. High-throughput discovery of SSR genetic markers in the mealybug,Phenacoccussolenopsis(Hemiptera: Pseudococcidae), from its transcriptome database[J].ActaEntomologicaSinica, 2014, 4: 395-400. [罗梅, 张鹤, 宾淑英, 等. 基于转录组数据高通量发掘扶桑绵粉蚧微卫星引物 [J]. 昆虫学报, 2014, 4: 395-400]
Ma J, Hu XN, Liu HJ,etal.PhenacoccussolenopsisTinsley was found on Hibiscus rosa-sinensis in Guang Zhou[J].PlantQuarantine, 2009, 23(2): 35-36. [马骏, 胡学难, 刘海军,等. 广州扶桑上发现扶桑绵粉蚧[J]. 植物检疫, 2009, 23(2): 35-36]
Mamoonurrashid M, Khattak MK, Abdullah K. Phenological response of cotton mealybug,PhenacoccussolenopsisTinsley (Sternorrhyncha: Pseudococcidae) to three prominent host plants [J].PakistanJ.Zool., 2012, 44(2): 341-346.
Muhammad Z, Ahmed, Jun M,etal. Genetic record for a recent invasion ofPhenacoccussolenopsis(Hemiptera: Pseudococcidae) in Asia[J].EnvironmentalEntomology, 2015, 44(3): 907-918.
Nagrare VS, Kranthi S, Biradar VK,etal. Widespread infestation of the exotic mealybug species,Phenacoccussolenopsis(Tinsley) (Hemiptera: Pseudococcidae), on cotton in India [J].BulletinofEntomologicalResearch, 2009, 99(5): 537-541.
Prasad YG, Prabhakar M, Sreedevi G,etal. Effect of temperature on development, survival and reproduction of the mealybug,PhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) on cotton [J].CropProtection, 2012, 39:81-88.
Qian J, Chen PY. Multiple comparison of multiple sample ratios [J].ChineseJournalofHealthStatistics, 2008, 25(2): 206-212. [钱俊,陈平雁.多个样本率的多重比较[J]. 中国卫生统计, 2008, 25(2): 206-212 ]
Sahito HA, Abro GH. Biology of mealybug,PhenacoccussolenopsisTinsley (Pseudococcidae) on okra and china rose under laboratory conditions [J].PakistanEntomologist, 2012, 34(2): 121-124.
Singh S, Sharma R, Kumar R,etal. Molecular typing of mealybugPhenacoccussolenopsispopulations from different hosts and locations in Punjab, India [J].JournalofEnvironmentalBiology, 2012, 33(3): 539-543.
Sreedevi G, Prasad YG, Prabhakar M,etal. Bioclimatic thresholds, thermal constants and survival of mealybug,Phenacoccussolenopsis(Hemiptera: Pseudococcidae) in response to constant temperatures on hibiscus [J].PLoSONE, 2013, 8(9).
Tanwar RK, Jeyakumar P, Singh A,etal. Survey for cotton mealybug,Phenacoccussolenopsis(Tinsley) and its natural enemies [J].JournalofEnvironmentalBiology, 2011, 32(3): 381-384.
Thomas A, Ramamurthy VV. Morphological and molecular studies on the intraspecific variations between populations of the cotton mealybugPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) [J].EntomologistNews, 2014, 123(5): 339-347.
Tian H, Li XF, Wan FH,etal. Identification ofPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) with species-specific COI (SS_COI) primers[J].ActaEntomologicaSinica, 2013, 56(6): 689-696. [田虎, 李小凤, 万方浩, 等. 利用种特异性COI引物(SS_COI)鉴别扶桑绵粉蚧[J]. 昆虫学报, 2013, 56(6): 689-696]
Vennila S, Deshmukh A, Pinjarkar D,etal. Biology of the mealybug,Phenacoccussolenopsison cotton in the laboratory [J].Journalofinsectscience, 2010, 10(115): 1.
Wang C, Chen F, Lu YY. Development and population growth of the cotton mealybug,Phenacoccussolenopsis(Hemiptera: Pseudococcidae), under different photoperiods[J].ActaEntomologicaSinica, 2014, 57(4): 428-434. [王超, 陈芳, 陆永跃. 不同光周期条件下棉花粉蚧的生长发育和种群增长能力[J]. 昆虫学报, 2014, 57(4): 428-434]
Wang C, Chen F, Lu YY. Study on the male fecundity of an alien invasive pest, cotton mealybugPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae)[J].JournalofEnvironmentalEntomology, 2014, 36(5): 661-666. [王超, 陈芳, 陆永跃.外来入侵害虫棉花粉蚧雄虫生殖能力研究[J]. 环境昆虫学报, 2014, 36(5): 661-666]
Wang C, He MY, Lu YY. Population growth, survival and devolopment ofPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) fed on differentHibiscusrosa-sinensisleaves[J].GuangdongAgriculturalSciences, 2016, 43(5): 107-111. [王超, 何沐阳, 陆永跃.不同类型扶桑叶片上棉花粉蚧存活、发育和增长能力[J]. 广东农业科学, 2016, 43(5):107-111]
Wang FF, Zhu YY, Huang F,etal. Effects of temperature on the development and reproduction ofPhenacoccussolenopsis(Hemiptera: Pseudococcidae)[J].ActaEntomologicaSinica, 2014, 4: 436-442. [王飞飞, 朱艺勇, 黄芳,等. 温度对扶桑绵粉蚧生长发育的影响[J]. 昆虫学报, 2014, 4: 436-442]
Wang QJ, GaoY, Chen T,etal. Studies on fitness and potential hazard of the invasive mealybug,PhenacoccussolenopsisTinsley on five species of plants[J].JournalofEnvironmentalEntomology, 2013, 35(6): 699-706. [王前进, 高燕, 陈婷, 等. 5种植物上扶桑绵粉蚧的适生性及其潜在为害分析[J]. 环境昆虫学报, 2013, 35(6): 699-706]
Wang WL, Effects of Hosts, Temperatures and Relative Humidities on the Growth and Development ofPhenacoccussolenopsisTinsley[D]. Nan Ning: Guangxi University, 2014. [王伟兰. 温湿度和寄主对扶桑绵粉蚧生长发育影响的研究[D]. 南宁: 广西大学, 2014]
Wang YY, Xu ZH, Zhang LL,etal. Developmental duration and life table of the laboratory population ofPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) fed on pumpkin at different temperatures[J].ActaEntomologicaSinica, 2012, 55(1): 77-83. [王莹莹, 徐志宏, 张莉丽, 等. 南瓜寄主上扶桑绵粉蚧不同温度下的发育历期和实验种群生命表[J]. 昆虫学报, 2012, 55(1): 77-83]
Williams D, Willink M. Mealybugs of central and South America [J].Wallingford, 1992, 635.
Wu SA, Zhang RZ. A new invasive pest,Phenacoccussolenopsis, threatening seriously to cotton production[J].ChineseBulletinofEntomology, 2009, 1: 159-162. [武三安, 张润志. 威胁棉花生产的外来入侵新害虫糎T7.50扶桑绵粉蚧[J]. 昆虫知识, 2009, 1: 159-162]
Zhang PJ, Huang F, Zhang JM,etal. The mealybugPhenacoccussolenopsissuppresses plant defense responses by manipulating JA-SA crosstalk[J].ScientificReports, 2015, 5.
Zhang RZ, Wang FX, Wang YX,etal.PhenacoccussolenopsisTinsley(Cotton Mealybug)[M]. Beijing: China Agricultural Scientech Press,2010. [张润志, 王福祥, 王玉玺, 等. 扶桑绵粉蚧 (棉花粉蚧) [M]. 北京: 中国农业出版社, 2010]
Zhao XN, Cui XH, Chen L,etal. Ontogenesis and adaptability of mealybugPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) on different varieties of cotton[J].CottonScience, 2012, 24(6): 496-502. [赵新宁, 崔旭红, 陈玲, 等. 不同类型棉花品种对扶桑绵粉蚧个体发育特征及其适应性的影响 [J]. 棉花学报, 2012, 24(6): 496-502]
Zhao XN. Biological Activity and Mechanism Reseach ofPhenacoccussolenopsisTinsley Damaging Different Varieties of Cotton[D]. Hangzhou: China Jiling University,2012. [赵新宁. 扶桑绵粉蚧为害不同品种棉花的生物学差异及其生理生化机制研究[D]. 杭州: 中国计量学院, 2012]
Zheng T, Cui XH, Wang T,etal. Effects of starvation on survival rate and oviposition of the invasive mealybugPhenacoccussolenopsis(Hemiptera: Pseudococcidae)[J].JournalofBiosafety, 2011, 20(3):239-242. [郑婷, 崔旭红, 汪婷, 等. 饥饿对扶桑绵粉蚧存活率和产卵量的影响[J]. 生物安全学报, 2011, 20(3): 239-242]
Zhu YY, Huang F, Lu YB. Bionomics of mealybugPhenacoccussolenopsisTinsley (Hemiptera: Pseudococcidae) on cotton[J].ActaEntomologicaSinica, 2011, 54(2): 246-252. [朱艺勇, 黄芳, 吕要斌. 扶桑绵粉蚧生物学特性研究[J]. 昆虫学报, 2011, 54(2): 246-252]
Review of research on the biological influences ofPhenacoccussolenopsisfeeding on different plants and cotton cultivars
LU Dong-Dong, SHANG Han-Wu*
(College of Life Sciences, China Jiliang Univercity, HangZhou 310018, China)
Phenacoccussolenopsispolyphagous pest, has a wide host range and strong adaptability. It was found in many countries and regions around the world, causing serious harm to agricultural production. Through research analysis of literatures aboutP.solenopsison different host plants, we could find suitable and unsuitable host plants, to adjust farming way timely, and make reasonable rotation, to prevent the pest from spreading; and these results further made clear the research direction ofP.solenopsisin the future. Research results on the morphological, reproductive characteristics, resistance and control ofP.solenopsiswere reviewed by collecting and analyzing the related literatures. And we analyzed comprehensively the changes ofP.solenopsison developmental duration, longevity and fecundity changes in cotton cultivars. Followed by Fuso and cotton,SolanumtuberosumandAmbrosiaartemisiifoliaare very conducive to the proliferation of mealybug.By analyzing ten varieties of cotton, we found that CCRI59 had better resistance toP.solenopsis, and Bt cotton RCH-134 didn’t have resistance. Morphological study ofP.solenopsisis more comprehensive, and molecular genetics, biological control and Insect-resistant breeding study toP.solenopsiswill be the important research fields in the future.
Phenacoccussolenopsis; host plants;cotton
国家重点基础研究发展计划(973计划)(2010CB126200)
鹿东东,女,1989年生,山东潍坊人,硕士研究生,研究方向为生物化学与分子生物学,E-mail:ldd617@163.com
Author for correspondence:商晗武,E-mail:hwshang@cjlu.edu.cn
2016-03-31;接受日期Accepted:2016-07-15
Q965; S433
A
1674-0858(2016)04-0669-11
