围封对荒漠草原土壤微生物数量及其酶活性的影响
2016-08-10罗冬王明玖郑少龙李元恒内蒙古农业大学生态环境学院内蒙古呼和浩特0009内蒙古通辽市奈曼旗林业局内蒙古通辽0800中国农业科学院草原研究所内蒙古呼和浩特0000
罗冬,王明玖*,郑少龙,李元恒. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 0009;. 内蒙古通辽市奈曼旗林业局,内蒙古 通辽 0800;. 中国农业科学院草原研究所,内蒙古 呼和浩特 0000
围封对荒漠草原土壤微生物数量及其酶活性的影响
罗冬1,王明玖1*,郑少龙2,李元恒3
1. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2. 内蒙古通辽市奈曼旗林业局,内蒙古 通辽 028300;3. 中国农业科学院草原研究所,内蒙古 呼和浩特 010010
摘要:以鄂尔多斯高原温性荒漠草原为对象,对比研究了放牧和围封条件对草原植物地下生物量、土壤有机质、土壤微生物数量及酶活性的影响,旨在为干旱区退化草地的恢复与重建提供科学依据。结果表明,(1)与放牧区相比,围封显著增加了表层土壤(0~10 cm)草地植物地下生物量23.81%,极显著增加了次表层土壤(10~20 cm)草地植物地下生物量37.21%,而对土壤有机质含量无显著影响。(2)围封极显著提高了表层土壤真菌种群数量71%,次表层土壤细菌种群数量147%,显著提高了底层土壤(20~40 cm)真菌164%和表层、次表层土壤放线菌种群数量56%和8.9%。(3)围封区的次表层土壤脲酶活性显著增加了 31.13% ,表层土壤蔗糖酶活性显著降低了 24.14%,表层土壤过氧化氢酶和碱性磷酸酶活性分别显著提高了16.59%和11.76%。(4)围封与放牧条件下,各指标相关性表现不尽一致;放牧条件下,放线菌种群数量与各指标相关性均不显著(P>0.05),而在围封下,放线菌与真菌种群数量及碱性磷酸酶活性无显著相关性(P>0.05),与其他指标均表现出显著(P<0.05)或极显著(P<0.01)正相关关系,相关系数达68%以上;放牧条件下,真菌种群数量与4种土壤酶活性均呈极显著(P<0.01)正相关关系,相关系数在84%以上,而围封条件下,其与4种酶活性均呈显著正相关关系(P<0.05)。
关键词:荒漠草原;土壤地下生物量;土壤有机质;微生物数量;土壤酶活性
引用格式:罗冬, 王明玖, 郑少龙, 李元恒. 围封对荒漠草原土壤微生物数量及其酶活性的影响[J]. 生态环境学报, 2016, 25(5): 760-767.
LUO Dong, WANG Mingjiu, ZHENG Shaolong, LI Yuanheng. Effects of Enclosure on Soil Microbial Quantity and Enzyme Activity in Desert Steppe [J]. Ecology and Environmental Sciences, 2016, 25(5): 760-767.
荒漠草原是干旱性最强的草原生态系统,在温带草原中占重要地位(韩国栋等,2007)。与其他类型草原相比,该生态系统稳定性差,服务功能下降明显,部分地段承受和抵御干扰的能力十分脆弱,有向逐渐裸露化或荒漠化发展的趋势(赛胜宝等,1995)。地处我国北方西部内陆气候与东部季风气候过渡带上的鄂尔多斯高原荒漠草原(任健美等,2005),因其高寒、干旱、风大沙多等生境及长期不合理利用,导致该类草地退化严重,成为了草地恢复研究的关键区域(姜联合,2005)。近年来,鄂尔多斯高原荒漠草原干旱化趋势尤为明显(韩芳等,2013),土壤蓄水保肥能力减弱,植被稀疏,物种多样性丧失,草地生产力下降。如何对退化草地进行恢复治理,恢复土壤肥力,对于保护当地生态,实现草地资源的永续利用具有重要的理论和现实意义。
围栏封育,一直被认为是草地管理和退化草地恢复的经济而有效的措施。以往对封育条件下荒漠草原恢复机理的研究多集中于植物净初级生产力(侯向阳等,2014)、土壤理化性质(李学斌等,2015)、物种多样性(王蕾等,2012)以及土壤种子库(沈艳等,2015)等方面,而关于土壤微生物学特性方面的研究相对较少(廖仰南,1993;红梅等,2009)。近年来,随着人们对微生物在整个生态系统中的重要作用的认识,愈来愈多的研究侧重于用微生物参数来评价土壤肥力和质量状况(王少昆等,2013;赵帅等,2011),但由于荒漠草原分布的广泛性和复杂性,研究结果尚无统一定论(牛得草等,2013;马文文等,2014;陈鸿洋等,2015),而对鄂尔多斯荒漠草原恢复过程中土壤微生物的研究更是鲜有报道。本研究以鄂尔多斯市鄂托克旗荒漠草原为对象,对比分析放牧和围封条件下草原的植物地下生物量、土壤有机质、土壤微生物数量、酶活性指标及各指标间的相关关系,旨在探讨封育在退化荒漠草原恢复过程中的贡献效果,为退化草地的改良提供科学依据。
1 材料与方法
1.1 研究区概况
试验区位于内蒙古自治区鄂尔多斯市鄂托克旗阿尔巴斯苏木(乡),地理坐标 N39°11′16",E107°55′41",海拔高度1441 m,年平均降水量约为265 mm,且主要集中在6—8月,约占全年降水量的63%,年平均蒸发量2470 mm。年平均风速在3 m·s-1以上;年平均沙尘暴日为9 d,集中于春季。年平均温度为 7.2 ℃,≥10 ℃的年积温为2800~3500 ℃,年平均日照2800~3100 h,无霜期平均122 d。土壤为淡棕钙土,植被低矮稀疏,平均高度为9 cm,盖度为18%~25%,种类组成贫乏。建群种为短花针茅(Stipa breviflora),在群落中起重要作用的植物为狭叶锦鸡儿(Caragana stenophylla)、无芒隐子草(Cleistogenes songorica)。主要伴生种有寸草苔(Carex duriuscula)、白草(Pennisetum centrasiaticum)、蒙古韭(Allium mongolicum)、阿尔泰狗娃花(Heteropappus altaicus)和牛枝子(Lespedeza davurica)等。栉叶蒿(Neopallasia pectinata)、砂蓝刺头(Echinops gmelini)、猪毛菜(Salsola collina)等一、二年生植物也很常见。
1.2 研究方法
1.2.1 样地设置及样品采集
于2012年4月,在试验区内选取地势较平坦、土壤和植被基本一致的区域,设置围栏。在围栏内外各随机设置6个固定样点,即为放牧处理区(CK)和围封处理区(WF)。于2014年(封育第3年)8月初,用内径8 cm根钻测取0~10、10~20和20~40 cm土层的植物地下生物量,每样点 3次重复,同层均匀混合,待清洗测定;土样采用对角交叉法按上述土层深度进行取样,每样点5次重复,同层均匀混合,取1 kg左右土样装入无菌聚乙烯袋并用生物冰袋冷藏后,带回实验室,将每份土样分为2份,其中一份用于微生物数量测定,快速(4 ℃下保存不超过24 h)进行分离培养、计数;另一份土样精细去除可见植物残体(根、茎、叶、凋落物),过筛(2 mm)后,在室温下风干,用于土壤有机质及土壤酶活性的测定。
1.2.2 样品分析
将含有根系的土壤放入尼龙网袋中用水冲洗,对根系进行分离,并在65 ℃下烘干至恒重后换算成单位面积的量,即植物地下生物量;土壤有机质采用重铬酸钾氧化法测定;土壤微生物数量测定采用稀释涂布平板计数法(沈萍等,2001)。
真菌的培养基为马丁-孟加拉红,即称取 10 g土样,在无菌条件下用无菌水配成不同浓度梯度悬浮液,取稀释度为10-1、10-2、10-3的土壤悬浮液各50 μL,接种于盛有灭菌培养基的培养皿中,用无菌刮刀涂抹均匀。每个浓度 3次重复,25 ℃恒温培养5~7 d,选取每皿菌落数为15~150的1个稀释度统计菌落数,按如下公式计算真菌数量。
菌数=菌落平均数×稀释倍数/干土重
放线菌以高氏一号为培养基,除悬浮液稀释倍数为10-3、10-4、10-5外,其余与真菌数量测定方法相同。恒温28 ℃恒温培养7~10 d,按上述方法和公式统计菌落数并计算放线菌数量。
细菌以牛肉膏蛋白胨琼脂为培养基除28 ℃恒温培养3 d统计菌落数外,其余与放线菌数量测定方法相同。
酶活性主要测定了脲酶、蔗糖酶、过氧化氢酶和碱式磷酸酶,分别以1 g干土37 ℃条件下培养24 h后所生成的NH3-N的质量数(mg)、释放葡萄糖的质量数(mg)、消耗0.1 mol·L-1高锰酸钾的体积数(mL)和生成苯酚的质量数(mg)(关松荫,1986)为计算指标,分别采用苯酚-次氯酸钠比色法、3,5-二硝基水杨酸比色法、高锰酸钾滴定法和磷酸苯二钠比色法进行测定。
1.3 数据分析
分别用Excel 2003和spss 19.0软件作图和数据统计分析。各指标放牧与围封处理间的比较及同一处理不同土层间的比较均采用单因素ANOVA统计分析,相关性分析采用Pearson's法。
2 结果与分析
2.1 围封对植物地下生物量及土壤有机质含量的影响
CK和WF区的植物地下生物量均随土层深度增加而显著减少(P<0.05)。对表层土壤植物地下生物量分析得,WF较 CK区显著增重 23.81% (P<0.05);在次表层土壤中,WF比 CK区增重37.21%,并达到极显著水平(P<0.01);底层土壤植物地下生物量在两区域内,无显著差异(P>0.05)。就土壤有机质含量而言,CK区表层土壤的有机质含量显著高于下两层(P<0.05),下两层差异不显著(P>0.05);WF区的土壤有机质含量随土层深度增加而显著减少(P<0.05);两区域同层土壤有机质含量均无显著差异(P>0.05)(表1)。
2.2 围封对土壤微生物数量的影响
WF与CK区的草地土壤,均以细菌种落数量占绝对优势,其次为放线菌,真菌最少,即细菌>放线菌>真菌。两区域次表层土壤细菌种群数量存在极显著差异(P<0.01),WF较CK区提高了147%;WF区的表层土壤真菌种群数量较CK提高了71%,且达到极显著水平(P<0.01),底层显著提高了164% (P<0.05);表层和次表层土壤放线菌种群数量,WF 较CK区分别提高了56%和8.9%,且达到显著水平(P<0.05);在次表层土壤上,WF区微生物总数与CK区存在极显著差异(P<0.01),较 CK提高了145%,其他层次土壤的菌种数量,在两区域内均无显著差异(P>0.05)。同一区域不同土层深度间,在CK区,除放线菌外,其他菌种及各土层微生物总数均表现为表层显著高于下两层(P<0.05),下两层无显著差异(P>0.05);WF区的细菌种群数量和各土层微生物总数随土层深度增加呈显著递减(P<0.05),真菌种群数量为表层显著高于下两层,放线菌为底层显著低于上两层(P<0.05);依3层土壤微生物群落数量之和来看,WF较 CK区提高25.97%(表2)。
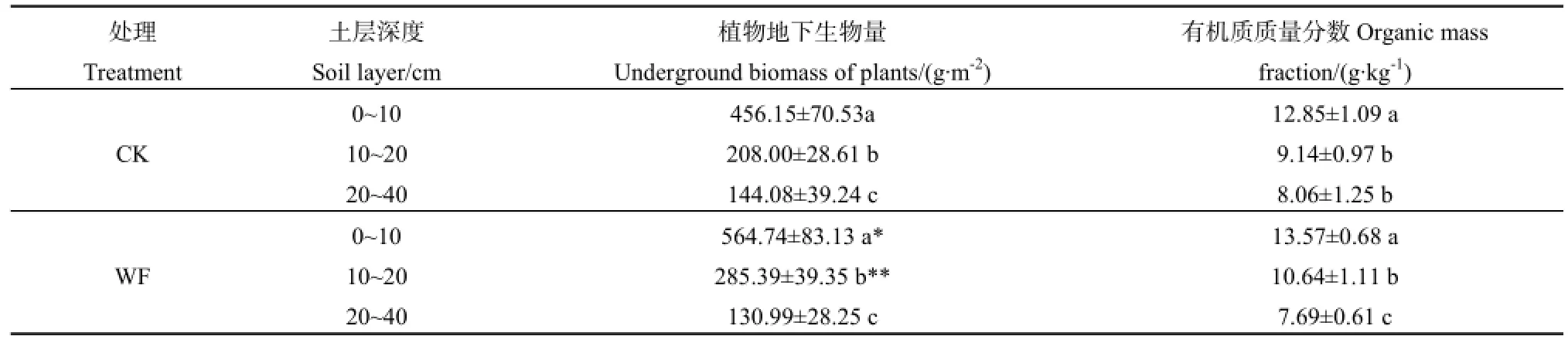
表1 围封对地下生物量及土壤有机质质量分数的影响Table 1 The influence of belowground biomass and soil organic mass fraction under enclosure

表2 围封对土壤3大类微生物种群数量的影响Table 2 The influence of the three main types of soil microbial populations under enclosure
2.3 围封对土壤酶活性的影响
本研究中,4种土壤酶活性对WF的响应不尽一致。退化草地围封3年后的表层土壤,除脲酶外,其他 3种酶的活性与 CK区均存在显著差异(P<0.05),WF较CK区,蔗糖酶活性降低了24.14%、过氧化氢酶和碱性磷酸酶活性分别提高了 16.59% 和11.76%;在次表层土壤中,两区域的脲酶和蔗糖酶活性存在显著差异(P<0.05),WF较CK区分别提高了31.13%和17.23%,其他2种酶均无显著差异(P>0.05);在底层土壤中,两区域的 4种酶活性均无显著差异(P>0.05)。就同区域不同土层深度而言,WF区的土壤脲酶和蔗糖酶活性均表现为随土层增加而显著降低(P<0.05),过氧化氢酶和碱性磷酸酶活性均表现为表层显著高于下两层(P<0.05),且下两层无显著差异(P>0.05)。CK区,除过氧化氢酶活性表现为随土层深度增加而显著降低外(P<0.05),其他 3种酶活性均表现为表层显著高于下两层(P<0.05),下两层差异不显著(P>0.05)(图1)。
2.4 相关性分析
对温性荒漠草原不同利用方式草地的植物地下生物量、土壤有机质质量分数、土壤微生物数量及酶活性进行相关分析(表3)。在CK区,放线菌与其他指标相关性不显著(P>0.05);细菌群落数量与蔗糖酶活性呈显著正相关关系(P<0.05),相关系数为86.6%;其他各指标间均呈极显著正相关关系(P<0.01),相关系数达到81%以上。在WF区,地下生物量除与放线菌群落数量呈显著正相关(P<0.05),相关系数为72.8%,与其他指标均呈极显著正相关关系(P<0.01),相关系数达到了 80%以上;有机质与真菌呈显著正相关(P<0.05),相关系数为70.2%,与其他指标均呈极显著正相关关系(P<0.01),相关系数达71%以上;细菌与放线菌群落数量呈显著正相关关系(P<0.05),相关系数达为73.3%,其他指标均呈极显著正相关关系(P<0.01),相关系数达83%以上;真菌除与放线菌群落数量无显著关系外(P>0.05),与4种土壤酶活性均呈显著正相关关系(P<0.05),相关系数达 66%以上;放线菌群落数量与脲酶和蔗糖酶活性呈极显著正相关关系(P<0.01),与过氧化氢酶活性呈显著正相关关系(P<0.05);除蔗糖酶和碱性磷酸酶呈显著正相关关系(P<0.05)外,其他土壤酶活性均呈极显著正相关关系(P<0.01),相关系数达85%以上。
3 讨论
3.1 围封对植物地下生物量及土壤有机质质量分数的影响
植物地下生物量是指存在于草地植被地表下草本根系和根茎生物量的总和(黄德华等,1988),是草地植被碳蓄积的重要组成部分,草地生物量的80%以上集中在地下(Mokany et al.,2005),它是土壤有机碳库的最主要输入源,准确测定地下生物量的变化是探索不同草地利用方式效果的基础。严格来说,地下生物量应该是本生长季新生的生物总量,但事实上,多年生草本植物含有大量的他年地下活体,当年新生部分也在不断死亡,很难严格加以区分。因此,这里的地下生物量是包含活的生物量和死的生物量积累量,即群落所有植物的根系及地下繁殖体。草地利用方式(放牧、围封、刈割等)、环境因子的干扰(火灾、干旱等)以及环境条件(气候条件、土壤类型等)都会对草原生态系统地下生物量产生影响(Collins et al.,2005;Hobbs et al.,2006)。本研究中,退化草地经 3年围封后,表层地下生物量显著高于CK区(P<0.05);次表层达到极显著水平(P<0.01),表明围封促进了荒漠草地地下生物量的累积。刘建军等(2005)对蒙古克氏针茅(Stipa krylovii)-冷蒿(Artemisia frigida)草原研究表明,围封较自由放牧提高了地下生产力,这一结论在本研究中得到证实。这可能是围封有效地控制了牲畜对植物的采食与践踏,促进了退化草地植物的生长,植物的光合面积及强度增加,营养传输量增大,从而促使地下生物量的积累。从同一处理不同土层来看,宇万太等(2001)对草地地下生物量研究发现,80%的地下根量分布于浅层土壤,并随土层深度呈指数下降。本研究中,两区域的草地地下生物量均随土层深度的增加而剧减,即垂直分布呈“T”型,与上述学者的研究结果基本一致,这可能是因为大部分土壤有机质和养分都储存于土壤表层,植物发展主要根系于该层以尽量获取更多的资源以满足生长需求。大量研究得出,放牧使得地下生物量出现浅层化趋势。本试验中,围封三年后草地地下生物量浅层化仍存在,根系浅层化是否能长期应对荒漠草原的环境变化,围封年限的合理化范围等问题还需进一步研究。
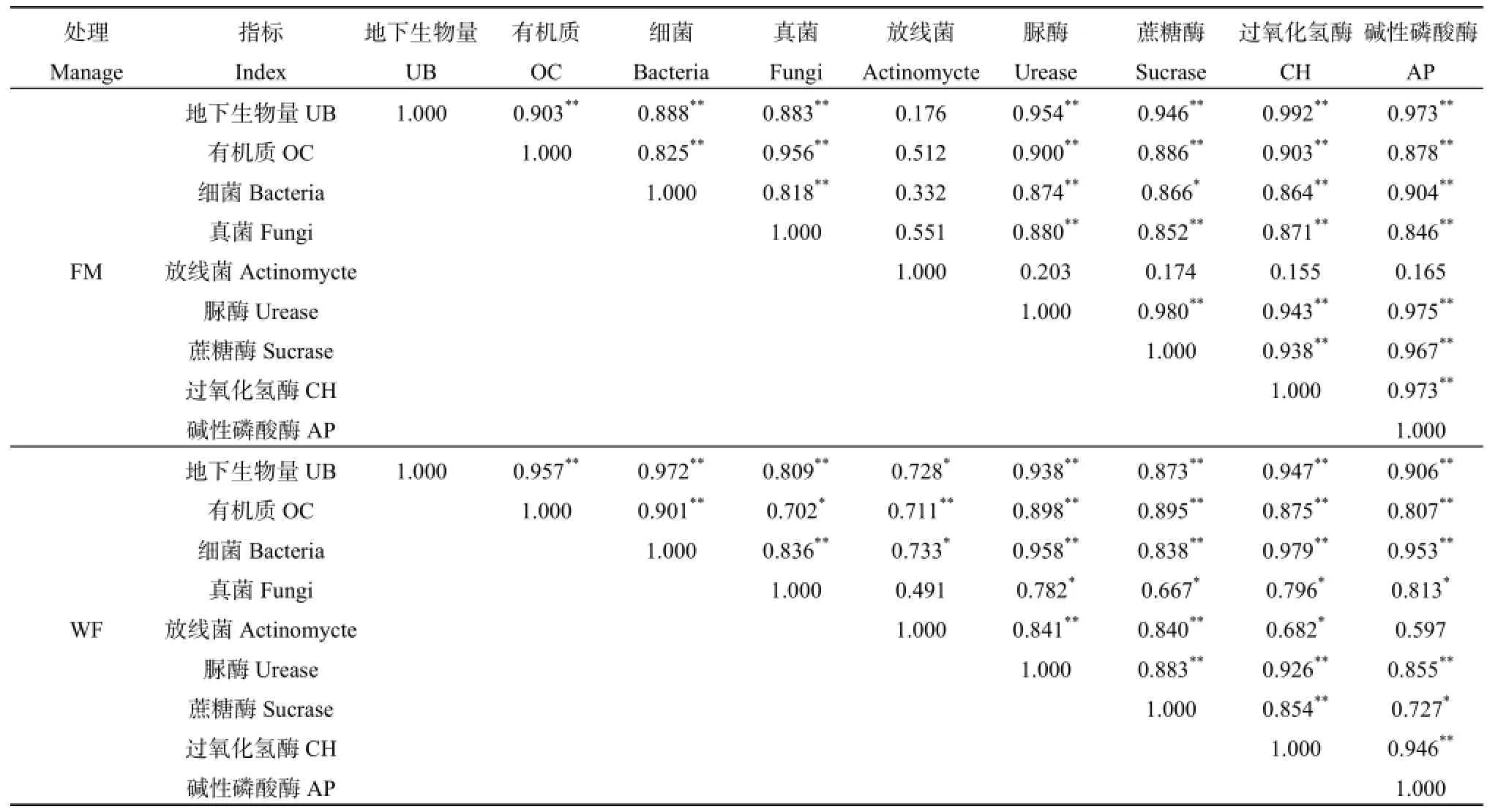
表3 各指标之间的相关性分析Table 3 Correlation analysis among the various indicators
土壤有机质是指进入土壤中的各种有机物质,在土壤微生物作用下形成的一系列有机化合物的总称,其来源主要包括微生物、植物、动物和人类活动。在自然条件下,地面植被残体和根系是土壤有机质的主要来源(吕贻忠等,2006)。杨合龙等(2015)采用封育措施探究伊犁绢蒿荒漠草地土壤养分变化的研究表明,随封育年限的增加,土壤有机质呈先降后升变化;程杰等(2007)对云雾山封育 23年的天然草地研究表明土壤有机质含量显著高于对照。本试验中,退化草地围封3年后与CK区同层相比,土壤有机质含量并未显著提高(P>0.05),这可能是因为有机质本身变化比较缓慢,难以及时反映土壤受干扰时的各种动态,加之围封年限较短,围封处理未能体现预期效果。同一区域不同土层,土壤有机质含量呈明显垂直分布,这可能是地表残体被分解后,因表土根系发达而吸收较多养分,随土层深度的增加,养分剩余量减少造成。
3.2 围封对微生物数量的影响
土壤微生物是土壤有机质的活性部分(Jenkinson et al.,1981),在土壤质量的演变过程中具较高的转化能力(Powlson et al.,1987),是土壤有效养分的贮存库(薛青芳等,2007)。在陆地生态系统中,土壤微生物主要扮演“分解者”的角色,几乎参与土壤中一切生物化学反应,发挥着地球C、N、P、S等物质循环的“调节器”、土壤养分植物有效性的“转化器”和污染环境的“净化器”等多方面生态功能(程杰等,2007)。其分布与活动,一方面反映了环境、生物因子对微生物的分布习性、群落组成、种群演替及功能的影响,另一方面说明微生物自身对植物生长、土壤肥力及物质循环和能量转化的现状和趋势的响应。本研究中,围封措施显著提高了底层真菌和表层、次表层放线菌的群落数量;极显著提高了次表层细菌和表层真菌的群落数量。这与邵玉琴等(2004)对恢复草地和退化草地土壤微生物类群数量分布特征的研究中得出的围封样地各土层中土壤微生物各类群数量均高于退化草地的结果基本吻合。原因:一方面可能是围封后植被的恢复有效防止了降雨引起的溅蚀和地表径流侵蚀,增加了草地土壤的保墒力;另一方面可能是地上植被的恢复及根系的增长,增加了土壤的养分输入量,这些条件的改善给微生物的存活与生长提供了更好的条件。同一区域不同土层看,细菌在微生物总数中占绝对优势,因此细菌的变化趋势基本决定了微生物总数的变化趋势。谭秋锦等(2014)对不同生态系统土壤微生物数量及生物量特征的研究表明,细菌在微生物群落中占绝对数量,放线菌次之,真菌数量最少。这可能由于细菌种群组成的多样性和对不同时期植物残体分解利用过程中各种群之间的接力更替有关。土壤微生物群落数量集中分布于土壤表层并表现出明显的垂直分布特性,原因可能是草地表土有机质丰富,植物根系的分泌物含量高,为微生物的繁殖提供了营养,有利于土壤微生物的生存和繁殖。本研究的不足之处在于未能细化各菌群代表菌落,如芽孢杆菌、链霉菌等,如有条件应深入研究以提供更精确的机理依据。
3.3 围封对土壤酶活性的影响
土壤酶类作为有机质分解过程的生物催化剂(向泽宇等,2011),主要来自于土壤微生物和植物根系(文都日乐等,2010),对外界环境因素引起的变化极为敏感并能够在短期内发生变化,可以作为衡量生态系统土壤质量变化的预警和敏感指标(Dick,1994;陈利军等,2002)。其活性强弱可表征土壤特定生化反应的方向和强度,对维持土壤生态系统的稳定起着重要作用(闫瑞瑞等,2011)。土壤脲酶直接参与土壤含氮化合物的分解与转化,土壤碱性磷酸酶的活性高低直接影响着土壤中有机磷的分解、转化及其生物有效性,二者的活性分别表征土壤的氮素和磷素的供应状况,与土壤肥力直接相关(文都日乐等,2010)。本研究中,围封区次表层脲酶活性与表层碱性磷酸酶活性显著高于放牧区(P<0.05),其原因可能是草地围封后,次表层根系及菌群数量显著增加,使得有机氮分解强度更高,促使该层脲酶活性有较大提高,使得脲酶源表现为根系和微生物两方面;而表层碱性磷酸酶活性提高可能是表层凋落物分解,土壤肥力增强,进而促进其活性提高,使得其来源偏重于根系。蔗糖酶活性与草地有机质的转化和呼吸强度有关,本试验中,围封导致了土壤表层蔗糖酶活性下降,其原因可能是围封促使土表凋落物增多,表层土壤通气性降低,致使表土的呼吸强度减弱;而放牧区有机质累积量不足,蔗糖酶的活性较强,也表现出草地生物活性的适应性调节能力。过氧化氢酶反映了土壤腐殖化强度和有机质积累程度。本试验中,围封区土壤表层过氧化氢酶活性显著高于放牧区,其原因可能是土表残留物增多,为提高腐殖化程度及有机质累积速率,其活性随之增强。围封对土壤酶的活性总体上有较显著的提高作用,这与牛得草等(2013)、闫瑞瑞等(2011)研究结果基本一致,但土壤深层的酶活性不因围封而发生变化,是否与围封年限或酶种选择有关,有待进一步研究。
3.4 各指标之间的关系
关于土壤微生物数量和土壤酶活性相关性的报道较多,但结果不尽一致,这可能与研究区域土壤类型及草地利用方式不同有关。马文文等(2014)在研究荒漠草原2种植物群落土壤微生物及土壤酶特征中发现,盐爪爪(Kalidium foliatum)群落草地土壤细菌与脲酶、磷酸酶呈极显著正相关关系,与蔗糖酶、过氧化氢酶呈显著正相关关系,骆驼刺(Alhagi sparsifolia)群落草地土壤细菌与蔗糖酶活性、过氧化氢酶活性呈极显著正相关关系,与脲酶活性、磷酸酶活性呈显著正相关关系。同样,吕桂芬等(2008)在内蒙古不同类型荒漠草原土壤微生物数量及其酶活性研究中得出,蔗糖酶与过氧化氢酶呈负相关关系但不显著。而闫瑞瑞等(2011)在不同放牧梯度下草甸草原土壤微生物和酶活性研究中得到,土壤微生物数量与蔗糖酶、过氧化氢酶呈显著正相关关系。本研究除土壤微生物数量和酶活性外,还补充了植物地下生物量及土壤有机质质量分数,较系统地阐述了不同草地利用方式下,植物地下部分和土壤多层次营养指标的变化,相关研究鲜有报道。本研究发现,放牧条件下的放线菌种群数量与其他指标均无显著相关关系(P>0.05),而围封后,放线菌与地下生物量、细菌群落数量、过氧化氢酶活性均呈显著正相关关系(P<0.05),与有机质含量、脲酶和蔗糖酶活性呈极显著正相关关系(P<0.01),出现上述结果可能主要与两种草地利用方式下土壤微生物群落结构的变化有关。另一方面,土壤有机质质量分数并未随利用方式的改变而变化,但与其他指标的相关性却很显著。综上所述,土壤有机质、土壤微生物数量与酶活性等存在复杂关系,不能笼统地确定土壤微生物数量提高有助于酶活性的增强,有机质含量与微生物数量或酶活性有直接关系等观点,这类问题的研究,需要进行长期试验或将一种土壤酶与特定分泌此类酶的微生物类群对应分析,才能从机理上回答各指标之间的关系。未来应进一步探究土壤酶活性降低是由分泌此酶的微生物数量引起的,还是由酶的活性基团减少引起等问题。
4 结论
对鄂尔多斯荒漠草地实施3年围封措施后,显著提高了草地表层和次表层土壤植物地下生物量,而对底层土壤植物地下生物量无显著影响;土壤有机质含量无显著变化;三大微生物种群数量在土壤表层和次表层均有显著或极显著增加,在底层土壤中,只有真菌种群数量显著增加;土壤脲酶在次表层有显著提高,蔗糖酶在表层显著下降,过氧化氢酶和碱性磷酸酶活性在表层显著提高,底层土壤中4种酶的活性无显著差异。各指标值均表现出较强的垂直分布特性。不同处理下,草地植物地下生物量、土壤有机质质量分数、土壤微生物数量及酶活性存在不同程度的相关关系。
参考文献:
COLLINS S L, SMITH M D. 2005. Scale-dependent interaction of fire and grazing on community heterogeneity in tall grass prairie [J]. Ecology,87(8): 2058-2067.
DICK R P. 1994. Soil enzyme activities as indicators of soil quality [C]// DORAN J W, COLEMAN D C, BEZDICEK D F, et al. Defining Soil Quality for a Sustainable Environment [J]. Soil Science, 159(1): 7.
HOBBS N T, SCHIMEL D S, OWENSBY C E. 1991. Fire and grazing in the tall grass prairie: contingent effects on nitrogen budgets [J]. Ecology, 72(4): 1374-1382.
JENKINSON D S, LADD J N. 1981. Microbial biomass in soil: Measurement and turnover [A]//Paul V E A, Ladd J N. Soil Biochemistry [M]. New York: Marcel Dekker Inc.
MOKANY K, RAISON R J, PROKUSHKIN A S. 2005. Critical analysis of root: shoot ratios in terrestrial biomes [J]. Global Change Biology,12(1): 84-96.
POWLSON D S, BROOKS P C, CHRISTENSEN B T. 1987. Measurement of soil Microbial biomass provides an early indication of changes in total soil organic matter due to straw incorporation [J]. Soil Biology and Biochemistry, 19(2): 159-164
陈鸿洋, 尚振艳, 傅华, 等. 2015. 荒漠区不同大小灌丛周围土壤微生物生物量及活性特征[J]. 草业学报, 24(2): 70-76.
陈利军, 武志杰. 2002. 与氮转化有关的土壤酶活性对抑制剂施用的响应[J]. 应用生态学报, 13(9): 1099-1103.
程杰, 高亚军. 2007. 云雾山封育草地土壤养分变化特[J]. 草地学报,15(3): 273-277.
关松荫. 1986. 土壤酶及其研究法[M]. 北京: 农业出版社.
韩芳, 刘朋涛, 牛建明, 等. 2013. 50a来内蒙古荒漠草原气候干燥度的空间分布及其演变特征[J]. 干旱区研究, 30(3):449-456.
韩国栋, 焦树英, 毕力格图, 等. 2007. 短花针茅草原不同载畜率对植物多样性和草地生产的影响[J]. 生态学报, 27(1): 182-188.
红梅, 敖登高娃, 李金霞, 等. 2009. 荒漠草原土壤健康评价[J]. 干旱区资源与环境, 23(5): 117-120.
侯向阳, 纪磊, 王珍. 2014. 荒漠草原与典型草原NPP、碳积累对不同降雨年份和利用方式的响应[J]. 生态学报, 34(21): 6256-6264.
黄德华, 陈佐忠, 张鸿芳. 1988. 贝加尔针茅、克氏针茅、线叶菊草原地下生物量的比较研究[A]//草原生态系统研究(第2集)[C]. 北京: 科学出版社.
姜联合. 2005. 鄂尔多斯高原飞播造林物种种苗生物学及植被恢复优化方案研究[D]. 北京: 北京林业大学.
李学斌, 陈林, 樊瑞霞, 等. 2015. 围封条件下荒漠草原4种典型植物群落枯落物输入对土壤理化性质的影响[J]. 浙江大学学报(农业与生命科学版), 41(1): 101-110.
廖仰南, 赵吉, 杜建军, 等. 1993. 内蒙古草原与荒漠地带土壤微生物的生态分布特性[J]. 植物学通报, 10(增刊): 39-41.
刘建军, 浦野忠朗, 鞠子茂, 等. 2005. 放牧对草原生态系统地下生产力及生物量的影响[J]. 西北植物学报, 25(1): 88-93.
吕桂芬, 吴永胜, 李浩, 等. 2008. 内蒙古不同类型荒漠草原土壤微生物数量及其酶活性研究[J]. 内蒙古师范大学学报, 37(6): 761-764.
吕贻忠, 李保国. 2006. 土壤学[M]. 北京: 中国农业出版社.
马文文, 姚拓, 靳鹏, 等. 2014. 荒漠草原2种植物群落土壤微生物及土壤酶特征[J]. 中国沙漠, 34(1): 176-183.
牛得草, 江世高, 秦燕, 等. 2013. 围封与放牧对土壤微生物和酶活性的影响[J]. 草业科学, 30(4): 528-534.
任健美, 尤莉, 高建峰, 等. 2005. 鄂尔多斯高原近40a气候变化研究[J].中国沙漠, 6(11): 82-87.
赛胜宝, 李德新. 1995. 荒漠草原生态系统研究[M]. 呼和浩特: 内蒙古人民出版社.
邵玉琴, 赵吉, 杨劼. 2004. 恢复草地和退化草地土壤微生物类群数量的分布特征[J]. 中国沙漠, 24(2): 111-114.
沈萍, 范秀容, 李广武. 2001. 微生物学实验[M]. 北京: 高等教育出版社.
沈艳, 刘彩凤, 马红彬, 等. 2015. 荒漠草原土壤种子库对草地管理方式的响应[J]. 生态学报, 35(14): 4725-4732.
谭秋锦, 宋同清, 彭晚霞, 等. 2014. 峡谷型喀斯特不同生态系统的土壤微生物数量及生物量特征[J]. 生态学报, 34(12): 3302-3310.
王蕾, 许冬梅, 张晶晶. 2012. 封育对荒漠草原植物群落组成和物种多样性的影响[J]. 草业科学, 29(10): 1512-1516.
王少昆, 赵学勇, 张铜会, 等. 2013. 造林对沙地土壤微生物的数量、生物量碳及酶活性的影响[J]. 中国沙漠, 33(2): 529-535.
文都日乐, 张静妮, 李刚, 等. 2010. 放牧干扰对贝加尔针茅草原土壤微生物与土壤酶活性的影响[J]. 草地学报, 18(4): 517-522.
向泽宇, 王长庭, 宋文彪, 等. 2011. 草地生态系统土壤酶活性研究进展[J]. 草业科学, 28(10): 1801-1806.
薛青芳, 高艳梅, 汪景宽, 等. 2007. 土壤微生物量碳氮作为土壤肥力指标的探讨[J]. 土壤通报, 38(2): 247-250.
闫瑞瑞, 闫玉春, 辛晓平. 2011. 不同放牧梯度下草甸草原土壤微生物和酶活性研究[J]. 生态环境学报, 20(2): 259-265.
杨合龙, 孙宗玖, 管光玉, 等. 2015. 封育对伊犁绢蒿荒漠草地土壤养分的影响[J]. 中国草地学报, 37(2): 107-111.
宇万太, 于永强. 2001. 植物地下生物量研究进展[J]. 应用生态学报,12(6): 927-932.
赵帅, 张静妮, 赖欣, 等. 2011. 放牧与围封对呼伦贝尔针茅草原土壤酶活性及理化性质的影响[J]. 中国草地学报, 33(1): 71-76.
DOI:10.16258/j.cnki.1674-5906.2016.05.005
中图分类号:X172
文献标志码:A
文章编号:1674-5906(2016)05-0760-08
基金项目:公益性行业(农业)科研专项(201003061)
作者简介:罗冬(1986年生),男,博士研究生,主要从事恢复生态学、生态经济可持续发展方面的研究。E-mail: luodong8611@163.com
*通信作者:王明玖(1961年生),男,教授,博士,博士生导师,主要从事草地管理和草地生态方面教学与科研工作。E-mail: wangmj_0540@163.com
收稿日期:2016-02-22
Effects of Enclosure on Soil Microbial Quantity and Enzyme Activity in Desert Steppe
LUO Dong1, WANG Mingjiu1*, ZHENG Shaolong2, LI Yuanheng3
1. College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Hohhot 010019, China;2. Bureau of Foresty, Naiman banner of Tongliao city in Inner Mongolia, Tongliao 028300, china;3. Institute of Grassland Research, Chinese Academy of Agricultural Sciences, Hohhot 010010, China;
Abstract:This study conducted on a Stipa breviflora dominated temperate desert steppe in Ordos plateaus, a comparative studied the impacts of grazing and enclosure on belowground standing biomass, soil organic matter, soil microbial quantity and enzyme activity. Objective of this study was to provide a scientific basis for the recovery and reconstruction of degraded grassland in arid areas. The key findings are, (1) Compared with the grazing area, enclosure significantly increased 23.81% and 37.21% grassland belowground standing biomass in the surface soil (0~10 cm) and in the subsurface soil (10~20 cm), respectively, while it had no significant effect on soil organic matter content. (2) Enclosure significantly improved 71% fungi population in soil surface and 147% bacterial population in subsurface, and also significantly increased 164% fungi population in underlying soil layer (20~40 cm) and 56% and 8.9% actinomycetes population in soil surface and in subsurface, respectively. (3) Enclosure significantly increased 31.13% urease activity in the subsurface soil, decreased 24.14% sucrose activity in the surface soil, and improved 16.59% catalase activity and 11.76% alkaline phosphatase activity in the surface soil. And (4) it was not consistent of the correlation between enclosure and grazing on different indexes, showing that the actinomycetes population was no significant correlation among different indexes under grazing treatment (P>0.05), while with the exception of no significant correlation between actinomycetes and fungi population with alkaline phosphatase activity under the enclosure (P>0.05), the other indexes showed significant (P<0.05) or very significant (P<0.01)positive correlation relationship with more than 68% correlation coefficient. In addition, the fungi population had a very significantly (P<0.01) positive correlation with four kinds of soil enzyme activity under the grazing condition, and the correlation coefficient is above 84%. And it was a significant positive correlation between the fungi population and four soil enzyme activity under the enclosure (P<0.05).
Key words:desert steppe; belowground biomass; soil organic matter; microorganism population; soil enzyme activity
