河北地区黄顶菊土壤种子库特征及其对替代控制的响应
2016-08-10张瑞海付卫东宋振晏静朱昌雄张国良中国农业科学院农业环境与可持续发展研究所北京100081
张瑞海,付卫东,宋振,晏静,朱昌雄,张国良中国农业科学院农业环境与可持续发展研究所,北京 100081
河北地区黄顶菊土壤种子库特征及其对替代控制的响应
张瑞海,付卫东,宋振,晏静,朱昌雄,张国良*
中国农业科学院农业环境与可持续发展研究所,北京 100081
摘要:土壤种子库在退化生态系统的恢复以及植被更新过程中发挥重要的作用。2001年黄顶菊(Flaveria bidentis)入侵河北地区,对当地生态系统造成了严重危害。为明确其潜在危害,调查了其在河堤与荒地两个典型生境中的土壤种子库特征。结果表明,(1)两种生境中地上植被均以黄顶菊最为丰富,频度达 100%。河堤及荒地黄顶菊种子总储量分别达 12 527和13 947 ind.·m-2,占整个种子库的88.05%及96.32%;黄顶菊种子库与植被间物种相似系数(S)分别为0.55(河堤)和0.40(荒地),属于中等及偏低相似水平。(2)黄顶菊种子多集中于上层土壤中(0~2 cm)中,且随土层深度的增加而减少,但土壤中层和下层(2~10 cm)中仍有近50%的黄顶菊种子。(3)在3次取样中,黄顶菊种子主要集中于4月份与6月份采集的土样中,8月份较少。本研究利用紫穗槐(Amorpha fruticosa)与沙打旺(Astragalus adsurgens)两种植物替代控制黄顶菊,分析了黄顶菊对替代控制后的响应,结果发现第2年控制效果优于第1年,其密度、单株结实量、土壤种子储量均极显著低于同期对照(P<0.01;)其次,其余生态指标如株高、分枝数、生物量等均受到不同程度的抑制,因此利用这两种替代植物可以限制黄顶菊的繁殖与扩散。该研究对黄顶菊的综合持续控制具有一定的实践指导意义。
关键词:黄顶菊;生物入侵;土壤种子库;时空特性;替代控制
引用格式:张瑞海, 付卫东, 宋振, 晏静, 朱昌雄, 张国良. 河北地区黄顶菊土壤种子库特征及其对替代控制的响应[J]. 生态环境学报, 2016, 25(5): 775-782.
ZHANG Ruihai, FU Weidong, SONG Zhen, YAN Jing, ZHU Changxiong, ZHANG Guoliang. Characteristics of Soil Seed Bank of Flaveria bidentis and Its Response to Control with Replacement Plants in Hebei, China [J]. Ecology and Environmental Sciences,2016, 25(5): 775-782.
土壤种子库(soil seed bank)是指存在于土壤上层凋落物和土壤中全部种子的总和(Roberts,1981),是植物潜在的种群,尤其是在受外来入侵植物危害的生态系统,土壤种子库的特征对生态系统的恢复和未来植被的结构、功能、组成与动态等更为重要(Gioria et al.,2012;2014)。根据种子在土壤中存活时间长短,土壤种子库可分为短暂土壤种子库(transient soil seed banks,存活时间小于1年)和持久土壤种子库(persistent soil seed banks,存活时间超过1年)(Thompson et al.,1979)。
近年来外来植物入侵已成为全球性生态问题,外来植物强大的生态适应性、可塑性使其成功入侵,入侵植物种子的萌发、种苗的补充以及群落的重组是其繁殖扩散的关键(Wagner et al.,2013)。土壤种子库的特征已成为入侵生态学的研究热点之一,大多数研究集中在其入侵机制(常瑞英等,2013;Fennell et al.,2014;Milakovic et al.,2016)、对群落结构的影响(唐樱殷等,2011;陈雯等,2012;Robertson et al.,2012)及综合防控(Alexander et al.,2003;李鸣光等,2012;)等方面,而对入侵植物采取防治措施后的土壤种子库的响应研究较少。
黄顶菊Flaveria bidentis (L.) Kuntze原产于南美洲,属一年生杂草,2001年在我国河北衡水湖及天津南开相继被发现,已在河北、天津、山东及河南等地区扩散蔓延,并形成优势群落,抑制其他植物的生长,造成生物多样性降低,严重破坏了入侵地的生态环境。在入侵农田后造成农作物减产和严重的经济损失(高贤明等,2004;刘全儒,2005;张国良等,2013119-125),成为亟待科学防控的外来入侵种之一。黄顶菊主要靠种子繁殖,繁殖量大,1株黄顶菊可产十几万粒种子(李香菊等,2007),且黄顶菊种子的寿命较长,室温下储藏的种子4年之后仍能萌发(李少青等,2010),这些特性使黄顶菊的防除和控制难度增加。因此,控制黄顶菊结实量、降低土壤中黄顶菊种子库被认为是有效控制黄顶菊的策略之一(Vitalos et al.,2009)。
目前对黄顶菊的研究侧重于其分布与危害、生长发育、活性物质提取、入侵机制及防控技术等方面(李香菊等,2007;任艳萍等,2008;张米茹等,2010;张瑞海,2010;张天瑞等,2011;Wei et al.,2011;纪巧凤等,2014;赵晓红等,2015),但对其土壤种子库的时空分布及其对替代控制措施种子繁衍的响应未见报道。本研究以此为切入点,对河堤与荒地两个典型入侵生境中的黄顶菊土壤种子库分布特征进行调查,并对替代控制措施后黄顶菊种子的生产能力及种子库进行了研究,旨在了解黄顶菊种子库土壤分布规律,明确替代控制措施对黄顶菊种子库的影响机制,以期为有效综合治理黄顶菊提供科学依据。
1 材料与方法
1.1 黄顶菊入侵生境特征调查
选取了两个黄顶菊发生的典型生境,生境一为河堤,位于河北冀州(37°34.593′N,115°21.347′E),海拔21 m,年平均气温12.7 ℃,年平均日照时数2571.2 h,年平均降水量510.3 mm,土壤类型为潮土。生境二为荒地,位于河北献县(38°13.800′N,115°57.578′E),海拔16 m,年平均气温12.3 ℃,年平均日照时数2800 h,年平均降水量560 mm,土壤类型为潮土。
在上述两个生境中,根据黄顶菊生长状况,采取对角线取样方法选取 30个样方(面积为 100 cm×100 cm),在样方内随机取10 cm×10 cm的小样方,分层取样,取样深度依次为0~2 cm(上层)、2~5 cm(中层)、5~10 cm(下层)。根据黄顶菊生长周期确定3次取样时间,分别为黄顶菊种子雨后且种子萌发前(2014年4月10日)、黄顶菊种子萌发中期(2014年6月20日)及黄顶菊种子成熟脱落前(2014年8月23日),其中在6月和8月两次取样中,记录样方内植物的种类及数量。
将取回的土样过 40目筛滤出杂物,然后均匀平铺于20 cm×15 cm(内径×高)的盆钵中,置于温室中,隔离外界种子干扰,定期浇水,逐日记录种子发芽数,对已萌发出的幼苗进行种类鉴定、计数后清除。如连续两周无种子萌发,再将土样搅拌混合,继续观察,直至连续两周不再有种子萌发。

表1 替代植物种植播种方法Table 1 Methods of sowing replacement plants
样方内植物群落计算Drude多度与频度(安树青等,1996),土壤种子库储量以密度来表达。本研究中,黄顶菊土壤种子库密度表达为每平方米单位面积上10 cm×10 cm的土壤中有活力的黄顶菊种子数量,以萌发的黄顶菊幼苗来计算,将取样面积的种子数目换算为1.0 m2,面积的数量即为种子库的种子密度。地上植被与种子库间物种的相似性系数采用Sørensen公式计算(Sørensen,1948):

其中,S为相似性系数,a为种子库中物种数,b为植被中物种数,c为a和b共有物种种数。
1.2 替代控制对黄顶菊结实量及种子库的影响
试验区设在河北省献县陌南镇撂荒地(无黄顶菊发生),实验地气候情况为上述荒地生境,土壤类型为潮土,pH偏弱碱性。通过前期预实验,选取了紫穗槐(Amorpha fruticosa)与沙打旺(Astragalus adsurgens)两种植物用于黄顶菊替代控制试验。紫穗槐幼苗购于河北保定博野万通苗木基地,株高40 cm左右,沙打旺种子购于北京鑫农丰农业技术研究所。所用试验荒地翻耕后,划定小区,试验设3个平行处理,分别为紫穗槐与黄顶菊混种、沙打旺与黄顶菊混种、黄顶菊单种(CK)。每个处理设置5个小区重复,每个小区面积为25 m2(5 m×5 m),具体播种方法见表1。整个试验过程中不施肥,不浇灌水(自然降水),不进行刈割处理。试验周期为2年,种植时间为2014年4月上旬,分别在第1年和第2年9月下旬测量黄顶菊株高、分枝数、生物量、单株结实量,并统计黄顶菊植株数量,计算黄顶菊密度,并在黄顶菊种子雨后(第1年和第2年11月底),统计各处理土壤中黄顶菊种子库大小。生物量测定方法:用烘箱在105 ℃杀青15 min后,在80 ℃烘干至恒重。在千分之一天平上称其干重。黄顶菊密度计算方法:在每个小区内,随机选择5个1 m2的样点,统计该样点内的黄顶菊植株数量,取平均值。
1.3 数据分析
本研究中两组数据采用t测验进行分析,多组数据采用One-Way ANOVA方差分析,所有数据统计与分析在Excel 2013和SPSS 19.0软件中完成。
2 结果与分析
2.1 黄顶菊入侵生境种子库特征
2.1.1 黄顶菊入侵生境植被组成
经调查,冀州河堤生境中有草本植物 16种,献县荒地生境中有草本植物 12种,两种生境中均以黄顶菊最为丰富,Drude多度极大,频度达100%。冀州河堤生境中除黄顶菊外其它较占优势的物种为苘麻(Abutilon theophrasti)和葎草(Humulus candens),频度分别达80%和50%;献县荒地生境中以牛筋草(Eleusine indica)和黄花蒿(Artemisia annua)为优势,频度分别达50%和46.7%(表2)。
2.1.2 黄顶菊种子库特征
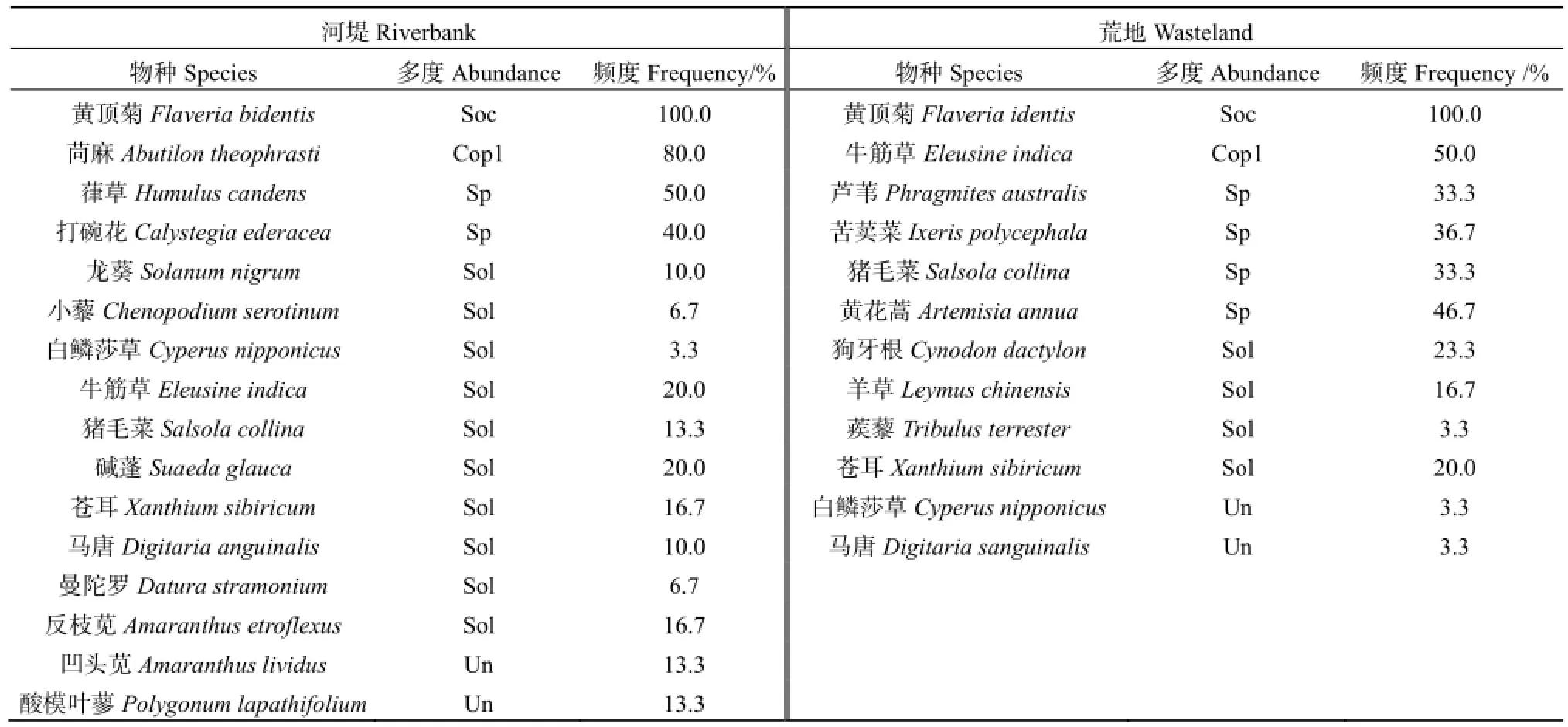
表2 调查区植被组成Table 2 Species composition of the vegetation investigated

表3 冀州河堤土壤种子库各物种组成及其所占储量比例Table 3 The species and proportion of plant of soil seed bank in riverbank in Jizhou
在两种生境的土壤种子库中共鉴定出 20种植物,隶属于10科18属。其中菊科与藜科植物为优势科,有5种生长在河堤生境中,4种生长在荒地生境中。河堤生境有17种植物,荒地样地有13种植物,两个样地的共有物种有9种。表3和表4分别列出了两种生境样地中各物种土壤种子库组成及其所占储量比例,从表中可以看出两种生境样地中黄顶菊种子量最多,分别占总储量的 88.05%及96.32%,成为优势群落,且黄顶菊种子库大小表现为荒地(13947 ind.·m-2)>河堤(12527 ind.·m-2)。除黄顶菊外,河堤生境种子库中碱蓬(Suaeda glauca)种子量最多,占总储量的 2.83%,而荒地生境中黄花蒿种子量最多,占总储量的1.22%。
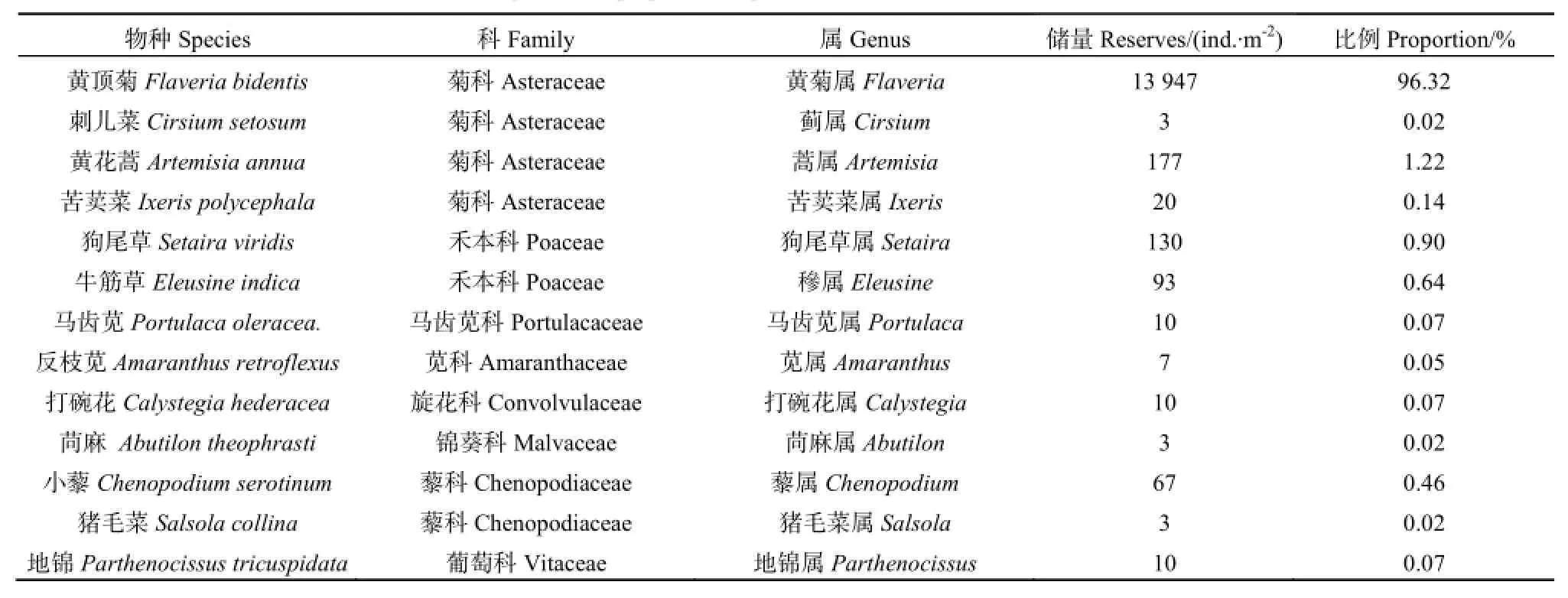
表4 献县荒地土壤种子库各物种组成及其所占储量比例Table 4 The species and proportion of plant of soil seed bank in wasteland in Xianxian
在垂直方向上,河堤与荒地两个生境中黄顶菊种子数量均随土壤深度的增加而减少(图1),在河堤生境3层土壤中黄顶菊种子量(n=30)平均分别为7150 ind.·m-2(0~2 cm)、3140 ind.·m-2(2~5 cm)、2337 ind.·m-2(5~10 cm);荒地生境3层土壤中黄顶菊种子数(n=30)平均分别为9747 ind.·m-2(0~2 cm)、2830 ind.·m-2(2~5 cm)、1370 ind.·m-2(5~10 cm)。两种生境中黄顶菊种子均主要分布在土壤上层(0~2 cm),分别占种子总量的51.87%及 69.88%。黄顶菊种子在5~10 cm深的土层内的存储量占总量的平均比例仍然较高,分别为 23.28%及9.82%。

图1 黄顶菊种子库垂直分布Fig. 1 Veritical distribution of the seed bank of F. bidentis in the soil
从图2可以看出,河堤与荒地两个生境中黄顶菊种子库大小随着时间的推移逐渐变小。在4月份采集的土样中黄顶菊种子最多,冀州河堤种子库大小平均为 6920 ind.·m-2,献县荒地平均为 7093 ind.·m-2;在6月份采集的土壤中黄顶菊种子量开始降低,分别为5273 ind.·m-2及6443 ind.·m-2,但差异不显著;在8月份采集时土壤种子库中仅有少量黄顶菊种子,显著低于前两次采样,种子库储量分别为433 ind.·m-2(河堤)和410 ind.·m-2(荒地)。
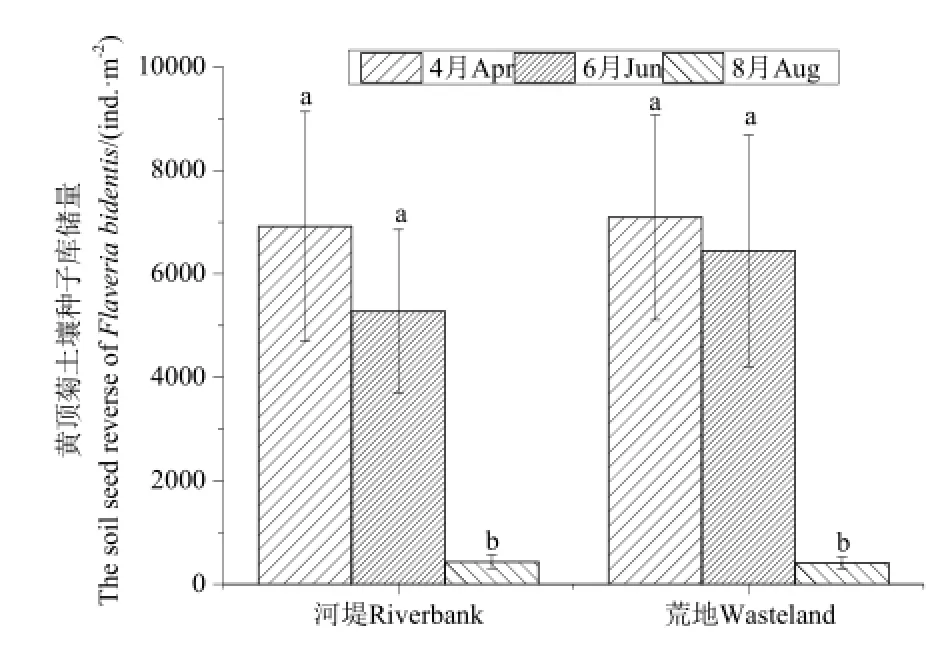
图2 黄顶菊种子库季节动态变化Fig. 2 The seasonal dynamics of the seed bank of F. bidentis in the soil
2.1.3 地上植被与种子库的关系
研究显示,河堤生境中黄顶菊种子库与植被间物种相似系数(S)为 0.55,属中等相似水平,地上植被与种子库中共有的物种有 10种,分别为黄顶菊、苘麻、打碗花(Calystegia ederacea)、小藜(Chenopodium serotinum)、牛津草、苍耳(Xanthium sibiricum)、马唐(Digitaria anguinalis)、曼陀罗(Datura stramonium)、凹头苋(Amaranthus lividus)和反枝苋(Amaranthus etroflexus)。荒地生境中黄顶菊种子库与植被物种相似系数(S)为 0.40,地上植被与种子库中共有的物种有4种,分别为黄顶菊、苦卖菜(Ixeris polycephala)、猪毛菜(Salsola collina)和黄花蒿,其中多数地上植被种类的种子并未在土壤种子库中出现。
2.2 替代控制措施下的黄顶菊种子库特征
2.2.1 替代控制处理对黄顶菊种子的影响
对采用两种替代植物进行替代控制的黄顶菊植株的密度展开调查,发现紫穗槐处理中的黄顶菊密度由第1年的18.5 plant·m-2下降到第2年的9.1 plant·m-2(表5),下降50.8%,与对照比差异极显著(P=0.003);沙打旺处理区中的黄顶菊密度由第 1年的 17.2 plant·m-2下降到第 2年的 0 plant·m-2。在替代控制后期,紫穗槐、沙打旺覆盖度大,与黄顶菊竞争光照、水分和养分,挤占了黄顶菊的资源生态位,使黄顶菊的生长及繁殖受到显著的限制。
在两种植物替代控制处理后,黄顶菊的结实量与土壤种子库总储量均有显著降低(表5)。在紫穗槐替代控制处理下,第1年黄顶菊种子产量已显著低于同期对照,到了第2年,仅为0.8万粒·株-1,与对照(18.2万粒·株-1)比差异极显著(P=0.000)。而沙打旺处理下黄顶菊单株结实量由10.2万粒·株-1下降至0万粒·株-1(截至试验结束,小区中未发现黄顶菊存活)。在紫穗槐替代控制处理下,黄顶菊土壤种子库由第1年的8250 ind.·m-2降到第2年的3831 ind.·m-2,与对照(14512 ind.·m-2)比差异极显著(P=0.003)。在沙打旺替代控制处理下,土壤种子库由第1年的6392 ind.·m-2降到第2年的1245 ind.·m-2,相比对照,差异极显著(P=0.000)。可见虽然试验结束时,无黄顶菊植株残留,但依然有大量的黄顶菊种子存留于土壤中。替代控制处理较好地控制了黄顶菊的生长结实,减少了土壤中黄顶菊种子库储量。
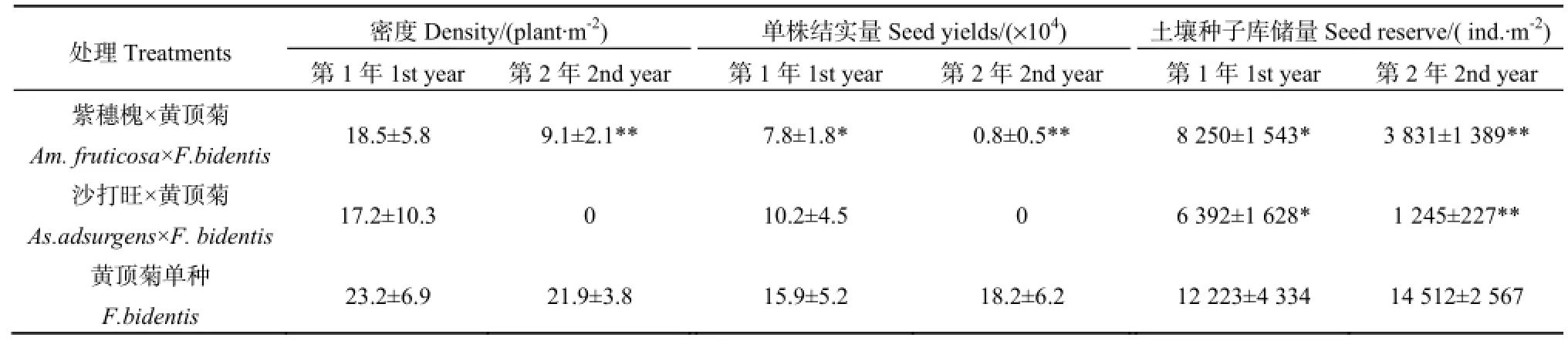
表5 替代控制对黄顶菊种子的影响Table 5 Effect of Am. fruticosa and Af. adsurgens growth on the destiny, numbers of seeds and seed bank of F. bidentis
2.2.2 替代控制处理对黄顶菊生态指标的影响
从其它指标方面可以看出,在替代控制处理第2年时紫穗槐处理下的黄顶菊株高(P=0.001)、分枝数(P=0.000)、生物量(P=0.000)均极显著低于对照(表6)。截至试验结束时,沙打旺与黄顶菊混种处理区并没有检测到黄顶菊植株及其种子,因此该处理区中黄顶菊各项指标均为0。

表6 紫穗槐和沙打旺对黄顶菊生态指标的影响Table 6 The impact of competition on Am. fruticosa and As.adsurgens against F.bidentis
3 讨论与结论
3.1 讨论
3.1.1 黄顶菊种子库特征
入侵植物的发生危害在很大程度上依赖于入侵植物土壤种子储量的大小(党伟光等,2008;张衍雷等,2015)。成熟的黄顶菊种子在落入土壤后的第1个萌发季节仍然保持有萌发活力,能够形成持久土壤种子库(Baskin et al.,1989)。通过调查河堤和荒地两个黄顶菊入侵的典型生境中的地上植被特征及土壤种子库特征,发现两种生境中群落组成比较简单,地上植被均以黄顶菊最为丰富,频度达100%,占总储量的88.05%(河堤)及96.32%(荒地)。因此,即使把地上黄顶菊种群全部灭除,土壤种子库中潜在的种群仍可形成再度入侵。
研究表明,大部分土壤种子库随土层加深,种子储量减少(彭军等,2000;Cao et al.,2010)。本研究结果体现了同样的规律,河堤和荒地两个生境中大于 50%的黄顶菊土壤种子分布在土壤上层(0~2 cm)。由于两个生境人为干扰少,黄顶菊种子雨后,大部分种子保留在上层,少数种子受风雨、畜牧、偶尔人为扰动等因素影响进入土壤中层与下层。处于土壤中下层的黄顶菊种子虽萌发率低,但可进入环境休眠状态,一旦条件合适即可萌发(张国良等,2013)170-171。因此,需要注意土壤中层与下层(2~10 cm)中黄顶菊种子的潜在危害。
在我国河北黄顶菊发生区,黄顶菊种子在4—6月均可萌发(张米茹,2009),本研究所得数据与之相符,大部分黄顶菊种子存在于4月份与6月份采集的土样中,8月份采集的土样中仍有少量黄顶菊种子萌发。黄顶菊这种萌发时间不一致、分批出苗的策略,延长了其在入侵地的发生时间,从而大大增加了防除难度(张国良等,2013)166-170。
地上植物种子雨是土壤种子库中种子的直接来源(于顺利等,2007),近年来对地上植物与物种组成及土壤种子库组成相似性有较多研究(盛丽等,2010;白文娟等,2012;刘庆艳等,2014),研究者发现在演替早期或受干扰程度较大的生境中,地上植被与土壤种子库间相似性高(张志权,1996)。本文中黄顶菊种子库与植被间物种相似系数(S)分别为0.55(河堤)和0.40(荒地),属于中等及偏低相似水平。出现该现象的原因可能是黄顶菊入侵后已适应当地环境,抑制其它伴生植物的生长,造成生态系统群落多样性降低;也可能与种子传播和种子繁殖策略有关,如芦苇(Xiong et al.,2001)、羊草(黄晓辉等,2013)等植物主要依靠根状茎进行营养繁殖,由于种子不具有活力或活力差,种子库中很难发现其种子。
本研究明确了黄顶菊典型生境中地上植被特征以及地下土壤种子库的结构与组成,并分析了二者直接的关系,可为科学预测黄顶菊的发生危害、种群扩张、防除提供科学指导。
3.1.2 替代控制措施对黄顶菊种子库的影响
Sheley et al.(2003)提出,对外来入侵物种的管理需要建立在生态学原则的基础上。除对原生态系统的保护措施外,替代控制措施是一种有效控制入侵植物的方法(Wan et al.,2010),且可以有效减少土壤中入侵植物种子量(王重云,2006),进而预防和控制入侵植物的发生与扩散。国内关于黄顶菊替代控制已有较多研究(皇甫超河等,2010;张瑞海等,2012),但这些研究中使用的替代植物多为一年生,很难对黄顶菊进行多年连续控制;且未对替代控制后的黄顶菊土壤种子库展开调查。本研究选用多年生植物紫穗槐与沙打旺作为替代植物,消除了短期控制的弊端。替代控制第1年,两种替代植物生长较黄顶菊缓慢,未能体现较好的控制效果,但第2年两种替代植物发芽早,生长迅速,很快成长为郁闭度较高的优势群体。而黄顶菊为典型的C4喜光植物(McKown et al.,2005),种子需要一定的光照刺激才能萌发(张米茹等,2010),替代植物的遮阴郁闭作用有效地阻止了次年黄顶菊种子的萌发(杨晴等,2014),而且萌发的黄顶菊也很难与长势高大、郁闭度高的紫穗槐、沙打旺争夺光照资源,黄顶菊植株生长受到较强抑制,其密度和结实量也随之显著降低,最终土壤中黄顶菊种子库储量也显著减少。即便是用沙打旺替代控制后黄顶菊植株消亡,但土壤中仍有大量黄顶菊种子,因此需要对黄顶菊进行长期有效的控制。选择具有一定经济价值的替代植物紫穗槐与沙打旺,可显著抑制黄顶菊的生长繁殖,减少土壤中的黄顶菊种子库储量,并且可实现对黄顶菊的多年持续控制,是一种非常有效的生态控制黄顶菊的方法,具有广泛应用价值。
3.2 结论
黄顶菊入侵河堤和荒地以后,群落组成逐渐单一化,两种生境中地上植被均以黄顶菊最为丰富,频度达 100%。土壤中黄顶菊种子库储量丰富,大部分黄顶菊种子存在于4月份与6月份采集的土样中,且大于50%的黄顶菊种子分布在土壤上层(0~2 cm),但仍需要注意土壤中层与下层(2~10 cm)中黄顶菊种子的潜在危害。此外本实验选用紫穗槐与沙打旺两种替代植物,紫穗槐是常见的护坡植物,沙打旺是常见的牧草,二者均不会引起新一轮的植物入侵。替代处理两年后,黄顶菊的株高、分枝数、生物量以及密度、结实量、进入土壤中种子储量均显著低于对照,因此利用这两种植物替代控制黄顶菊能够取得较好的控制效果,可用于控制公路两边、荒地、农田周边黄顶菊的发生与蔓延。本研究对黄顶菊的综合持续控制具有一定的指导意义。
参考文献:
ALEXANDER J M, D'ANTONIO C M. 2003. Seed bank dynamics of French broom in coastal California grasslands: Effects of stand age and prescribed burning on control and restoration [J]. Restoration Ecology,11(2): 185-197.
BASKIN J M, BASKIN C C, SPOONER D M. 1989. Role of temperature,light and date: seeds were exhumed from soil on germination of four wetland perennials [J]. Aquatic Botany, 35(3): 387-394.
CAO M, TANG Y, SHENG C, et al. 2000. Viable seeds buried in the tropical forest soils of Xishuangbanna, SW China [J]. Seed Science Research, 10(3): 255-264.
FENNELL M, GALLAGHER T, VINTRO L L, Osborne B A. 2014. Using soil seed banks to assess temporal patterns of genetic variation in invasive plant populations [J]. Ecology and Evolution, 4(9): 1648-1658.
GIORIA M, JAROŠÍK V, PYŠEK P. 2014. Impact of invasions by alien plants on soil seed bank communities: emerging patterns [J]. Perspectives in Plant Ecology, Evolution and Systematics, 16(3): 132-142.
GIORIA M, PYŠEK P, MORAVCOVA L. 2012. Soil seed banks in plant invasions: promoting species invasiveness and long-term impact on plant community dynamics [J]. Preslia, 84(2): 327-350.
MCKOWN A D, MONCALVO J M, DENGLER N G. 2005. Phylogeny of Flaveria (Asteraceae) and inference of C4photosynthesis evolution [J]. American Journal of Botany, 92(11): 1911-1928.
MILAKOVIC I, KARRER G. 2016. The influence of mowing regime on the soil seed bank of the invasive plant Ambrosia artemisiifolia L [J]. NeoBiota, 28: 39-49.
ROBERTS H A. 1981. Seed banks in soils [J]. Advances in Applied Biology, 6: 1-55.
ROBERTSON S G, HICKMAN K R. 2012. Aboveground plant community and seed bank composition along an invasion gradient [J]. Plant Ecology 213(9): 1461-1475.
SHELEY R L, KRUEGER-MANGOLD J. 2003. Principles for restoring invasive plant-infested rangeland [J]. Weed Science, 51(2): 260-265.
SØRENSEN T. 1948. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish common [J]. Biologiske Skrifter/Kongelige Danske Videnskabernes Selskab, 5: 1-34.
THOMPSON K, GRIME J. 1979. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats [J]. Journal of Ecology,67(3): 893-921
VITALOS M, MILAKOVIC I, KARRER G. 2009. Weed management by soil seed bank management-on the model of Ambroisia artemisiifolia L [C]//XIIIème Colloque International sur la Biologie des Mauvaises Herbes, Dijon, France, 8-10 Septembre 2009. Association Française de Protection des Plantes (AFPP): 496.
WAGNER, K, BOGUSCH, W, ZOTZ, G. 2013. The role of the regeneration niche for the vertical stratification of vascular epiphytes [J]. Journal of Tropical Ecology, 29(29): 277-290.
WAN F H, LIU W X, GUO J Y, et al. 2010. Invasive mechanism and control strategy of Ageratina adenophora (Sprengel) [J]. Science China Life Sciences, 53(11): 1291-1298.
WEI Y, XIE Q, FISHER D, et al. 2011. Separation of patuletin-3-O-glucoside, astragalin, quercetin, kaempferol and isorhamnetin from Flaveria bidentis (L.) Kuntze by elution-pump-out high-performance counter-current chromatography [J]. Journal of Chromatography A,1218(36): 6206-6211.
XIONG S, NILSSON C. 2001. The effects of plant litter on vegetation: a meta-analysis [J]. Journal of Ecology, 87(6): 984-994.
安树青, 林向阳, 洪必恭. 1996. 宝华山主要植被类型土壤种子库存初探[J]. 植物生态学报, 20(1): 41-50.
白文娟, 章家恩, 全国明. 2012. 土壤种子库研究的热点问题及发展趋向[J]. 土壤, 44(4): 562-569.
常瑞英, 王仁卿, 张依然, 等. 2013. 入侵植物空心莲子草的入侵机制及综合管理[J]. 生态与农村环境学报, 2013(1): 17-23.
陈雯, 李涛, 郑荣泉, 等. 2012. 加拿大一枝黄花入侵对土壤动物群落结构的影响[J]. 生态学报, 32(22): 7072-7081.
党伟光, 高贤明, 王瑾芳, 等. 2008. 紫茎泽兰入侵地区土壤种子库特征[J]. 生物多样性, 16(2): 126-132.
高贤明, 唐廷贵, 梁宇, 等. 2004. 外来植物黄顶菊的入侵警报及防控对策[J]. 生物多样性, 12(2): 274-279.
皇甫超河, 张天瑞, 刘红梅, 等. 2010. 三种牧草植物对黄顶菊田间替代控制[J]. 生态学杂志, 29(8):1511-1518.
黄晓辉, 胡小文, 徐宗海, 等. 2013. 羊草种子休眠和萌发的激素调控研究[J]. 草业学报, 22(5): 183-189.
纪巧凤, 宋振, 张国良, 等. 2014. 黄顶菊入侵对土壤磷细菌多样性的影响[J]. 农业资源与环境学报, 31(2): 175-181.
李鸣光, 鲁尔贝, 郭强, 等. 2012. 入侵种薇甘菊防治措施及策略评估.生态学报, 32(10): 3240-3251.
李少青, 倪汉文, 方宇, 等. 2010. 黄顶菊种子休眠与种子寿命研究[J].杂草科学, (2): 18-21.
李香菊, 王贵启, 张朝贤, 等. 2007. 外来植物黄顶菊的分布, 特征特性及化学防除[J]. 杂草科学, (4): 58-61.
刘庆艳, 姜 明, 吕宪国, 等. 2014. 湿地土壤种子库与地上植被相似性关系研究评述[J]. 生态学报, 34(24): 7465-7474.
刘全儒. 2005. 中国菊科植物一新归化属——黄菊属[J]. 植物分类学报,43(2): 178-180.
彭军, 董鸣. 2000. 重庆四面山亚热带常绿阔叶林种子库研究[J]. 植物生态学报, 24(2): 209-214.
任艳萍, 江莎, 古松, 等. 2008. 外来植物黄顶菊(Flaveria bidentis)的研究进展[J]. 热带亚热带植物学报, 16(4): 390-396.
盛丽, 王彦龙. 2010. 退化草地改建对土壤种子库及其与植被关系的影响[J]. 草业科学, 27(8): 39-43.
唐樱殷, 沈有信. 2011. 云南南部和中部地区公路旁紫茎泽兰土壤种子库分布格局[J]. 生态学报, 31(12): 3368-3375.
王重云. 2006. 紫茎泽兰迹地上不同替代植物群落结构和物种多样性的变化[D]. 西双版纳: 中国科学院西双版纳热带植物园.
杨晴, 李婧实, 郭艾英, 等. 2014. 遮阴和藜竞争对黄顶菊光合荧光和生长特性的影响[J]. 应用生态学报, 25(9): 2536-2542.
于顺利, 郎南军, 彭明俊, 等. 2007. 种子雨研究进展[J]. 生态学杂志,26(10): 1646-1652.
张国良, 付卫东, 郑浩. 2013. 黄顶菊入侵机制及综合治理[M]. 北京:科学出版社.
张米茹, 李香菊. 2010. 光对入侵性植物黄顶菊种子萌发及植株生长的影响[J]. 植物保护, 36(1): 99-102.
张米茹. 2009. 入侵性杂草黄顶菊生态学特性研究[D]. 北京: 中国农业科学院研究生院.
张瑞海, 付卫东, 张国良, 等. 2012. 紫花苜蓿和向日葵对黄顶菊的替代控制机理分析[J]. 西南大学学报(自然科学版), 34(2): 31-37.
张瑞海. 2010. 黄顶菊替代植物的筛选及其与黄顶菊竞争效应的研究[D]. 福州: 福建农林大学.
张天瑞, 皇甫超河, 杨殿林, 等. 2011. 外来植物黄顶菊的入侵机制及生态调控技术研究进展[J]. 草业学报, 20(3): 268-278.
张衍雷, 张瑞海, 付卫东, 等. 2015. 不同农作措施对少花蒺藜草(Cenchrus pauciflorus Benth)种子库及其繁殖能力的影响[J]. 农业资源与环境学报, 32(3): 312-320.
张志权. 1996. 土壤种子库[J]. 生态学杂志, 15(6): 36-42.
赵晓红, 杨殿林, 王慧, 等. 2015. 黄顶菊入侵对不同地区土壤氮循环及微生物量的影响[J]. 草业学报, 24(2): 62-69.
DOI:10.16258/j.cnki.1674-5906.2016.05.007
中图分类号:Q948
文献标志码:A
文章编号:1674-5906(2016)05-0775-08
基金项目:农业部公益性行业(农业)科研专项(200803022;201103027)
作者简介:张瑞海(1985年生),男,博士研究生,研究方向为入侵植物防控。E-mail: ruihai.321@163.com
*通信作者。E-mail: zhangguoliang@caas.cn
收稿日期:2016-03-30
Characteristics of Soil Seed Bank of Flaveria bidentis and Its Response to Control with Replacement Plants in Hebei, China
ZHANG Ruihai, FU Weidong, SONG Zhen, YAN Jing, ZHU Changxiong, ZHANG Guoliang*
Institute of Agricultural Environment and Sustainable Development, CAAS, Beijing 100081, China
Abstract:The soil seed bank plays an important role in the restoration of degraded ecosystems and vegetation renewal, Flaveria bidentis has caused serious damage to the native ecological system since its first occurrence in Hebei province of China in 2001. We surveyed the characteristics of soil seed bank of F. bidentis in two habitats riverbank and wasteland. The results showed that, (1) F. bidentis was the most abundant species in two habitats with its 100% frequency. The seed density of F. bidentis was 12 527 and 13 947 ind.·m-2, accounting for 88.05% and 96.32% of the soil seed bank in riverbank and wasteland respectively, the species similarity coefficient (s) between F. bidentis seed bank and vegetation was 0.55 (riverbank) and 0.40 (wasteland), belongs to the medium and lower similarity level, respectively. (2) The seeds of F. bidentis was decreased with the increasing of soil depth, seed density in the 0~2 cm soil layer was the highest, followed by the 2~5 cm layer while the 5~10 cm depth had the lowest seed density. (3) The seeds of F. bidentis can be mainly collected in April and June, fewer in August. In addition, we replaced F. bidentis by using Amorpha fruticosa and Astragalus adsurgens and investigeted its responses. The results indicated that effect of replacement control was better in the second year than the first year, the density, seed yield and soil seed reserve of F. bidentis were decreased significantly after two years (P < 0.01), and the other ecological indicators of F. bidentis such as the height, number of branches and biomass were inhibited by different degrees, thus it can limit the propagation and spread of F. bidentis by using these two kinds of substitutive plants. The results and findings in the present study can help in the sustainable management of F. bidentis.
Key words:Flaveria bidentis; biological invasion; soil seed bank; temporal and special dynamics; replacement control
