筇竹光合日变化特征研究
2016-08-09王林昊董文渊唐海龙张营顾西南林业大学环境与工程学院昆明6504西南林业大学云南生物多样性研究院昆明6504西南林业大学林学院昆明6504
王林昊董文渊唐海龙张 营顾 宝(西南林业大学环境与工程学院昆明6504 西南林业大学云南生物多样性研究院昆明6504 西南林业大学林学院昆明6504)
筇竹光合日变化特征研究
王林昊1董文渊2唐海龙3张 营3顾 宝3
(1西南林业大学环境与工程学院昆明650224 2西南林业大学云南生物多样性研究院昆明650224 3西南林业大学林学院昆明650224)
对筇竹春季、夏季和秋季的光合日变化特征进行了研究。结果表明:筇竹春季净光合速率日变化为单峰曲线,没有光合 “午休”现象;夏季和秋季净光合速率日变化为双峰曲线,有明显的光合 “午休”现象;筇竹光合 “午休”现象的主要原因是非气孔因素所致;夏季的净光合速率最高,光合能力最强,是筇竹枝叶生长旺季。影响筇竹不同季节净光合速率的最稳定环境因子为光合有效辐射和大气温度。筇竹夏季和秋季蒸腾速率变化趋势均表现为双峰曲线,春季为单峰曲线;夏季和秋季均有蒸腾 “午休”现象,春季则没有。光合有效辐射、大气温度和气孔导度是影响筇竹不同季节蒸腾速率的最稳定因子。
筇竹;光合特性;日变化;季节性
筇竹 (Qiongzhuea tumidinoda)是优良的笋材两用的中小型混生竹种,是国家3级保护珍稀濒危竹种,仅分布于金沙江下游两岸的局部区域,是西南地区特有的竹种。长期以来,研究者先后开展了筇竹种子特性及实生苗生长发育规律、筇竹无性系种群生物量结构和生态适应性、筇竹无性系地下茎生长规律以及筇竹群落的生物多样性研究等[1-2];但有关筇竹光合生理方面的研究未见报道,本文对筇竹不同季节光合日变化进行了初步研究,旨在揭示筇竹光合作用的变化规律。
1 试验材料与方法
1.1 试验地概况
试验地位于云南省昭通市大关县木杆镇银吉村徐家湾,海拔1 328~1 350 m,属中温带,年平均气温9~12℃;年平均降水量为1 000~1 300 mm,常年多雾多雨,空气湿度较大。试验地土壤主要以黄壤为主,母岩为玄武岩和紫色页岩,土层比较疏松并且湿润深厚,林下有较厚的腐殖层。
1.2 试验材料与研究方法
试验林为天然生长的筇竹林。因昭通市是云南省3个寡日照地区之一,特别是冬季日照时数少,且冬季筇竹林多为大雪覆盖,光合测定无法正常进行,因此选择春季、夏季和秋季进行光合测定。于2014年10月、2015年4月和8月各选择晴朗天气5 d,在8∶00—18∶00时间段内每隔2 h测定1次;测定时选取1年生生长健康的筇竹5株,每株取位于中部向南生长的成熟健康叶片3片。
采用ECA-PB0402光合测定仪进行光合测定,测定的光合指标有:气温 (T,℃),大气相对湿度(RH,%),光合有效辐射 [PAR,μmol/(m2· s)],净光合速率 [Pn,μmol CO2/(m2·s)],气孔导度 [Gs,mol H2O/(m2·s)],胞间CO2浓度 (Ci,μmol/mol),蒸腾速率 (Tr,mmol H2O/ (m2·s)]和水分利用率 (WUE,μmol CO2/mmol H2O)。测定的数据运用Microsoft Excel 2007分析,再利用 SPSS 17.0对其进行方差分析和相关性分析。
2 结果与分析
2.1 大气参数日变化
光合有效辐射 (PAR)日变化的分析结果见图1。由图1可知,筇竹春、夏、秋3个季节的PAR变化趋势均是单峰曲线,都是在12∶00左右达到峰值,峰值分别为 1036.38、1569.13和 1295.79 μmol/ (m2·s)。10∶00—14∶00时PAR相对较强,是筇竹进行光合作用适宜时期,10∶00之前与14∶00之后PAR相对较低,此时限制光合作用的主要因素是光照强度。
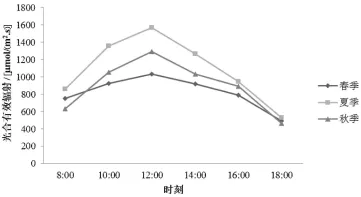
图1 光合有效辐射日变化特征
大气温度 (T)日变化的分析结果见图2。从图2看出,筇竹春、夏、秋3个季节的气温变化差异很大,变化大致范围分别是15.2~23.6℃、25.1~33.9℃和15.7~25.7℃。夏季温度在14∶00出现最大值,春季和秋季均是在12∶00出现最大值。中午的高温对光合作用 “午休”现象可能有一定的影响。
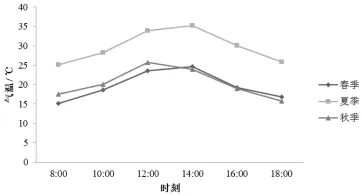
图2 气温日变化特征
相对湿度 (RH)日变化的分析结果见图3。可以看出,3个季节的RH均呈先下降后上升的趋势,不同的是夏季在14∶00下降到低谷,而春季和秋季在12∶00下降到最低值,然后开始上升。筇竹春、夏、秋3个季节的 RH变化范围分别为37.1% ~52.2%、47.3%~60.1%、38.1%~49.1%。
2.2 净光合速率日变化
从筇竹净光合速率 (Pn)的日变化特征来看(图4),春季Pn的日变化为单峰曲线,无光合 “午休”现象;夏季和秋季Pn的日变化均为双峰曲线,有明显的光合 “午休”现象;不同的是夏季筇竹在14∶00左右出现 “午休”现象,而秋季则是在12∶00左右出现 “午休”现象,且秋季的 “午休”现象没有夏季明显。
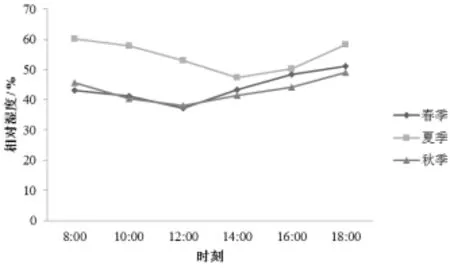
图3 相对湿度日变化特征
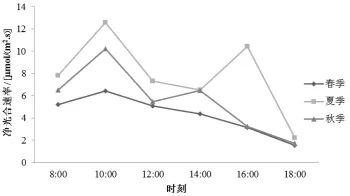
图4 筇竹净光合速率日变化特征
筇竹3个季节的Pn均是从8∶00开始上升,在10∶00左右达到峰值,随后春季Pn处于持续下降状态,夏季Pn下降后在16∶00左右上升到另一个峰值,秋季Pn则是在14∶00达到峰值。筇竹春季的PAR和T相较与夏季和秋季低,因此没有 “午休”现象。筇竹夏季和秋季的PAR均是在12∶00左右最大,但T在夏季14∶00达最高,在秋季12∶00为最高,这说明过高的气温是影响光合 “午休”现象的主要环境因素,是导致夏季和秋季光合 “午休”现象出现时间不同的原因。
2.3 气孔导度日变化
不同季节筇竹气孔导度 (Gs)的变化也不相同(图5),春季表现为单峰曲线,夏季和秋季为双峰曲线。3个季节Gs均是在10∶00达到峰值,随后春季Gs持续下降,夏季和秋季Gs分别在16∶00和14 ∶00达到另一峰值。Gs的变化主要与光照强度和温度变化紧密相关。早上日出后,光照强度不断增强,温度随之升高,气孔慢慢张开,因此Gs增加;但随着光照强度的增加,温度持续升高,为了减少水分蒸发,气孔关闭,Gs出现减小的情况;过了14∶00之后,光照强度下降,温度也随之降低,气孔张开程度又出现变化,Gs也随之变化;16∶00之后,由于光照强度和温度缓慢降低,气孔逐渐关闭,Gs处于持续下降过程。
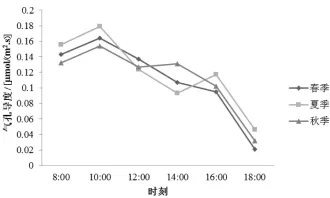
图5 筇竹气孔导度日变化特征
2.4 筇竹胞间CO2浓度日变化
植物胞间CO2浓度 (Ci)是外界CO2进入植物叶片细胞过程中所受到的不同驱动力、阻力以及叶肉细胞内光合作用与呼吸作用最终平衡的结果[3-4]。分析结果显示 (图6),筇竹3个季节的Ci均是从早上开始下降,并在10∶00降到低谷;春季在10∶00之后一直处于持续上升状态,而夏季和秋季则是处于一个曲折的上升与下降状态。夏季Ci在16∶00左右出现另一低谷,而秋季Ci的低谷则是在14∶00出现。出现这种现象的原因主要与Pn的变化有关,Pn变化快,细胞固定CO2能力就强,Ci就低;反之Pn变化慢,细胞固定CO2能力就弱,Ci就高。

图6 筇竹胞间CO2浓度日变化特征
2.5 蒸腾速率日变化
从筇竹蒸腾速率 (Tr)的测定结果看 (图7),3个季节的Tr变化与Pn的变化基本一致,夏季和秋季均是双峰曲线,春季是单峰曲线。3个季节Tr均是从8∶00开始增加,10∶00达到第1个峰值,分别为2.82、4.40和3.82 mmol/(m2·s);随后春季Tr一直下降,夏季Tr在16∶00达到第2个峰值,秋季Tr则在14∶00达到峰值。夏季和秋季均有蒸腾“午休”现象,春季没有表现出此现象。出现这种现象的原因是早上光照强度增强,温度也随之升高,气孔慢慢打开,植物体散失到空气中的水分随气孔张开程度的增加而增加,因此Tr是一个增长的趋势;随着光照强度的不断增加,温度也持续升高,植物为了减少水分的蒸发,气孔开闭程度慢慢减小,Tr出现降低现象。
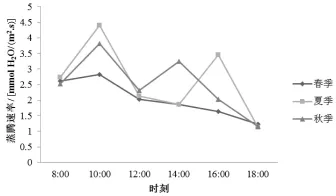
图7 筇竹蒸腾速率日变化特征
2.6 水分利用效率日变化
从筇竹水分利用效率 (WUE)的变化来看 (图8),3个季节的WUE变化趋势大致为单峰曲线,但每个季节在不同时段的变化有很大差异。筇竹夏季WUE明显高于春季和秋季,有研究表明,植物为了保持生长和水分的消耗平衡,在遇到相对缺水的环境时,WUE会发生相应的变化[5]。在木杆镇春季和秋季的降雨量远大于夏季,而且夏季的光照强度和温度较高,因此,夏季相较于春季和秋季比较缺水,所以WUE要高于春季和秋季。
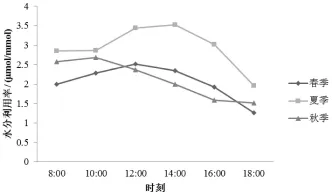
图8 筇竹水分利用率日变化特征
2.7 筇竹不同季节光合性状比较与方差分析
筇竹不同季节光合性状的测定结果与方差分析结果见表1。筇竹3个季节的Pn差异较大,夏季的Pn明显高于春季和秋季,即呈现夏季>秋季>春季的变化趋势。方差分析结果表明,不同季节间筇竹Pn差异显著,其中夏季与春季差异极显著,秋季与春季和夏季差异显著,说明不同季节筇竹对光能的利用效率存在明显差异,夏季的光合能力最强,是筇竹枝叶生长旺季。筇竹的Tr夏季最高,达到2.62 mmol H2O/(m2·s)以上,春季最小。筇竹不同季节的WUE从大到小的变化顺序为夏季>秋季>春季,表明筇竹在夏季有高效的水分利用率。影响植物WUE大小的因素不仅有植物本身的生物结构特征,如植物的根、茎、叶,而且受外界环境因子的影响也比较大[6],随着光照强度的逐渐增强,气温逐步上升,空气相对湿度下降,气孔张开程度发生变化,Pn与Tr也随之发生变化,导致WUE有所不同[7]。3个季节的Gs日均值均为0.060~0.165 mol H2O/(m2·s)。筇竹的Ci在春季处于一个较高水平,夏季和秋季基本处于同一水平。方差分析结果表明,不同季节间的Tr、Ci差异均不显著,夏季WUE与春季和秋季有显著差异,春季Gs与夏季和秋季相比差异显著。
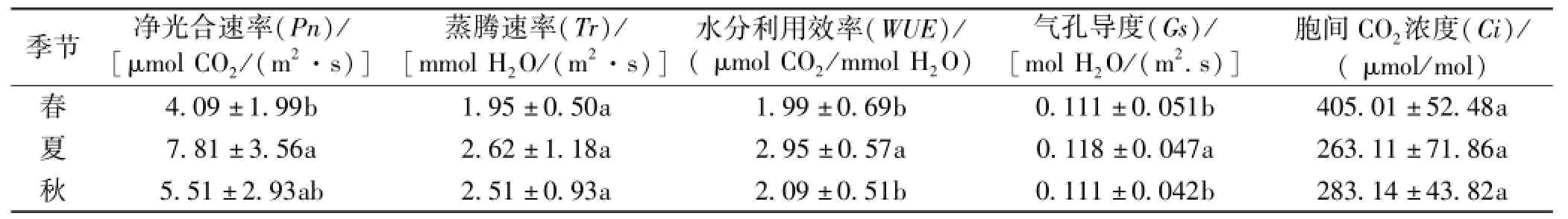
表1 不同季节光合性状测定值与差异性
2.8 不同季节筇竹净光合速率、蒸腾速率、水分利用效率与各环境因子的相关性分析筇竹光合性状与环境因子的相关性分析结果见表2。
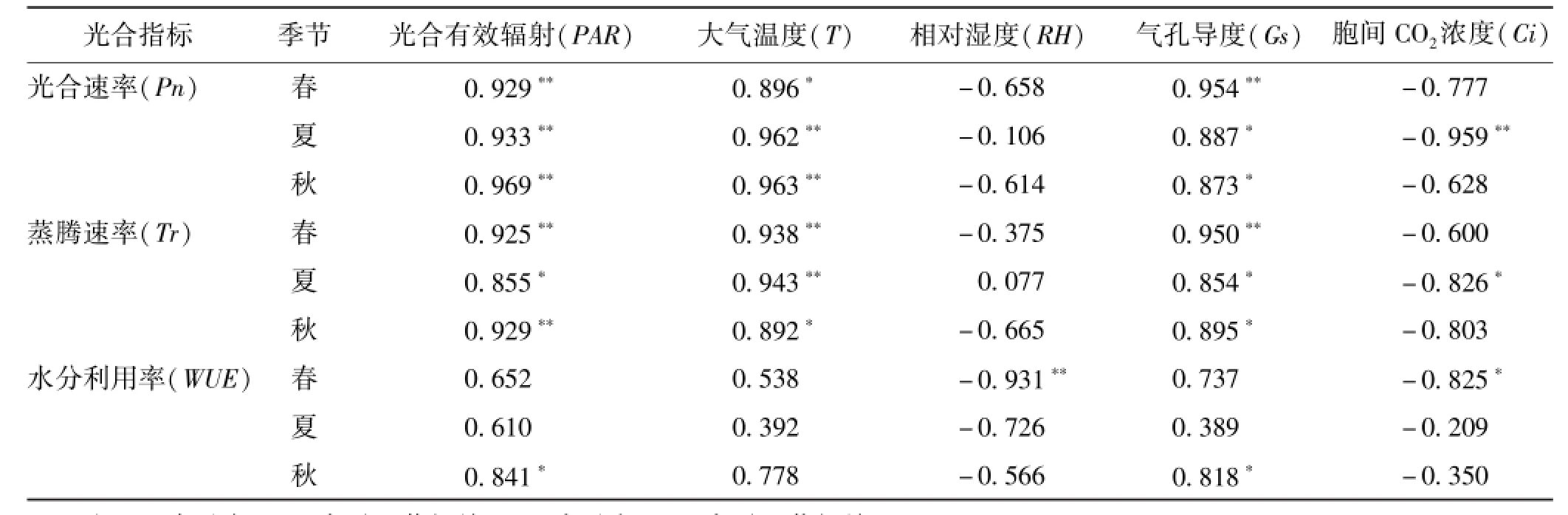
表2 不同季节Pn、Tr、WUE与各因子之间相关性分析
筇竹春季Pn与PAR和Gs有极显著正相关关系,与T呈显著正相关关系,与Ci的负相关性大于RH。筇竹夏季Pn与PAR、T和Ci有极显著相关关系,与Gs有显著正相关关系。筇竹秋季Pn与PAR 和T有极显著正相关关系,与Gs有显著正相关关系。结果表明,影响筇竹不同季节Pn最主要的环境因子是PAR和T;除此之外,春季受到Gs的影响也比较大,其次是Ci和RH;影响夏季Pn最明显的生理因子是Ci,秋季则是Gs。这表明筇竹不同季节Pn的不同,不仅仅是因为不同季节的环境因子不同所导致,还与自身的生理状况有关,环境因子与生理因子共同决定着筇竹不同季节Pn的变化。
筇竹春季Tr与PAR、T和Gs有极显著正相关关系;夏季Tr与T有极显著正相关性,PAR和Gs与其有显著的正相关关系,Ci与其呈显著负相关关系;而秋季Tr与PAR有极显著正相关性,与T和Gs有显著正相关关系。影响筇竹不同季节Tr变化的主要因子有PAR、T和Gs,但相同因子在不同季节的影响程度也是不同的,春季Tr主要是受Gs的影响,夏季则主要受T的影响,PAR对秋季的Tr影响比较大。此外,筇竹夏季Tr与Ci的相关性高于春季和秋季,这可能是夏季Tr大于春季和秋季的一个原因。
筇竹春季WUE与RH有极显著负相关关系,与Ci有显著负相关性。夏季WUE与RH的相关性较高,而秋季WUE则是与PAR和Gs有显著正相关关系。这说明影响筇竹不同季节WUE变化的因子各不相同,春季WUE主要受RH和Ci的影响;夏季WUE的影响因子主要是RH,其次是PAR;而PAR 和Gs则是影响秋季WUE变化的主要因素。
3 结论与讨论
植物光合作用效率与其生长环境有密切关系,植物往往通过改变其代谢来适应其生长环境,通过长期进化形成与环境相适应的光合特性,以此保证其正常的生长、发育以及繁殖[8-10]。日变化曲线类型的不同是植物光合生理活性与其环境因素的综合表现。不同季节筇竹光合作用日变化趋势和峰值出现的不同,主要是由环境条件引起气孔开闭水平的不同以及受到植物本身生理活性的影响导致[11-13]。
筇竹春季净光合速率变化为单峰曲线,没有光合 “午休”现象;筇竹夏季和秋季净光合速率均为双峰曲线,有明显的光合 “午休”现象。关于 “午休”现象有研究者认为,胞间CO2浓度高低是判断光合现象不可缺少的条件,即当净光合速率和气孔导度同时减小,胞间CO2浓度增加,说明光合抑制现象是由非气孔因素决定的;当净光合速率和气胞间CO2浓度同时减小,气孔导度增加,此时光合抑制现象是由气孔因素决定的[11]。据此判断,夏季和秋季上午净光合速率达到峰值后开始下降,气孔导度也是随之下降,而胞间CO2浓度处于上升状态,这表明光合 “午休”现象不是由气孔因素所致,而是由非气孔因素所引起的。不同季节筇竹对光能的利用效率存在明显差异,夏季光合能力最强,是筇竹枝叶生长的旺季。
影响筇竹不同季节净光合速率最主要的环境因子是光合有效辐射和大气温度;此外,春季影响净光合速率的主要因子为气孔导度,其次是胞间CO2浓度和相对湿度;夏季影响净光合速率最明显的生理因子是胞间CO2浓度,秋季则是气孔导度。
蒸腾速率的变化代表着植物水分运输强弱的不同。大部分植物净光合速率与蒸腾速率呈正相关关系,这主要是因为光合产物的生成需要水分以及不同矿物质营养的不断供应[12-15]。筇竹3个季节的蒸腾速率变化与净光合速率的变化基本一致。影响筇竹不同季节蒸腾速率的主要因子有光合有效辐射、大气温度和气孔导度,但在不同季节相同因子的影响程度是不同的。春季蒸腾速率主要是受气孔导度的影响,夏季则是受大气温度的影响,光合有效辐射对秋季的蒸腾速率影响比较大。筇竹夏季蒸腾速率与胞间CO2浓度的相关性高于春季和秋季。
植物水分利用效率不同主要取决于蒸腾速率与净光合速率的变化,主要受植物不同结构组织特征的影响,但气温、叶温、光强、湿度、气孔导度以及土壤水分等环境因子在一定程度上也影响着植物的水分利用效率[16-19]。筇竹3个季节的水分利用效率变化趋势大致为单峰曲线。筇竹夏季水分利用效率明显高于春季和秋季。影响水分利用效率大小的因素不仅有植物本身的生物结构特征,而外界环境因素对其也有一定的影响:春季主要受相对湿度和胞间CO2浓度的影响,夏季主要受相对湿度、光合有效辐射的影响,秋季主要受光合有效辐射和气孔导度的影响。
对不同季节筇竹光合日变化的研究初步揭示了筇竹光合生理特性,要更全面系统地了解筇竹光合生理特性,尚需开展筇竹光合和叶绿素年动态研究以及叶片整个生活期光合动态等研究。
[1] 董文渊.筇竹无性系地下茎生长规律的研究[J].竹子研究汇刊,2002,21(4):56-60.
[2] 董文渊,黄宝龙,谢泽轩,等.筇竹生长发育规律的研究[J].南京林业大学学报(自然科学版),2004,26 (3):43-47.
[3] 刘娟,马媛,廖康,等.新疆主栽杏品种的光响应曲线[J].经济林研究,2012,30(1):45-50.
[4] 迟丽华,宋凤斌.松嫩平原4种植物光合响应特性的研究[[J].吉林农业大学学报,2007,29(2):119-122,138.
[5] 马阔东,高丽,闫志坚,等.库布齐沙地三种植物光合蒸腾特性和水分利用效率研究[J].中国草地学报,2010,32(2):116-120.
[6] 喻方圆,徐锡增.水分和热胁迫处理对4种针叶树苗木气体交换和水分利用效率的影响[J].林业科学,2004,40(2):38-44.
[7] 山仑.水分利用效率[M]//邹承鲁.当代生物学.北京:中国致公出版社,2000:399-400.
[8] 周文杰,卢站根,魏淑珍.曼地亚红豆杉蒸腾速率日变化及因子分析[J].植物研究,2004,24(4):425-427.
[9] 黄启民,杨迪蝶,高爱新,等.不同条件下毛竹光合作用的研究[J].竹类研究,1989,8(2):8-16.
[10] 王俊刚,宋新青.不同竹龄雷竹若干光合特性的比较研究[J].浙江林业科技,2002,22(1):11-13.
[11] 黄勇.绿竹种源的光合特性研究[J].福建林业科技,2003,30(3):50-53.
[12] 郑炳松,金爱武,程晓建,等.雷竹光合特性的研究[J].福建林学院学报,2001,21(4):359-362.
[13] 陈海玲,吴立伟,石凤翎,等.苜蓿雄性不育系杂交组合光合特性及杂种优势分析[J].种子,2013,32(3):65-69.
[14] 黄春燕,吴卫,郑有良,等.鱼腥草光合蒸腾特性及影响因素的分析[J].西北植物学报,2006,26(5):989-994.
[15] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[16] 李秧秧.不同水分利用效率的高羊茅水分和光合特性研究[J].草业科学,1998,15(1):14-17,26.
[17] 刘庚山,郭安红,任三学,等.不同覆盖对夏玉米叶片光合和水分利用效率日变化的影响[J].水土保持学报,2004,18(2):152-155.
[18] 邓雄,李小明,张希明,等.四种荒漠植物的光合响应[J].生态学报,2003,23(3):598-605.
[19] 陶磊,严俊霞,陈良富,等.太原盆地玉米和大豆光合作用日变化和光合光响应[J].安徽农业科学,2011,39(5):2521-2523.
Study on Diurnal Variation of Photosynthetic Characteristics of Qiongzhuea tumidinoda
Wang Linhao1Dong Wenyuan2Tang Hailong3Zhang Ying3Gu Bao3
(1 College of Environment Science and Engineering,Southwest Forestry University,Kunming 650224,Yunnan,China 2 Research Institute of Biodiversity,Southwest Forestry University,Kunming 650224,Yunnan,China 3 College of Forestry,Southwest Forestry University,Kunming 650224,Yunnan,China)
We studied the diurnal variation of photosynthetic characteristics of Qiongzhuea tumidinoda in spring,summer and autumn.The results showed that the net photosynthetic rate in spring was showed as the“single peak”curve with no“midday depression”phenomenon while the rate in autumn and summer was showed as“double peak”curves with obvious“midday depression”phenomenon.Non-stomatal factor was the main cause of“midday depression”phenomenon.The net photosynthetic rate and the photosynthesis ability were the strongest in summer,in which Q.tumidity grows vibrantly.Photosynthetically active radiation(PAR)and air temperature(T)are the most stable environmental factors affecting net photosynthetic rate in different seasons.The transpiration rates(Tr)of Q.tumidity in summer and autumn all showed the double peak curve,but single peak curve in spring;transpiration “lunch break”phenomenon appeared in summer and spring but not in spring.PAR,and stomatal conductance(Gs)are the most stable factors to transpiration rate in different seasons.
Qiongzhuea tumidinosa,photosynthetic characteristics,diurnal variation,seasonality
10.13640/j.cnki.wbr.2016.02.002
国家林业公益性行业科研专项经费项目 (编号:201204013)。
王林昊 (1991-),男,硕士生,研究方向为竹林生态。E-mail:929306008j@qq.com。
董文渊 (1962-),男,教授,博士生导师,研究方向为竹类无性系种群生态、竹林培育和生态经济。E-mail:wydong6839@ sina.com。
