基于形态和16S rRNA基因的拟相手蟹分类研究
2016-08-09杨明柳徐敬明吴斌阎冰
杨明柳,徐敬明,吴斌,阎冰
(1. 广西科学院 广西红树林研究中心 广西红树林保护与利用重点实验室,广西 北海536000;2. 重庆文理学院 林学与生命科学学院,重庆 永川402168)
基于形态和16S rRNA基因的拟相手蟹分类研究
杨明柳1,徐敬明2*,吴斌1,阎冰1
(1. 广西科学院 广西红树林研究中心 广西红树林保护与利用重点实验室,广西 北海536000;2. 重庆文理学院 林学与生命科学学院,重庆 永川402168)
摘要:拟相手蟹属因其形态极其相似成为相手蟹科分类中最有疑问的一个属。通过对中国沿海近亲拟相手蟹Parasesarma affine、斑点拟相手蟹P. pictum、三栉拟相手蟹P. tripectinis、P. ungulatum及褶痕拟相手蟹P. plicatum 5种拟相手蟹的形态对比发现,可从它们的螯足形状(包括螯足掌节背面的梳状栉,可动指背面突起数目)及雄性第一腹肢形状对其进行区分。对其线粒体16S rRNA基因序列进行分子系统发育分析,结果表明5种拟相手蟹之间的遗传距离为1.9%~9.3%,达到了种间差异水平;构建的系统发育树显示近亲拟相手蟹与褶痕拟相手蟹汇聚成一支,随后与P. ungulatum聚在一起,斑点拟相手蟹与三栉拟相手蟹汇聚成独立支系。形态和分子证据均支持5种拟相手蟹分别为独立有效物种。
关键词:拟相手蟹;形态特征;16S rRNA基因;分类研究
1引言
拟相手蟹属(Parasesarma)隶属于方蟹总科(Grapsoidea),相手蟹科(Sesarmidae),全世界共记录34种[1—3]。有关拟相手蟹属的分类学研究,国内外学者虽进行了大量的研究[4—6],但因早期不同学者对一些新种、新记录的描述不够详细,或者一些模式标本的缺失,致使出现同物异名、异物同名等分类混乱局面,加上对一些种类了解甚少,因此,拟相手蟹属成为相手蟹科中最难分类的属之一。
中国记录的拟相手蟹有4种,分别是近亲拟相手蟹Parasesarmaaffine、斑点拟相手蟹P.pictum、三栉拟相手蟹P.tripectinis、精巧拟相手蟹P.exquisitum[7]。这4个种在形态上非常相似,均表现为头胸甲近方形,胃、心分区明显,额弯向下方,额后部具明显的4叶;外眼窝角呈三角形,指向前方,侧缘无齿;雄性螯足比雌性的大,掌节厚而短,背面具2~3列梳状栉;步足扁平等特征,很容易将其混淆。
国外已有研究表明广布于中国南部沿海的褶痕拟相手蟹P.plicatum实为近亲拟相手蟹P.affine,而中国目前尚未发现褶痕拟相手蟹P.plicatum,故在《中国海洋生物名录》中将其更正为近亲拟相手蟹P.affine[1,7—8]。然而诸多学者在红树林湿地蟹类相关研究中,褶痕拟相手蟹P.plicatum仍被沿用至今。Rahayu 和Ng[8]在检查南海(海南)标本时,发现存在P.ungulatum,由于这个种外形与褶痕拟相手蟹极其相似,其雄性螯足掌节背面具2列梳状栉,可动指背缘的突起少于15个,亦被误认为是褶痕拟相手蟹的同物异名种,但其可动指背缘的突起较褶痕拟相手蟹的小,且其雄性第一腹肢与褶痕拟相手蟹的相差很大,因此这些特征的差异表明P.ungulatum并非是褶痕拟相手蟹的同物异名种。精巧拟相手蟹、三栉拟相手蟹分别是戴爱云和宋玉枝[9]、Shen[10]发表的新种,自被报道以来,相关的研究还非常少,对它们的分布范围也缺乏相关资料,亦鲜有将多种拟相手蟹的形态区别进行综合对比描述。
作者对2012年7月至2013年6月于中国南部沿海红树林湿地及2015年6月广西北海冠头岭所采集的几种拟相手蟹进行了分类鉴定,通过对其形态特征的综合分析与比较,获得了从形态上区分它们的方法,并运用线粒体DNA的16S rRNA基因片段对其进行序列分析以及构建系统发育树,探讨其分子系统发育关系,以期利用形态学与分子生物学相结合的手段,解决我国拟相手蟹属的分类混乱局面,为今后的蟹类分类提供基础资料。
2材料与方法
2.1材料
几种拟相手蟹于2012年7月至2013年6月在中国南部沿海红树林湿地及2015年6月广西北海冠头岭所采集(表1),所有样品均浸泡于95%酒精中固定,并保存于-20℃冰箱,所有样品均保存于广西红树林研究中心生物室。
2.2方法
2.2.1形态描述分析
对拟相手蟹的头胸甲、腹部、螯足、步足、雄性第一腹肢等形态特征进行观察,拍照,描述。
2.2.2分子生物学分析
取约10 mg步足肌肉用基因组DNA提取试剂盒(OMEGA公司)进行提取,提取的DNA保存于-20℃。褶痕拟相手蟹及外群弧边招潮的序列则从GenBank里下载(表1)。

表1 拟相手蟹及外群弧边招潮16S rRNA基因片段信息
用于扩增16S rRNA目的基因片段的两个引物分别为:16Sar 5′-CGCCTGTTTAACAA AAACAT-3′, 16Sbr 5′-CCGGTCTGAACTCAGATCACGT-3′[13]。PCR扩增反应体积为50 μL,其中Taq10 2× MasterMix (北京奥赛博公司)19 μL,模板 DNA 2 μL,引物各 2 μL,Milli-Q-Water 补足到50 μL。反应条件为94℃预变性1.5 min后,94℃变性30 s,49℃退火45 , 72℃延伸1 min, 39个循环,最终72℃延伸5 min。产物经1%琼脂糖凝胶电泳,溴化乙锭(EB)染色检测之后,将样品送至广州英潍捷基测序公司进行纯化,并用ABI 3730XL测序仪进行正反链双向测序。
序列由DNAstar软件包的SeqMan、MegAlign和EditSeq程序进行编辑、校对和排序,并加以手工校正;MegAlign对比序列;DNAsp5.0检测多态位点数、简约信息位点数。首先对5种拟相手蟹的序列通过MrModeltest获取最佳替代模型HKY+I,再用Mega4.0选择其近似模型Tamura-Nei计算种内和种间遗传距离。在Mega4.0软件中采用Kimura-双参数距离模型构建NJ树,系统发育树各结点的支持率以序列数据集1 000次重复抽样检验的自引导值表示。MrBayes3.1构建贝叶斯树( Bayesian tree),采用PAUP软件[14]作同质性检验,通过MrModeltest的AIC准则(Akaike information criterion)获取最佳替代模型GTR+G,用MrBayes软件运行 Markov chain Montecarlo (MCMC) 运算1 000 000 代,由一个随机的树开始运算 4 条平行的链,每 100代产生 1 个树,得到贝叶斯树。
3结果与分析
本研究利用形态分类学和分子生物学相结合的方法对所采集的拟相手蟹进行了系统的分类学研究,共鉴定出4种拟相手蟹,分别为近亲拟相手蟹P.affine(737只)、斑点拟相手蟹P.pictum(6只)、三栉拟相手蟹P.tripectinis(2只)和P.ungulatum(9只)。
3.1物种形态描述
方蟹总科Superfamily Grapsoidea MacLeay, 1838
相手蟹科Family Sesarmidae Dana, 1851
拟相手蟹属GenusParasesarmaDe Man,1895
3.1.1近亲拟相手蟹ParasesarmaaffineDe Haan,1837
Grapsus(Pachysoma)affinisDe Haan, 1837: 66, pl. 18, fig. 5.
Sesarmaaffinis— H. Milne Edwards 1853: 183. -Heller 1862: 68.
Sesarmaquadratavar.affinis— Ortmann 1894: 724.
Parasesarmaplicatum— Miyake1983: 239.
Sesarma(Pachysoma)plicatum— Yamaguchi & Baba 1993: 481 (lectotype designated).
Sesarma(Parasesarma)plicata— Crosnier 1965: 65, figs. 83, 88, 93, 105. - Dai et al. 1986: 490, pl. 69 (2), fig. 275 (2). - Dai & Yang 1991: 537, pl. 69 (2), fig. 275 (2). - Shen 1932: 191, text fig. 119-120, pl. 7, fig. 8. -Shen & Dai 1964: 135. -Sakai 1939: 683, pl. 110, fig. 2.
Parasesarmaplicatum— Wang & Liu 1996: 117, figs. 150, 151. -Ng et al. 2001: 43.
Parasesarmaaffinis— Liu. 2008: 802.
Parasesarmaaffine— Ng et al. 2008: 222. - Rahayu & Ng 2010:1-22, figs, 5-7.
样品采集:东寨港2只(1♀,1♂);深圳福田3只(1♀,2♂);珠海39只(23♀,16♂);北海山口12只(4♀,8♂);北海草头村38只(17♀,21♂);防城港珍珠湾46只(22♀,24♂);雷州土角村82只(28♀,54♂);阳江32只(10♀,22♂);云霄竹塔村23只(10♀,13♂);龙海金定村34只(15♀,19♂);雷州企水海角村56只(24♀,32♂);海南东方9只(5♀,4♂);海南洋浦106只(40♀,66♂);大冠沙237只(84♀,153♂);徐闻仕尾18只(2♀,16♂)。
形态描述:头胸甲呈方形,宽度大于长度,宽度约当长度的1.35倍,表面稍隆,前半部及鳃区具粗糙颗粒及斜行颗粒隆线。额弯向下方,中部稍凹,额后部的4叶突出。外眼窝角锐三角形,指向前方,侧缘在外眼窝角之后有一个不明显的齿痕。雄性螯足粗壮等大,长节外侧缘具颗粒隆脊,近末端具刺,内侧缘近末端具一枚三角型刺,外侧面有横纹;掌节厚而短,表面具颗粒,背面具2梳状栉,分别具17~23及9~13个梳状齿;可动指背面具8~9个较大的突起,内缘具大小不等的钝齿。步足宽大扁平,第二、三对步足等长,较其他两对步足长,其长度约当头胸甲宽度的1.5倍;第三对步足长节长约当宽的2倍。雄性腹部宽三角形,尾节较短而宽,末缘半圆形,长度与第六节长度相当;第六节宽度约当长度的2倍多。雄性第一腹肢细长,末端趋长,弯曲呈45°,几丁质突起长而直,指向外方。雌腹部圆大,尾节宽大[4,8](图1,图2)。
分布:广西,广东,海南,浙江,福建,台湾,香港,日本,印度-西太平洋。

图1 近亲拟相手蟹(成体♂,头胸甲宽22.14 mm×长16.49 mm):A. 整体背面观;B. 正面观;C. 腹部;D. 右螯;E. 右螯可动指正面观;F. 螯足掌节背面梳状栉Fig.1 P. affine(adult male, 22.14 mm×16.49 mm): A. overall dorsal views; B. frontal view of carapace; C. ventral view; D. right cheliped; E. dorsal surface of dactylus of right cheliped showing dactylar tubercles; F. dorsal surface of chela showing diposition of pectinate crest

图2 近亲拟相手蟹雄性第一腹肢:A~D为右雄性第一腹肢整体观及末端放大;比例尺=1 mm(仿Rahayu 和Ng[8])Fig. 2 The male first gonopod of P. affine : A-D. right male first gonopod and enlargement of tip of gonopod one; scal=1 mm (modified from Rahayu and Ng[8])
3.1.2斑点拟相手蟹ParasesarmapictumDe Haan, 1835
Grapsus(Pachysoma)pictusDe Haan 1835:61, pl. 16, fig. 6.
Sesarmapicta— Krauss 1843: 45 - Alcock 1900: 414. - Stimpson 1907: 135.
Sesarma(Parasesarma)picta— De Man 1895: 183. - Tesch 1917: 186. - Balss 1922: 156. - Shen 1932:186, text-figs. 117,118, pl. 7, fig.7. - Sakai 1939: 682, pl. 78, fig. 2; 1976: 656, pl 226, fig. 2. - Shen & Dai 1964:135. - Dai et al. 1986: 489, 490, pl. 69(1), fig. 275(1).
Parasesarmapictum— Ng et al. 2008: 222. - Liu. 2008: 802.
样品采集:汕头1只(1♀);北海冠头岭5只(1♀,4♂)。
形态描述:头胸甲呈方形,宽度约当长度的1.2倍,表面扁平,前半部具短的横行颗粒隆线,胃、心区处具“H”形沟,鳃区具斜行隆线。额部弯向下方,中部凹入,额后部4叶明显。眼窝深,背缘光滑,腹缘内齿三角形。外眼窝角呈锐三角形,指向前方。侧缘呈光滑的隆线形,几乎平行,无齿。螯足雄比雌大,掌节厚而短,内、外侧面均具颗粒,背面具1~2列梳状栉和数条斜行颗粒隆线,两列梳状栉分别具13~15及10~11个梳状齿,其中第二列梳状齿较第一列低平,圆大,或呈颗粒状;螯足可动指背面具1列约13~20个卵圆形突起,雌螯约10个颗粒状突起,近末端的颗粒状突起不易区分,两指内缘具大小不等的钝齿。步足细长,以第3对步足最长,其长节长约当宽的2.4倍。雄性第一腹肢粗短,末端趋尖,弯曲呈60°,几丁质突起短,顶端呈截状斜面,弯向背外方。腹部宽三角形,尾节末缘圆钝,长于第6节;第6节宽度约当长度的2.8倍。雌性腹部圆大,尾节基部嵌入第六节中[4](图3,图4)。

图3 斑点拟相手蟹(成体♂,头胸甲宽17.75 mm×长14.64 mm):A. 整体背面观;B. 正面观;C. 腹部;D. 左螯;E. 左螯可动指正面观;F. 螯足掌节背面梳状栉Fig.3 P. pictum(adult male , 17.75 mm×14.64 mm): A. overall dorsal views; B. frontal view of carapace; C. ventral view; D. left cheliped; E. dorsal surface of dactylus of left cheliped showing dactylar tubercles; F. dorsal surface of chela showing diposition of pectinate crest

图4 斑点拟相手蟹雄性第一腹肢:A~C. 为右雄性第一腹肢整体观及末端放大;比例尺为1 mm(仿戴爱云等[4])Fig. 4 The male first gonopod of P. pictum: A-C. right male first gonopod and enlargement of tip of gonopod one; scal=1 mm (modified from Dai Aiyun et al.[4])
分布:广西,广东,福建,浙江,山东半岛,台湾,朝鲜,日本,印度尼西亚。
3.1.3三栉拟相手蟹ParasesarmatripectinisShen, 1940
Sesarma(Parasesarma)tripectinisShen 1940: 261-262, figs. 17-21.
Parasesarmaacis— Davie 1993: 65-74, figs. 1, 2.
Parasesarmatripectinis— Ng et al. 2008: 222. - Liu. 2008: 802.
样品采集:雷州企水海角村2只(1♀,1♂)。
外形描述:头胸甲近方形,头胸甲宽约长的1.25~1.3倍,背甲稍拱。表面隆起具分散颗粒,胃、心区具“H”形浅沟,鳃区具斜行隆线。额宽稍大于头胸甲宽度的1/2,弯向下方,额后4叶突出明显。外眼窝角尖锐,指向前方,侧缘无齿,近平行。螯足粗壮,等大,长节背面密布颗粒,前缘近末端具一锐刺;腕节内缘具颗粒,外缘及背面有条纹;雄性掌节背面具3列梳状栉,靠近可动指基部第一列具15~20个梳状齿,第二列约21个梳状齿,第3列具10~13个梳状齿;可动指背缘的突起约18~24个。步足中等长度,宽大扁平;第3对步足最长,约当头胸甲宽的1.7倍,长节长约长节宽的2.4倍。雄性第一腹肢末端几丁质突起长而趋尖,弯指向腹外方,几成直角。雄性腹部宽三角形,腹部第6节基部的宽度约当长度的2.6倍;尾节近圆形,长于第6节[4,15](图5,图6)。
分布:广东,福建,浙江,胶州湾,台湾,日本。

图5 三栉拟相手蟹(成体♂,头胸甲宽11.00 mm×长7.99 mm);A. 整体背面观;B. 正面观;C. 腹部;D. 左螯可动指正面观;E. 螯足掌节背面梳状栉;F. 左螯Fig.5 P. tripectinis(adult male, 11.00 mm×7.99 mm): A. overall dorsal views; B. frontal view of carapace; C. ventral view; D. dorsal surface of dactylus of left cheliped showing dactylar tubercles; E. dorsal surface of chela show-ing diposition of pectinate crest; F. left cheliped

图6 三栉拟相手蟹雄性第一腹肢:A~B为左雄性第一腹肢整体观及末端放大;比例尺为1 mm(仿Davie[15])Fig. 6 The male first gonopod of P. tripectinis: A-B. left male first gonopod and enlargement of tip of gonopod one; scal=1 mm (modified from Davie[15])
3.1.4ParasesarmaungulatumH. Milne Edwards, 1853
SesarmaungulateH. Milne Edwards 1853:184.
Sesarmaquadrata— A. Milne-Edwards 1873:302.
Sesarmaquadrata— Thallwitz 1892: 37. - De Man 1892: 328. - Lanchester 1900: 257. - Schenkel 1902: 549.
Sesarma(Parasesarma)quadrata— Lanchester 1902: 550.
Sesarma(Parasesarma)quadratavar.affinis— De Man 1902: 533.
Sesarma(Parasesarma)plicatum— Rathbun 1907: 34. - Rathbun 1910: 309.
Parasesarmaungulatum— Rahayu & Ng 2010:1-22, figs, 8-10.
样品采集:海南三亚青梅港9只(6♀,3♂) 。
形态描述:头胸甲宽约当长的1.23倍,胃、心区分区明显,肠区适度分区;背甲壳表面散布有稀疏的短刚毛,侧面具斜行隆线,侧缘具短刚毛。额弯向下方,中部稍凹,额后部的4叶突出。外眼窝角呈三角形,指向前方,侧缘无齿。螯足等大、粗壮;螯足长节呈三棱形,各边缘均呈锋锐隆脊状,内腹缘近末端具一锐刺;腕节背面近方形,表面粗糙具皱襞和颗粒;掌节厚而短,掌部外侧面具颗粒,内侧面的颗粒较细,掌中部有一列突起约9个;掌节背面具2梳状栉,分别具15~17及12个梳状齿,可动指背缘具11个纵行突起。雄性腹部相对较宽,尾节近圆形,长度略比第6节长;第六节宽度约当长度2.5倍,侧缘稍凸。雄性第一腹肢粗短,挺直,末端延伸稍弯曲呈60°,几丁质突起短而宽,顶端呈截状。雌性螯足相对较小,掌节基部背面的梳状栉则为2列突起所替代,可动指突起10~12个,不易区分[8](图7,图8)。
分布:海南,香港,泰国,马来西亚,新加坡,印度尼西亚。

图7 P. ungulatum(成体♂,头胸甲宽16.86 mm×长13.05 mm):A. 整体背面观;B. 正面观;C. 腹部;D. 右螯;E. 右螯可动指正面观;F. 螯足掌节背面梳状栉Fig.7 P. ungulatum(adult male, 16.86 mm×13.05 mm): A. overall dorsal views; B. frontal view of carapace; C. ventral view; D. right cheliped; E. dorsal surface of dactylus of right cheliped showing dactylar tubercles; F. dorsal surface of chela showing diposition of pectinate crest
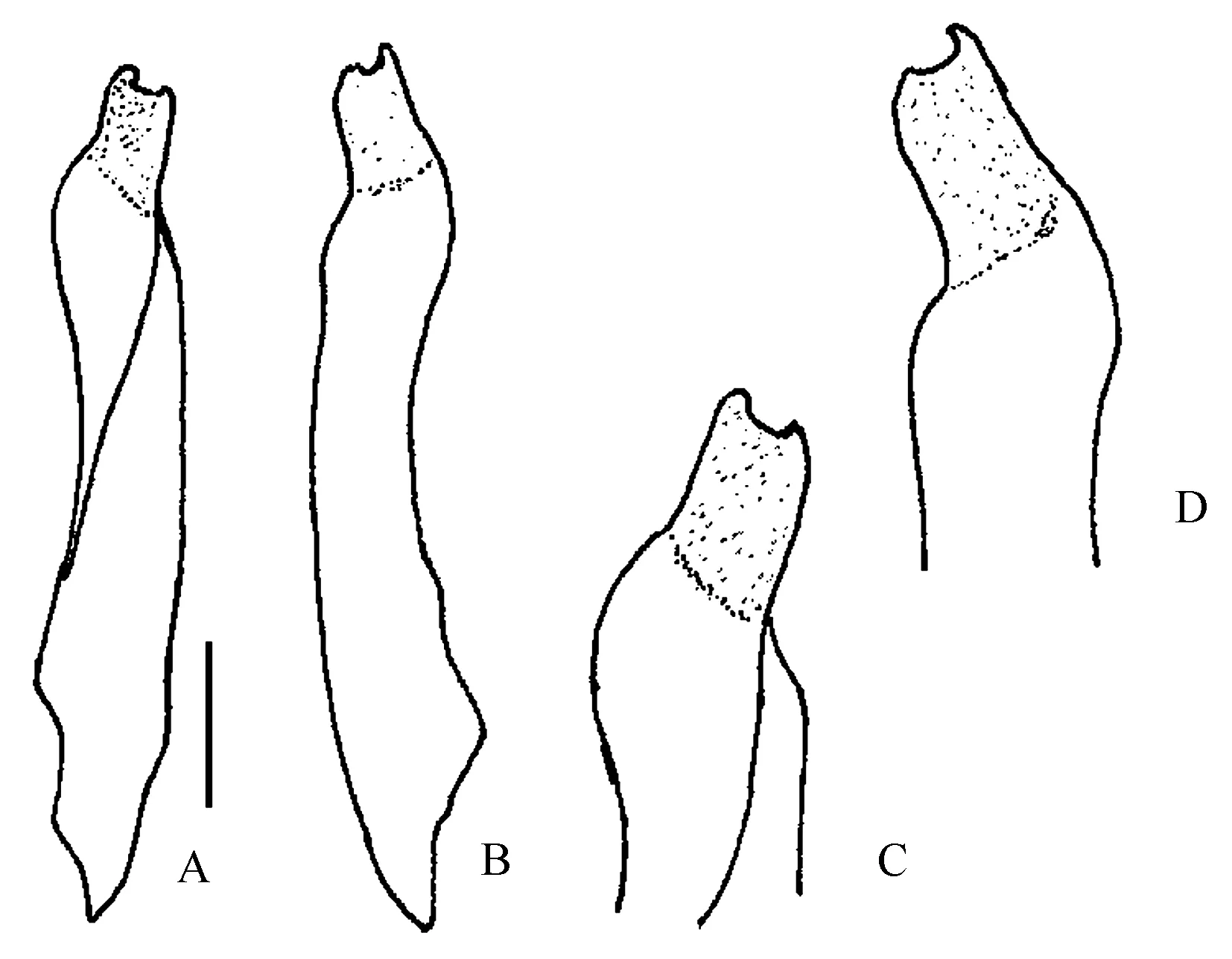
图8 P. ungulatum雄性第一腹肢:A~D为右雄性第一腹肢整体观及末端放大;比例尺为1 mm(仿Rahayu 和Ng[8])Fig.8 The male first gonopod of P. ungulatum: A-D. right male first gonopod and enlargement of tip of gonopod one; scal=1 mm (modified from Rahayu and Ng[8])
3.2序列分析与分子系统发育
5种拟相手蟹7个单倍型的16S rRNA基因序列对比获得494bp的同源序列(含插入/缺失位点),A+T平均含量为76.7%,其中T为39.2%,C为8.0%,A为37.5%,G为15.4%,除插入/缺失位点外共检测到58个变异位点,其中简约信息位点40个(图9)。5种拟相手蟹种间遗传距离达1.9%~9.3%(表2)。采用NJ法与贝叶斯法构建的系统发育树拓扑结构基本一致(图10),近亲拟相手蟹与褶痕拟相手蟹汇聚成一支,随后与P.ungulatum聚在一起,斑点拟相手蟹与三栉拟相手蟹汇聚成独立支系。

图9 5种拟相手蟹16S rRNA基因片段变异位点Fig.9 The variation sites of 16S rRNA gene fragment among five species Parasesarma

物种P.affinehap1P.affinehap2P.pictumP.tripectinisP.ungulatumhap1P.ungulatumhap2P.plicatumP.affinehap1P.affinehap20.002P.pictum0.0720.075P.tripectinis0.0770.0790.019P.ungulatumhap10.0520.0550.0660.066P.ungulatumhap20.0500.0530.0640.0680.002P.plicatum0.0480.0500.0880.0930.0570.055

图10 基于16S rRNA基因序列,采用NJ法、贝叶斯法构建5种拟相手蟹及外群的系统发育树Fig.10 Phylogenetic tree base on 16S rRNA of 5 species Parasesarma and outgroup using NJ and Bayesian inference (BI) methods发育树上结点的数值分别为NJ、BI的自引导值Significance values are list on the phylogenetic consensus tree in the order NJ, BI (bootstrap value)
4讨论
长期以来,诸多学者认为广布于印度-西太平洋的拟相手蟹为褶痕拟相手蟹,这一类群的外形特征为步足宽扁,雄性螯足掌节背面具2列梳状栉,可动指背缘的突起少于15个。因此Cancerquadrates、Ocypode(pachysoma)affinis、Sesarmaungulatum这3个种均被当作是褶痕拟相手蟹的同物异名种,并且为大多数学者所引用。《中国海洋蟹类》中描述的种名亦为褶痕拟相手蟹,其分布于中国南部沿海,为广温广布种[4]。Ng和戴爱云的后续研究中表明产于日本的Ocypode(pachysoma)affinis区别于褶痕拟相手蟹,它们属于不同的种类[16]。基于此,Ng等[1]将近亲拟相手蟹作为独立种与褶痕拟相手蟹区分开来。Rahayu 和Ng[8]检查了大量来自印度、东南亚、香港、南海以及日本的标本,发现褶痕拟相手蟹可动指背面突起为11~14个,突起较近亲拟相手蟹的明显且圆大,雄性第一腹肢较近亲拟相手蟹细长,末端几丁质突起尖细,较近亲拟相手蟹的弯曲,并且褶痕拟相手蟹分布于孟买、印度泰米尔纳德邦、泰国普吉岛、龙目岛,而近亲拟相手蟹广泛分布于中国海区、朝鲜半岛、日本、印度尼西亚。通过对比它们的螯足、腹部、雄性第一腹肢和雌性生殖孔的差异及分布区域,证实了近亲拟相手蟹与褶痕拟相手蟹确为各自独立物种,且广布于中国沿海的为近亲拟相手蟹,目前在中国沿海尚未发现褶痕拟相手蟹的分布。
Milne[17]在发现新种P.ungulatum时,并没有指出采集的样品数量及在西里伯斯岛的具体采集区域,仅有一个干样本和简要的描述,这增加了后来学者对区分这一物种与其他拟相手蟹的难度,Ng等[1]亦将P.ungulatum列为近亲拟相手蟹的同物异名种。学者们在做了诸多研究后,发现P.ungulatum与褶痕拟相手蟹、近亲拟相手蟹并非同物异名种,虽然它们的头胸甲形状和螯足可动指背面突起数相近,但是P.ungulatum的雄性第一腹肢较褶痕拟相手蟹、近亲拟相手蟹的粗短,挺直,几丁质突起较宽短,褶痕拟相手蟹雄性第一腹肢细长,末端弯曲呈45°,几丁质突起稍弯曲,顶端趋圆,而近亲拟相手蟹的雄性第一腹肢末端弯曲呈45°,几丁质突起趋直,它们的雄性第一腹肢差异明显。研究表明来自中国南部、香港、泰国、新加坡、马来西亚、巴布亚和印度尼西亚的一部分标本,实际上属于P.ungulatum,P.ungulatum为有效种[8]。

表3 6种拟相手蟹的形态比较
拟相手蟹检索表
Ⅰ.无前侧齿,掌节背面具2~3列横行梳状栉。可动指背面有一纵列突疣
拟相手蟹属(GenusParasesarma)
A. 可动指背面突起8~11个。
a. 可动指背面突起8~9个,雄性第一腹肢细长,末端趋长,弯曲呈45°,几丁质突起长而直,指向外方……………………………近亲拟相手蟹P.affine
b. 可动指背面突起11个,雄性第一腹肢粗短,挺直,末端延伸稍弯曲呈60°,几丁质突起短而宽,顶端呈截状
P.ungulatum
B. 可动指背面突起13~24个。
a. 可动指背面突起13~20个, 雄性第一腹肢粗短,末端趋尖,弯曲呈60°,几丁质突起短,呈截状斜面
斑点拟相手蟹P.pictum
b. 可动指背面突起18~24个, 雄性第一腹肢末端几丁质突起长而趋尖,弯指向腹外方,弯曲约呈90°…………………………三栉拟相手蟹P.tripectinis
c. 可动指背面突起17~19个,雄性第一腹肢细长,挺直近三棱形,末部的角质突起细锐,斜指向外侧方
精巧拟相手蟹P.exquisitum
近亲拟相手蟹与P.ungulatum、斑点拟相手蟹大小相近,在外形上也比较相似。从表3中的对比可以看出,近亲拟相手蟹螯足可动指背面突起8~9个,P.ungulatum螯足可动指背面突起11个,斑点拟相手蟹螯足可动指背面突起13~20个。斑点拟相手蟹的头胸甲较近亲拟相手蟹、P.ungulatum扁平,它们的雄性第一腹肢差异大(图2,图4,图8),因此可以相对容易地将三者区分开来。三栉拟相手蟹与精巧拟相手蟹体型较小,螯足可动指背面突起数较多,然而三栉拟相手蟹螯足掌节具3列梳状栉,可动指背面突起约18~24个;雄性第一腹肢较粗短,中度弯曲,几丁质长而趋尖,弯曲呈90°,明显区别于其他几种拟相手蟹因此这几种拟相手蟹可以从它们的螯足形状包括螯足掌节背面的梳状栉,可动指背面突起数目,以及雄性第一腹肢的形状进行区分(见检索表)。
基于16S rRNA序列分析表明,5种拟相手蟹的种间遗传差异达到1.9%~9.3%(表2),种间遗传差异远远大于种内遗传差异(0.2%),许多蟹类16S rRNA基因序列研究表明,种内的遗传差异一般在0.0%~1.0%之间[18—21],其上限为1.5%[22],因此,这5种拟相手蟹的遗传差异均达到了种间差异。近亲拟相手蟹与褶痕拟相手蟹的遗传差异达4.8%~5.0%,系统发育树显示近亲拟相手蟹形成独立支系再与褶痕拟相手蟹汇聚(图10),从分子水平上也证实近亲拟相手蟹与褶痕拟相手蟹确为不同的物种。系统发育树的拓扑结构和较高的置信度,支持斑点拟相手蟹与三栉拟相手蟹互为姐妹种(二者的遗传差异为最小值的1.9%),是5种拟相手蟹中最先分化出来的种类,与另外3种拟相手蟹的亲缘关系较远;P.ungulatum分化时间较近亲拟相手蟹和褶痕拟相手蟹早,而近亲拟相手蟹与褶痕拟相手蟹分化时间最晚,两者的亲缘关系较近,致使它们在形态上有更高的相似度。综合形态特征描述与16S rRNA序列分析,形态和分子证据均支持这5种拟相手蟹分别作为独立物种。
由于拟相手蟹种类之间的区分难度较大,相关研究较少,致使其在国内的种群数量、分布信息等不清,特别是P.ungulatum的相关信息以及中国是否有褶痕拟相手蟹的分布等问题,还需要做更广泛更深入的调查研究。
参考文献:
[1]Ng P K L, Guinot D, Davie P J F. Systema Brachyurorum: Part Ⅰ. An annotated checklist of extant brachyuran crabs of the world[J]. The Raffles Bulletin of Zoology, 2008(S17): 1-286.
[2]Davie P J F, Pabriks L. A new species ofParasesarma(Crustacea: Brachyura: Sesarmidae) from the mangroves of Western Australia[J]. Zootaxa, 2010, 2564: 62-68.
[3]Koller P, Liu H C, Schubart C D. A new semiterrestrial species ofParasesarmaDe Man, 1895, from Taiwan (Decapoda, Brachyura, Sesarmidae)[M]//Studies on Malacostraca: Lipke Bijdeley Holthuis Memorial Volume. Crustaceana Monographs, 2011, 14: 357-368.
[4]戴爱云, 杨思谅, 宋玉枝, 等. 中国海洋蟹类[M]. 北京: 海洋出版社, 1986: 482-491.
Dai Aiyun, Yang Siliang, Song Yuzhi, et al. Crabs of the China Seas[M]. Beijing: China Ocean Press, 1986: 482-491.
[5]Rahayu D L, Ng P K L. Two new species ofParasesarmaDe Man, 1895, from Southeast Asia (Crustacea: Decapoda: Brachyura: Sesarmidae)[J]. Zootaxa, 2009, 1980: 29-40.
[6]Naderloo R, Schubart C D. Description of a new species ofParasesarma(Crustacea; Decapoda; Brachyura; Sesarmidae) from the Persian Gulf, based on morphological and genetic characteristics[J]. Zoologischer Anzeiger-A Journal of Comparative Zoology, 2010, 249(1): 33-43.
[7]刘瑞玉. 中国海洋生物名录[M]. 北京: 科学出版社, 2008: 772-809.
Liu Ruiyu. Checklist of Marine Biota of China Seas[M]. Beijing: Science Press, 2008: 772-809.
[8]Rahayu D L, Ng P K L. Revision of theParasesarmaplicatum(Latreille, 1803) species-group (Crustacea: Decapoda: Brachyura: Sesarmidae)[J]. Zootaxa, 2010, 2327: 1-22.
[9]戴爱云, 宋玉枝. 广西北部湾潮间带蟹类的初步研究[G]//《甲壳动物学论文集》编辑委员会. 甲壳动物学论文集. 北京: 科学出版社, 1986: 54-62.
Dai Aiyun, Song Yuzhi. Intertidal crabs from Beibu Gulf of Guangxi[C]// Editorial Committee of Symposium on Crustacea. Symposium on Crustacean Research in China. Beijing: Science Press, 1986: 54-62.
[10]Shen C J. Four new species of Brachyura from Chinese seas[J]. Journal of the Hong Kong Fisheries Research Station, 1940, 1(2): 255-262.
[11]Schubart C D, Cannicci S, Vannini M, et al. Molecular phylogeny of grapsoid crabs (Decapoda, Brachyura) and allies based on two mitochondrial genes and a proposal for refraining from current superfamily classification[J]. Journal of Zoological Systematics and Evolutionary Research, 2006, 44(3): 193-199.
[12]Shih H T, Ng P K L, Liu M Y. Systematics of the Indo-West Pacific broad-fronted fiddler crabs (Crustacea: Ocypodidae: genusUca)[J]. The Raffles Bulletin of Zoology, 2013, 61(2): 641-649.
[13]Palumbi S R. Genetic divergence, reproductive isolation, and marine speciation[J]. Annual Review of Ecology and Systematics, 1994, 25: 547-572.
[14]Swofford D L. PAUP*: phylogenetic analysis using parsimony, version 4. 0 b10[Z]. Sunderland, MA, USA: Sinauer Associates, 2003.
[15]Davie P J F. A new species of Sesarmine crab (Brachyura: Grapsidae) from Japan and Taiwan, previously known asSesarmaerythodactylaHess, 1865[J]. Crust Res, 1993, 22: 65-74.
[16]Ng P K L, Wang C H, Ho P H, et al. An annotated checklist of brachyuran crabs from Taiwan (Crustacea: Decapoda)[M]//National Taiwan Museum Special Publication Series, No. 11. Taipei, Taiwan, China: National Taiwan Museum, 2001: 1-86.
[17]Milne-Edwards H. Mémoire sur la famille des Ocypodiens[G]//Annales des Sciences Naturelles (Zoologie) Series 3. 1853, 20: 163-228.
[18]Yeo D C J, Shih H T, Meier R, et al. Phylogeny and biogeography of the freshwater crab genus Johora (Crustacea: Brachyura: Potamidae) from the Malay Peninsula, and the origins of its insular fauna[J]. Zoologica Scripta, 2007, 36(3): 255-269.
[19]Shih H T, Ng P K L, Schubart C D, et al. Phylogeny and phylogeography of the genusGeothelphusa(Crustacea: Decapoda, Brachyura, Potamidae) in southwestern Taiwan based on two mitochondrial genes[J]. Zoological Science, 2007, 24(1): 57-66.
[20]Shih H T, Suzuki H. Taxonomy, phylogeny, and biogeography of the endemic mudflat crabHelice/Chasmagnathuscomplex (Crustacea: Brachyura: Varunidae) from East Asia[J]. Zoological Studies, 2008, 47(1): 114-125.
[21]徐敬明, 孙翰昌, 孙世春. 厚蟹线粒体16S rRNA基因序列分析及系统发育研究[J]. 海洋学报, 2011, 33(5): 124-132.
Xu Jingming, Sun Hanchang, Sun Shichun. Molecular phylogeny of the crab generaHeliceandHelicana(Crustacea, Decapoda) based on the partial sequences of mitochondrial 16S rRNA gene[J]. Haiyang Xuebao, 2011, 33(5): 124-132.
[22]Schubart C D, Koller P. Genetic diversity of freshwater crabs (Brachyura: Sesarmidae) from central Jamaica with description of a new species[J]. Journal of Natural History, 2005, 39(6): 469-481.
收稿日期:2015-11-12;
修订日期:2016-03-13。
基金项目:广西红树林保护与利用重点实验室基金(GKLMC-201205);“红树林和海草生态系统保育与生态监测”特聘专家岗位专项。
作者简介:杨明柳(1988—),女,广西壮族自治区桂林市人,主要从事动物资源保护学研究。E-mail:yangmingliu2010@163.com *通信作者:徐敬明,教授,博士,主要从事蟹类形态与分子生理生态研究。E-mail:xjingming@163.com
中图分类号:Q959.223
文献标志码:A
文章编号:0253-4193(2016)08-0062-11
Taxonomic study on the genus Parasesarma based on the morphology characteristics and mtDNA 16S rRNA gene
Yang Mingliu1,Xu Jingming2,Wu Bin1,Yan Bing1
(1.GuangxiKeyLaboratoryforMangroveConservationandUtilization,GuangxiAcademyofSciences,GuangxiMangroveResearchCenter,Beihai536000,China; 2.CollegeofForestryandLifeScience,ChongqingUniversityofArtsandSciences,Yongchuan402168,China)
Abstract:The taxonomy of the genus Parasesarma is one of the most problematic in the Sesarmidae, with their very similar morphology. We compared the morphology characteristics of Parasesarma affine, P. pictum, P. tripectinis, P. ungulatum(from the coast of China) and P. plicatum(from Singapore), and found that these five species could be distinguished from the shape of chelipeds(which included pectinate crest of dorsal surface of palm, the number of dactylar tubercles of the chelipeds)and the male first gonopod. The partial sequences of mitochondrial DNA 16S rRNA gene of these five Parasesarma species were contributed to molecular phylogenetic analysis, and the result showed that the genetic distances was 1.9%~9.3% between interspecies, and had reached the level difference of interspecies; the phylogenetic tree indicated that P. affine and P. plicatum were clustered together and then clustered into a clade with P. ungulatum, P. pictum and P. tripectinis were clustered into a distinct clade. Both of evidence of morphological and molecular data supported that the five Parasesarma species were separate and valid species.
Key words:Parasesarma; morphology characteristics; 16S rRNA gene; taxonomic study
杨明柳,徐敬明,吴斌,等. 基于形态和16S rRNA基因的拟相手蟹分类研究[J].海洋学报,2016,38(8):62—72, doi:10.3969/j.issn.0253-4193.2016.08.007
Yang Mingliu,Xu Jingming,Wu Bin, et al. Taxonomic study on the genusParasesarmabased on the morphology characteristics and mtDNA 16S rRNA gene[J]. Haiyang Xuebao,2016,38(8):62—72, doi:10.3969/j.issn.0253-4193.2016. 08.007
