毛竹幼苗对不同氮含量及形态的营养响应
2016-08-07顾红梅邓光华杨海燕杨清培
顾红梅,邓光华,黄 玲,杨海燕,杨清培,2,邹 娜,2
(1.江西农业大学 园林与艺术学院, 江西 南昌 330045;2.江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045)
毛竹幼苗对不同氮含量及形态的营养响应
顾红梅1,邓光华1,黄 玲1,杨海燕1,杨清培1,2,邹 娜1,2
(1.江西农业大学 园林与艺术学院, 江西 南昌 330045;2.江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045)
采用室内可控环境盆栽方法研究毛竹幼苗对不同氮供应强度和形态的响应。结果表明,在低氮条件下不同氮素形态造成的生长差异很小;随着氮浓度的提高,毛竹幼苗苗高,叶绿素含量,叶片数,叶面积,地上地下生物量和根系构型等各生长指标在铵氮供应条件下的生长显著优于以硝氮为唯一氮源。在pH3.8条件下,毛竹幼苗能正常生长,而且生长各指标对不同氮素形态具有pH5.8条件下相似的响应特征,表明毛竹幼苗对土壤pH的适应性较广,对酸性环境具有较强的耐受能力。
毛竹;氮含量;氮形态;生长响应
毛竹(Phyllostachysheterocyclacv.Pubescens)属禾本科刚竹属,为我国分布最广,面积最大,经济价值最高的竹种[1]。它具有成长快,成材早,产量高,收益大,用途广等特点,成为我国南方重要的森林资源,在我国林业生产中占有非常重要的地位[2]。虽然竹林扩张为林业发展、林农增收发挥了积极的作用[3-4],但近年来毛竹林向周边邻近常绿阔叶林或针阔混交林边界扩张蔓延趋势日益明显,并引发了一系列生态问题,如影响邻近生态系统的群落组成与结构、生物多样性、土壤性质、生态过程与功能, 并造成生态景观的破坏[5]。
氮素是植物生长不可或缺的营养元素,在森林生态系统中,土壤有效氮主要以NH4+-N和NO3--N形式存在,NH4+-N和NO3--N也是植物从土壤中吸收N的主要形式。由于不同植物对氮素的吸收利用能力不同,并且在长期的进化过程中形成了对不同氮素形态的偏向选择性即在N03-(或NH4+)占优势的N 营养环境中,植物吸收N 的能力、生理反应、生长速度较好,表现出明显的喜硝性(或喜铵性)[6]。喜NH4+植物如针叶树种、杜鹃科植物和水稻等[6-8],而另一些植物在以纯N03-或以N03--N 占优势的营养环境中生长的较好,如大麦和番茄等多数农作物和蔬菜[9-10]。
近几十年来, 由于化石燃料和农牧业含氮化肥饲料的广泛使用, 全球大气氮沉降量明显增加, 一些地区已经达到饱和甚至超过了生态系统所能承受的临界负荷[11]。如在我国江西分宜县大冈山林场的杉木(CunninghamiaLanceolata)林和马尾松(Pinusmassoniana)林中,降雨氮输入分别为 60.6 kg·hm-2·a-1和57.0 kg·hm-2·a-1[12],这些数字均高于或远远高于森林在生长季节对氮的需求量(约 5~8 kgN·hm-2·a-1)[13]。
不同植物的N 营养特性及对土壤N 供给变化的特异性反应对森林生态系统中优势植物间的竞争关系和森林发展过程中物种的选择起着非常重要的作用[14-16]。近年来,毛竹扩张趋势不断加强,其是否与近年来氮沉降量显著增加造成的土壤氮供给变化及土壤酸化等有关尚不清楚。因此,本试验以毛竹为材料,研究毛竹幼苗对不同N水平梯度,不同NH4+/NO3-配比,不同pH值环境下的营养响应,以期从土壤氮营养角度对毛竹扩张进行探讨并为毛竹育苗和人工林的培育提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为毛竹幼苗。毛竹种子采自广西桂林灌阳县洞井乡。
1.2 试验方法
1.2.1 种子处理及培养 毛竹种子先置于40 ℃温水浸种12 h,然后用0.1%代森锰锌溶液消毒12 h,取出后放入培养皿中并置于人工气候箱中进行种子萌发。培养温度(25±1)℃,光照时间12 h·d-1,光照强度为2 000 Lx,相对湿度65%。
1.2.2 试验处理 待第2片真叶完全展开后,挑选长势一致的毛竹实生苗移入装有灭菌蛭石的花盆中(规格为130 mm×150 mm)。试验苗移栽后15天,用含有不同氮浓度及铵硝配比(NH4+/NO3-)的无机氮源培养两个月,氮浓度设低N(0.4 mM)、中N(2 mM)和高N(8mM)3个水平, NH4+-N/NO3--N比为100/0、50/50和0/100共9个处理,每个处理16株苗。NH4+以(NH4)2SO4形式提供,NO3-以KNO3和Ca(NO3)2·4H2O形式提供,每周浇一次营养液。用于配置不同氮处理的营养液组成修改自Norisada M and Kojima K (2005),见表1。所有营养液中均加硝化抑制剂双氰胺(C2H4N27 μm·L-1)充分抑制硝化作用,保障NH4+/NO3-比例不发生很大的变化,用pH计(Orion 86801)和0.1 mol·L-1NaoH或0.1 mol·L-1HCI调节所有处理的营养液pH值保持在5.8或3.8左右。除了特殊说明,实验中所获得数据均为pH 5.8条件下的。
1.3 研究方法
苗高和叶片数采用常规的方法测量;叶绿素含量(SPAD值)用叶绿素仪(SPAD-502,Minolta)取顶端第2、3片叶进行测定;叶面积采用叶面积仪YMJ-A测定。采收后的苗用清水将根部洗净,地下部根系用EPSON COLOR IMAGE SCANNER LA1600+扫描仪进行扫描,然后用根系图象分析软件WinRHIZO 2012对各处理根系的体积、表面积、总长,根尖数和平均直径进行定量测定。此后将苗分为根、茎、叶3部分经杀青(105 ℃,30 min)后70 ℃烘至恒重并称重。
1.4 数据处理
所有数据采用 Excel 2003和SPSS 13.0进行统计分析,图表使用Sigmaplot 13.0生成并由Photoshop排版。
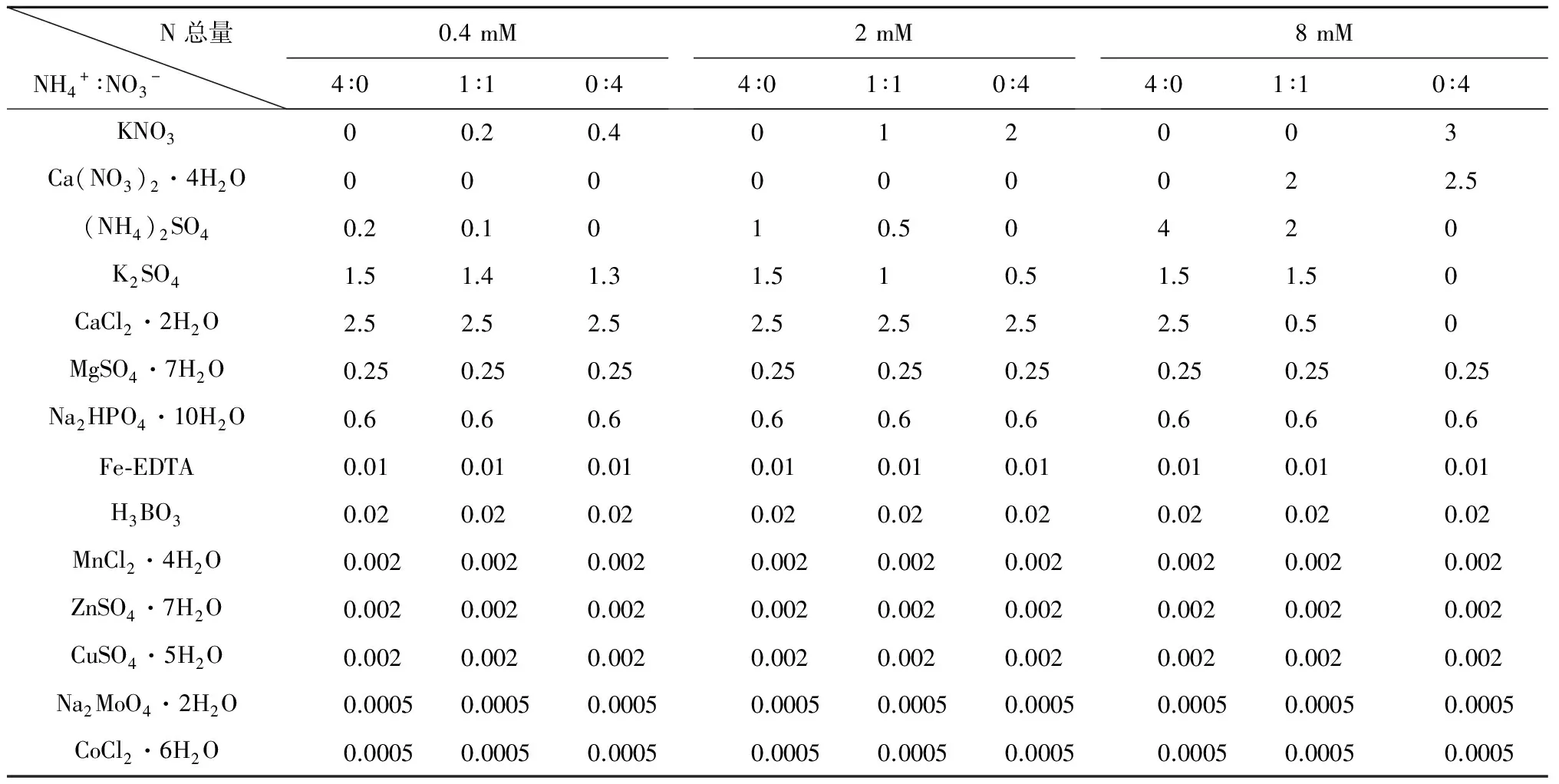
表1 不同N含量及形态营养液组成
Concentrations are in mM.
2 结果与分析
2.1 不同氮含量及形态对毛竹地上部生长和叶绿素含量的影响
图1结果表明,随着氮含量的增加,毛竹幼苗生长和叶绿素含量在不同氮素形态间的差异增大。在低氮浓度时(0.4 mM),等浓度的不同氮素形态对毛竹幼苗株高和叶片数量间的影响差异不大;在中浓度时(2 mM),铵氮处理的幼苗株高和叶片数量显著高于经等浓度的硝态氮及铵硝混合氮源处理的幼苗;在高浓度时(8 mM),经铵氮及铵硝混合氮源处理的幼苗株高和叶片数量显著高于经等硝态氮浓度的处理的幼苗。此外,毛竹幼苗株高随着铵氮浓度的增加而显著增加,而硝氮浓度增加对毛竹幼苗高生长并未有明显影响(图1A)。
毛竹幼苗单株叶片数量在铵氮中具有最大值,并且与硝氮间的差异在中高氮浓度条件下达到显著水平(图1B)。铵氮处理幼苗单株叶面积显著高于硝氮,并且这种差异随着氮浓度的增加而增大;此外,叶面积基本随着铵氮浓度的增加而增加,但经高硝氮浓度处理的毛竹幼苗叶面积显著低于中低浓度处理的(图1C)。
不同氮素形态对毛竹幼苗叶绿素含量的影响差异随着氮含量的增加而增大;就氮形态而言铵处理的幼苗叶绿素含量显著高于硝氮处理的幼苗(图1D)。上述结果表明一定浓度的铵氮更有利于毛竹幼苗地上部生长和叶绿素积累。
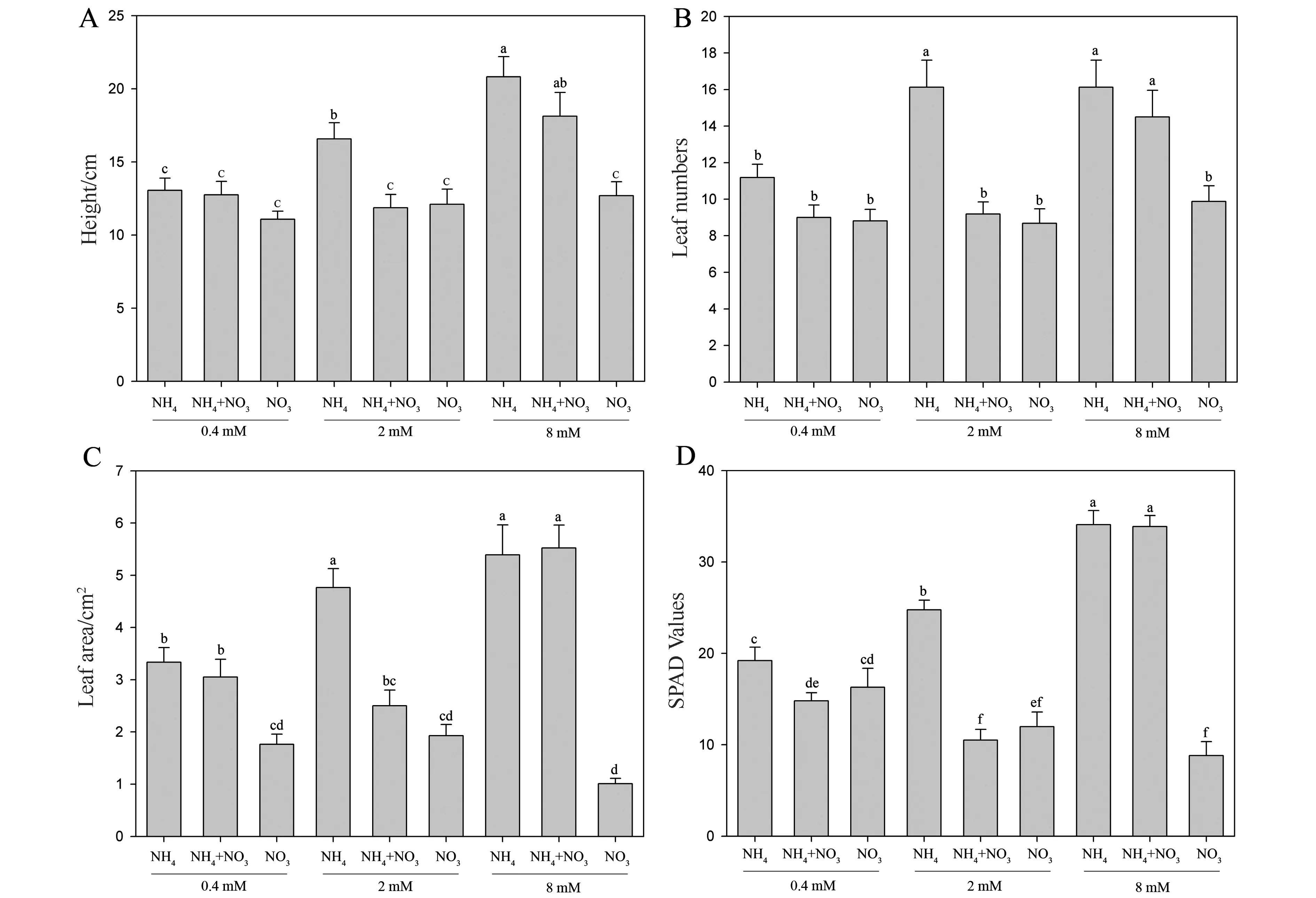
图1 不同氮含量及形态对毛竹株地上部生长和叶绿素含量的影响Fig.1 Effects of N concentration and forms on the growth and chlorophyll content (SPAD) of Phyllostachys heterocycla cv. Pubescens
2.2 不同氮含量及形态对毛竹根系构型的影响
毛竹实生苗地下根系为须根,根系发达。表2结果表明,不同氮含量及氮素形态对毛竹根系各参数的影响不同,其中以高浓度的铵硝混合氮源对根系各参数如根长、根系表面积、根系平均直径、根系体积和根尖数具有较好的促进作用。就不同氮素形态对根系各指标的生长而言,以铵氮显著优于硝氮,且这种生长优势随着氮浓度的增加。
2.3 不同氮含量及形态对毛竹各部位生物量及根冠比的影响
毛竹各部分生物量以叶最大,其次是根,茎最小,各部位干物质的积累随着氮总量的增加而呈增加趋势(图2)。不同氮素形态对各指标的影响差异随着氮浓度的增加而增大,就氮形态而言混合氮源优于铵氮及硝氮(图2A)。毛竹总干重以8mM 的混合氮源最优,但与等浓度的铵氮间差异不显著;毛竹干物质的积累以铵氮优于硝氮,并且这种差异随着氮浓度的增加而增大(图2B)。根冠比随着营养液中硝浓度的增加基本呈上升趋势,但在中低氮浓度条件下,各处理间差异不显著。
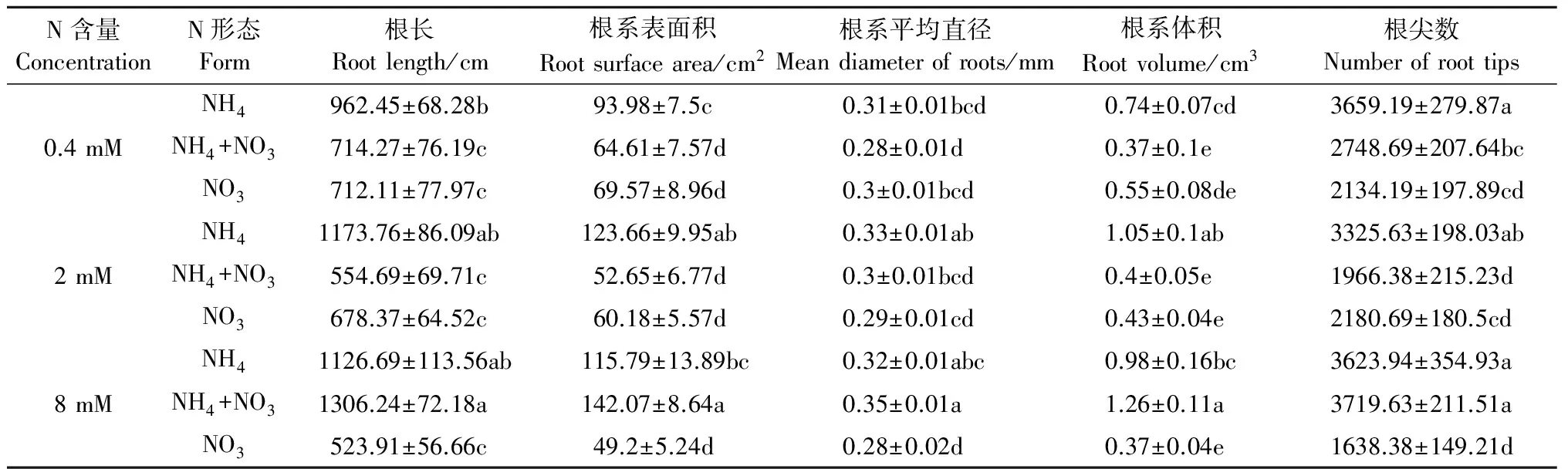
表2 不同氮含量及形态对毛竹根系构型的影响
注:表中的值为平均值±SE,同列数据后不同小写字母表示处理间差异显著(P<0.05)。
Note:The values represent the average ± SE, different letters within the same column mean significantly different atP﹤0.05.
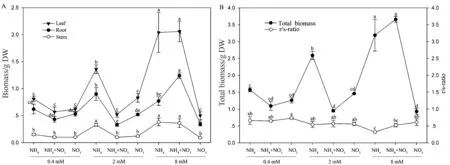
图2 不同氮含量及形态对毛竹各部位生物量及根冠比的影响Fig.2 Effects of N concentration and forms on biomass and root stem ratio of Ph.heterocycla cv. Pubescens
2.4 不同pH条件下氮含量及形态对毛竹生长各参数的影响
由于一般的文献研究所采用的营养液pH值都为5.8,而我们野外试验发现毛竹生长的土壤环境pH值低于4.0[18],因此我们在试验过程中设置了两个不同的pH值。图3结果表明,在不同pH条件下,毛竹各生长参数对处理中的氮素含量及形态具有相似的变化规律。除了少数参数的个别点在不同pH值条件下有差异外,绝大部分指标在不同pH值条件下对氮含量及形态的响应无明显影响(图3)。
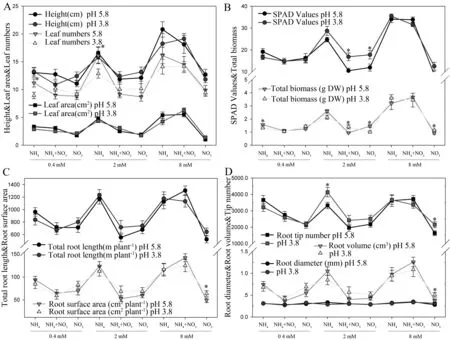
图3 不同pH条件下氮含量及形态对毛竹生长各参数的影响Fig.3 Effects of N concentrations and forms on growth index of Ph. heterocycla cv. Pubescens under different pH
3 讨论与小结
3.1 讨论
研究结果表明毛竹幼苗生长是氮素供给浓度和氮素形态共同作用的结果。当氮素浓度为0.4 mmol·L-1时,氮素形态差异造成的生长差异很小甚至是不存在的,如苗高和叶片数(图1A,B)及根茎叶各部位生物量及根冠比(图2)等在不同氮素形态间的差异均未达到显著水平。但随着氮供应强度的增加,丝栗栲幼苗生长在不同氮素形态间的差异逐渐增大,在8.0 mmol·L-1铵氮供应条件下的幼苗地上部生长(表1)和根茎叶各部位生物量及总重(图1)显著高于等浓度的硝氮。这些结果说明像大多数酸性原始森林中树种(如如很多松柏类树种)一样[19],毛竹是典型的喜铵树种。
植物能否有效获得土壤养分,在很大程度上取决于其根系形态和构型,另一方面土壤养分有效性及分布对植物根系也具有一定的调控作用[20]。本试验的研究结果表明,与偏喜铵态氮相一致,施用铵态氮相比于单一使用硝态氮素对毛竹根系构型各指标(如根长、根系表面积、根尖数、根系直径和根系体积)生长有更好的促进效应。
此外,在中低浓度时毛竹生物量积累和根系构型各指标生长以铵氮优于铵硝混合氮源,但高浓度时根系生物量、根系表面积和体积在铵硝混合氮源处理条件下最好。其原因一方面可能是铵态氮和硝态氮对毛竹幼苗生长的作用机制不同,铵硝混合氮源处理对根系生长具有协同增效的作用[7,20];另一方面也可能是NH4+作为一种典型的对植物生长呈现自相矛盾的养分离子,即:是植物生长必需的养分离子及重要的中间代谢产物,但高浓度时对植物毒害作用又非常明显[21]。生长在较高NH4+环境中,植物时常会表现出铵毒害症状,如:根系粗短、根冠比下降、叶子黄化、种子萌发及植株整体生长受抑制、严重时发生死亡[22]。本研究中,8mM 铵氮处理与铵硝氮源混合处理可能对毛竹根系生长已造成了一定的抑制作用,其机制是否与铵毒害有关尚需进一步研究。
氮沉降量增加导致红壤酸化加剧,我国南方已成为继欧美之后的第三大酸沉降区。本研究中,在不同pH(pH 3.8和pH 5.8)条件下进行培养,发现毛竹幼苗都能正常生长,而且生长各指标对不同氮素形态具有相似的响应特征,说明毛竹对土壤pH变化的适应范围较广,而且具有较强的耐酸能力,这也与毛竹能在当前酸性土壤环境中正常生长相一致。目前我国大气氮沉降以NH4+-N为主(其中NH4+-N占2/3), NH4+-N比国外高3~10倍, NO3-则略低于国外水平[11],而且毛竹适生地土壤中NH4+-N 含量也超过60.0%[18],这与毛竹的氮形态偏好相吻合。因此,我们的研究结果表明,毛竹扩张可能与大气氮沉降有关,但尚需进一步研究论证。
3.2 小结
毛竹幼苗苗高和生物量等生长指标随着氮浓度特别是铵氮的提高而呈增加的趋势,但硝氮含量的增加对毛竹生长并没有显著影响,甚至对叶面积和叶绿素含量呈抑制作用,表明毛竹属于喜铵树种,而且对铵态氮肥的需求量较大。因此,在毛竹育苗及竹林培育过程中氮肥施用应以铵氮为主。此外,毛竹幼苗对土壤pH的适应性较广,对酸性环境具有较强的耐受能力。因此,我们的研究结果表明,当前大气氮沉降增加在一定程度上可能会促进毛竹扩张。
[1] 卢榕泉.抚育施肥措施对毛竹林笋产量的影响及效益分析[J].亚热带农业研究,2008,4(4):261-263.
[2] 郭晓敏,牛德奎,张斌,等.集约经营毛竹林平衡施肥效应研究[J].西南林学院学报,2005,25(4):84-88.
[3] Pérez MR, Zhong MG, Belcher B, et al. The role of bamboo plantations in rural development: The case of Anji county, Zhejiang, China [J]. World Development, 1999, 27(1):101-114.
[4] 萧江华.我国竹业发展现状与对策[J].竹子研究汇刊,2000,19(1):1-5,8.
[5] 杨清培,杨光耀,宋庆妮,等.竹子扩张生态学研究:过程、后效与机制[J].植物生态学报,2015,01:110-124.
[6] 崔晓阳,宋金凤.原始森林土壤NH4+/NO3-生境特征与某些针叶树种的适应性[J].生态学报,2005,25(11):3 082-3 092.
[7] Kronzucker HJ, Siddiqi MY, Glass ADM, et al. Nitrate-ammonium synergism in rice: A subcellular analysis[J]. Plant Physiology. 1999,119(3):1 041-1 046.
[8] 郭亚芬,孔凡婧,付连云,等.不同 NH4+/NO3-配比对红松幼苗生长的影响[J].土壤通报,2010,41(4): 923-926.
[9] Britto DT, Kronzucker HJ. Constancy of nitrogen turnover kinetics in the plant cell: Insights into the integration of subcellular N fluxes [J]. Planta, 2001,213(2):175-181.
[10] Lu YL, Xu YC, Shen QR, et al. Effects of different nitrogen forms on the growth and cytokinin content in xylem sap of tomato (LycopersiconesculentumMill.) seedlings [J]. Plant and Soil, 2009,315(1):67-77.
[11] 周薇,王兵,李钢铁,等.大气氮沉降对森林生态系统影响的研究进展[J].中央民族大学学报(自然科学版),2010,19(1):34-40.
[12] 马雪华.在杉木林和马尾松林中雨水的养分淋溶作用[J].生态学报,1989,9(1):15-20.
[13] 郝吉明,谢绍东,段雷,等. 酸沉降临界负荷及其应用[M].北京:清华大学出版社,2001.
[14] 王晋萍,董丽佳,桑卫国,等.不同氮素水平下入侵种豚草与本地种黄花蒿、蒙古蒿的竞争关系[J].生物多样性,2012,20(1):3-11.
[15] Butler SM, Melillo JM, Johnson JE, et al. Soil warming alters nitrogen cycling in a New England forest: implications for ecosystem function and structure [J]. Oecologia, 2012,168 (3):819-828.
[16] Novem Auyeung DS, Suseela V, Dukes JS. Warming and drought reduce temperature sensitivity of nitrogen transformations [J]. Global Change Biology, 2013, 19(2): 662-676.
[17] Norisada M, Kojima K. Nitrogen form preference of six dipterocarp species[J]. Forest Ecology and Management, 2005, 216(1): 175-186.
[18] 宋庆妮,杨清培,刘骏,等.毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响[J].应用生态学报,2013,24(2):338-344.
[19] Yao B, Cao J, Zhao C, et al. Influence of ammonium and nitrate supply on growth, nitrate reductase activity and N-use efficiency in a natural hybrid pine and its parents[J].Journal of Plant Ecology,2011,4(4):275-282.
[20] 邹娜,强晓敏,施卫明.不同供铵水平对番茄根系生长的影响[J].土壤,2012,05:827-833.
[21] Kronzucker HJ, Britto DT, Davenport RJ, et al.Ammonium toxicity and the real cost of transport [J]. Plant Science, 2001, 6(8):335-337.
[22] 李保海,施卫明.拟南芥幼苗对高NH4+响应的特征及不同生态型间的差异[J]. 土壤学报,2007,44(3):508-515.
Growth Response of Bamboo Seedlings to Nitrogen Nutrition inPhyllostachysheterocyclacv.Pubescens
GU Hong-mei1, DENG Guang-hua1, HUANG Ling1, YANG Hai-yan1, YANG Qing-pei1,2, ZOU Na1,2
(1.College of Landscape and Art, Jiangxi Agricultural University, Nanchang 330045, Jiangxi,China;2.Jiangxi Provincial Key Laboratory for Bamboo Germplasm Resources and Utilization,Jiangxi Agricultural University, Nanchang 330045, Jiangxi, China)
Seedlings ofPhyllostachysheterocyclacv.Pubescenswere supplied with different content of nitrogen as nitrate, ammonium, or both in vermiculite culture in a controlled condition. The results showed that there were little differences in the growth ofP.heterocyclacv.Pubescensto different N forms under low content; while with the increase of N concentration, the growth indicators such as seedling height, chlorophyll content (SPAD), leaf numbers, leaf area, biomass and root system configurations performed significantly better with the treatment of ammonium than that of nitrate as the sole nitrogen source. The seedlings showed similar growth response to N source supplied under ether pH 3.8 or pH 5.8, indicating their wide adaptability to soil pH and high tolerance to acid environment.
Phyllostachysheterocyclacv.Pubescens; N concentration; N form; Growth response
2015-01-26
国家自然科学基金青年科学基金项目(31300521)和高等学校博士学科点专项科研基金(20133603120001)资助
顾红梅(1991-),女,在读硕士,主要从事植物氮营养方面的研究。通信作者:邹娜,博士,副教授,主要从事园林植物生物技术及氮营养生理方面的研究。E-mail:nzouyy@126.com
