流溪河常绿阔叶林常见种幼苗非结构性碳水化合物研究
2016-08-06黄润霞贾小容吴回军饶书培罗婉莹林书航
黄润霞,贾小容,吴回军,饶书培,罗婉莹,李 灵,林书航
(1.华南农业大学林学与风景园林学院,广东 广州 510642;2.广东省乐昌林场,广东 韶关 512219)
流溪河常绿阔叶林常见种幼苗非结构性碳水化合物研究
黄润霞1,贾小容1,吴回军2,饶书培1,罗婉莹1,李 灵1,林书航1
(1.华南农业大学林学与风景园林学院,广东 广州 510642;2.广东省乐昌林场,广东 韶关 512219)
为探讨常绿阔叶林常见种幼苗非结构性碳水化合物(NSC)及其组分含量的种间差异及其在根、茎、叶不同器官中的分配格局,以流溪河森林公园常绿阔叶林15种常见种幼苗为研究对象,分别测定其根、茎、叶的可溶性糖、淀粉、非结构性碳水化合物的含量和可溶性糖/淀粉的比值。结果表明,物种、植物器官及其交互作用对植株非结构性碳水化合物及其组分含量具有显著影响;15种植株幼苗普遍表现为叶片的可溶性糖和可溶性糖/淀粉比值最高,根部的淀粉和非结构性碳水化合物含量最高;乔木树种幼苗和灌木树种幼苗的NSC及其组分含量在植物根部和叶片中无显著差异,乔木和灌木树种幼苗茎的可溶性糖与NSC含量也无显著差异,但常绿乔木的茎可溶性糖/淀粉显著高于灌木,淀粉含量显著低于灌木。常绿阔叶林常见种幼苗的NSC及其组分含量的种间差异及其在不同器官中的分布差异,体现了植株幼苗的叶和根分别是非结构性碳水化合物的合成与储存结构,为植株的生长发育提供营养物质,同时也反映了其应对环境的不同生存策略。
幼苗;非结构性碳水化合物;常绿阔叶林;流溪河
黄润霞,贾小容,吴回军,等.流溪河常绿阔叶林常见种幼苗非结构性碳水化合物研究[J].广东农业科学,2016,43(6):81-89.
非结构碳水化合物(non-structural carbohydrates,NSC)是对植物碳分配与碳消耗差值的量化,能够反映植物利用光合产物的水平[1],以及植物对其生存环境的响应。非结构性碳水化合物在植物器官之间的流动反映了碳吸收和碳消耗的平衡关系[2],反映了植物体碳供应水平,是一种动态变化的过程[3]。非结构性碳水化合物主要包括可溶性糖(如葡萄糖、蔗糖、果糖等)和淀粉[4],可溶性糖是光合作用的直接产物,是碳运输和代谢过程中的主要形式,用于满足植物当前生命活动需要,并起到调节渗透压的作用;淀粉是植物主要的长期贮存形式,主要供应植物将来需求[5-7]。两者在一定条件下可以相互转换[8]。可溶性糖与植物的抗逆性密切相关,可溶性糖浓度高,植物的生命活动较旺盛[9]。另外,可溶性糖/淀粉比值可以直接表示植物体内可溶性糖、淀粉的分配,反映植物体的抗逆能力,在一定程度上反映出植物对外界环境的适应。
有关流溪河森林公园森林生态系统的研究不多,主要包括植物区系[10-11]、水文效益[12-13]以及森林资源调查[14]等方面,作为一个国家级的森林公园,上述研究对其森林生态系统结构和功能的揭示是不够的。本文以流溪河森林公园常绿阔叶林15种常见种幼苗为研究对象,探讨非结构性碳水化合物及其组分含量的种间差异和其在根、茎、叶不同器官中的分配格局,深入了解植物幼苗时期对环境的适应情况和其应对环境的生存策略,为进一步揭示并评价幼苗的生长状况,更好地保护和管理森林公园的植物群落提供科学依据。
1 材料与方法
1.1 研究地概况
流溪河森林公园位于广州市郊从化市北部境内,距广州市区93 km,地理坐标23°32′~23° 50′N、113°45′~113°54′E,属于南亚热带湿润季风气候区。空气相对湿度79%。年均气温20.3℃,极端最高气温39.2℃,极端最低气温6.8℃,1月平均气温11.5℃,7月平均气温27.4℃。年平均降水量为2 104.7 mm,全年各月均有降水,但季节分配不均匀。植被为亚热带季风常绿阔叶林,植被外貌终年常绿,结构复杂[12-13]。
1.2 试验方法
于2014年7月在流溪河森林公园常绿阔叶林内随机选取15种生长良好、健康的树种幼苗为研究对象,分别为灌木树种:山血丹(Ardisia lindleyana)、香港瓜馥木(Fissistigma uonicum)、九节(Psychotria rubra)、草珊瑚(Sarcandra glabra)、狗骨柴(Diplospora dubia)、三桠苦(Evodia lepta)和八角枫(Alangium chinense);乔木树种:红枝蒲桃(Syzygium rehderianum)、鸭脚木(Schefflera octophylla)、猴耳环(Pithecellobium clypearia)、黧蒴(Castanopsis fissa)、米锥(Castanopsis chinensis)、罗浮锥(Castanopsis fabri)、木荷(Schima superba)和亮叶猴耳环(Pithecellobium lucidum)。每种幼苗取3株,幼苗高度在0.3~0.7 m左右,将每株幼苗按根、茎、叶分开,所有样品获取后立即装入密封袋中保鲜,于当日带回实验室,用清水把植株幼苗的根部漂洗干净,除去表面粘附的土壤,晾干;将所有样品置于105℃恒温箱中杀青20 min后,于75℃烘干至恒重,备用。
1.3 指标测定
植物样品中的可溶性糖用无水乙醇提取,淀粉用高氯酸法提取[15],均借鉴Buysse等[16]的测定方法,利用蒽酮-浓硫酸比色法测定可溶性糖、淀粉含量。NSC含量为可溶性糖含量与淀粉含量之和。
1.4 数据统计分析
数据通过Microsoft Excel进行整理,利用Statistica12.5统计分析软件进行二元方差分析(Two-way ANOVA)、一元方差分析(One-ANOVA)和Duncan’s多重比较,并作图。
2 结果与分析
2.1 15种常见幼苗NSC含量分析
15种常见种幼苗可溶性糖含量为41.8(± 8.85)~132.79(±38.02)mg/g,其中鸭脚木最大,其次为木荷(92.02±31.98 mg/g),草珊瑚最小;淀粉含量则是山血丹最大(76.1±55.52 mg/g),其次为草珊瑚(59.67±54.02 mg/g),狗骨柴最小(13.64±6.33 mg/g);非结构性碳水化合物含量为58.63(±11.03)~159.64(±39.58) mg/g,其中最小的是红枝蒲桃,最大为鸭脚木;可溶性糖/淀粉八角枫最大(5.82±3.29 mg/g),其次是鸭脚木(5.11±1.69 mg/g),黧蒴最小(1.15±0.64 mg/g)(表1)。对15种常见种幼苗根、茎、叶3个不同植物器官的非结构性碳水化合物及其组分含量进行方差分析,结果表明,物种、植物器官和其交互作用对非结构性碳水化合物及其组分含量均存在显著影响(表2)。
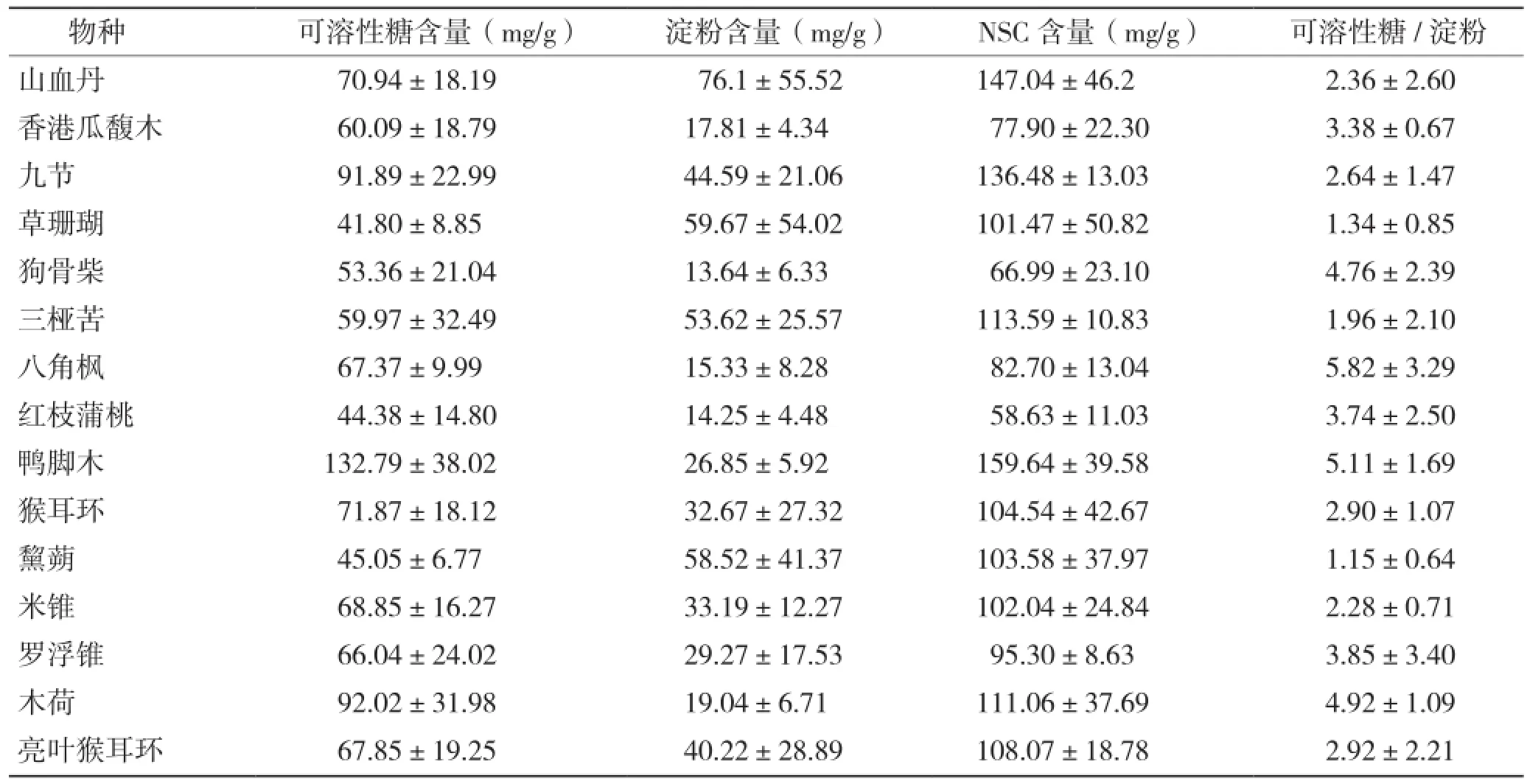
表1 各物种非结构性碳水化合物及其组分含量

表2 物种和植物器官及其交互作用对非结构性碳水化合物及其组分含量的方差分析
2.2 15种幼苗各器官非结构性碳水化合物含量及其分配
2.2.1 植物种间可溶性糖含量及其分配 15种常见种幼苗普遍以植物叶片的可溶性糖含量最多,且不同植物器官间差异性显著,其中只有狗骨柴、猴耳环和米锥3种植物根部可溶性糖含量最高,分别为80.99(±1.83)、91.87(±5.86)、87.37(± 7.92)mg/g,其次为叶,茎的可溶性糖含量最小,其含量分别为36.22(±1)、52.93(±2.54)、55.73 (±3.38)mg/g(图1)。八角枫和黧蒴2种植物可溶性糖含量在根、茎、叶3种植物器官中差异不显著;山血丹、草珊瑚、红枝蒲桃和木荷均为叶片中的可溶性糖含量与根、茎间有显著差异,而根和茎中的可溶性糖含量无显著差异;香港瓜馥木、三桠苦和罗浮锥叶片可溶性糖含量最高,分别为82.72 (±5.84 )、101.74(±4.16)、94.31(±1.87)mg/g,茎次之,分别为55.66(±1.39)、47.47(± 1.86)、64.69(±1.61)mg/g,根部最小,其含量分别为41.89(±1.82)、30.7(±2.67)、39.11(± 0.46)mg/g,且三者间差异性显著;九节和鸭脚木则是叶片和根部可溶性糖含量无显著差异,茎的可溶性糖含量最小(66.74±5.17 mg/g和88.89±2.56mg/g);此外,亮叶猴耳环根部的可溶性糖含量最小(45.38±4.07 mg/g),且与茎和叶片差异性显著(图1)。
2.2.2 植物种间淀粉含量及其分配 从图2可以看出,各植物在根、茎、叶3个不同的植物器官中的淀粉含量存在显著差异。草珊瑚、红枝蒲桃、猴耳环、黧蒴和亮叶猴耳环5种植物根部淀粉含量均比茎、叶高,且差异性显著,而茎、叶间无显著差异;山血丹和罗浮锥根部的淀粉含量最高(141.7±10.26 mg/g和51.11±2.25 mg/g),其次为茎(70.73±5.01 mg/g和25.09±1.01 mg/g),叶片淀粉含量最小(15.87±0.8 mg/g和11.6±0.69 mg/g),且三者间差异显著;狗骨柴、三桠苦和鸭脚木叶片的淀粉含量最小,其含量分别为6.22(± 0.48) 、21.97(±1.63)、22.26(±1.8) mg/g,根和茎的淀粉含量间无显著差异,两者均与叶片的淀粉含量存在显著差异;香港瓜馥木、米锥和木荷叶片淀粉含量最高,其含量分别为22.84(±1.71)、43.2(±5.14)、26.17(±3.92 )mg/g,且与根、茎存在显著差异;此外,九节和八角枫茎部的淀粉含量最高(68.93±8.77 mg/g和25.25±2 mg/g),且与根部、叶片的淀粉含量具有显著差异。
2.2.3 植物种间非结构性碳水化合物含量及其分配 从图3可以看出,植株幼苗的NSC含量在根、茎、叶不同植物器官中差异显著。九节、三桠苦、八角枫和亮叶猴耳环4种植物幼苗NSC在根、茎、叶中差异不显著;香港瓜馥木、红枝蒲桃、罗浮锥和木荷4种植物叶片的NSC含量最高,其含量分别为105.56(±6.45)、71.98(±2.16)、105.91 (±1.18)、154.91(±16.25)mg/g,且与根部和茎差异显著,而根部的NSC和茎的NSC之间无显著差异;山血丹、狗骨柴、猴耳环和黧蒴4种植物幼苗根部的NSC含量最高,其含量分别为202.36 (±18.13)、97.5(±1.37)、154.01(±21)、153.53(±2.68)mg/g,且与叶片和茎的NSC含量存在显著差异;草珊瑚、鸭脚木和米锥的根部NSC含量最高,其含量分别为168.33(±3.08)、182.29 (±5.06)、125.39(±8.79)mg/g,叶片次之,其含量分别为75.69(±4.21)、182.21(±22.3)、106.64(±7.72)mg/g,茎的NSC含量最少,其含量分别为60.38(±2.16)、114.43(±5.68)、74.09(±3.46)mg/g;草珊瑚的NSC含量在根、茎、叶3种植物器官中差异显著,鸭脚木和米锥的根部和叶片的NSC含量无显著差异,且都与茎存在显著差异(图3)。

图1 可溶性糖含量在植物不同器官中的分配差异
2.2.4 植物种间可溶性糖/淀粉及其分配 从图4可以看出,植物幼苗不同器官中的可溶性糖/淀粉存在显著差异,山血丹、三桠苦、八角枫、红枝蒲桃和鸭脚木5种植物的叶片可溶性糖/淀粉比值最高,其值分别为5.81(±0.29)、4.70(±0.47)、9.45(±1.11)、6.46(±1.44)和7.14(±0.38),叶片的可溶性糖/淀粉比值均与根部和茎存在显著差异,且根部和茎间差异不显著;草珊瑚、罗浮锥和亮叶猴耳环3种植物幼苗的叶片可溶性糖/淀粉比值最高,分别为2.20(±0.08)、8.20(±0.61)和5.56(±0.51),茎次之,根部最少,分别为0.28 (±0.03)、0.77(±0.02)和0.62(±0.08),且三者间差异显著;九节和狗骨柴叶片的可溶性糖/淀粉比值最高(4.13±0.31和6.94±0.3),根部次之(2.75±0.54和5.28±1.13),茎最少(1.02± 0.22和2.04±0.26),九节的可溶性糖/淀粉比值在根、茎、叶中差异显著,狗骨柴的叶片和根部的可溶性糖/淀粉比值间无显著差异,且均与茎存在显著差异;香港瓜馥木、猴耳环和黧蒴3种植物幼苗的叶片和茎的可溶性糖/淀粉比值无显著差异,且两者均与根部的可溶性糖/淀粉比值存在显著差异;米锥茎的可溶性糖/淀粉比值最高(3.03± 0.18),根部次之(3.03±0.18),叶片最少(1.5± 0.12),三者间存在显著差异;木荷的根部可溶性糖/淀粉比值最高(5.89±0.43),叶片次之(5.03 ±0.52),茎的可溶性糖/淀粉比值最少(3.84± 0.28),且根部与茎两者之间存在显著差异。

图2 植物不同器官中淀粉含量的分配差异

图3 非结构性碳水化合物在植物体内不同器官中的分配差异

图4 可溶性糖/淀粉在植物体内不同器官中的分配差异
2.3 不同生长型树种非结构性碳水化合物及其组分含量的比较
我们将15种植物分常绿乔木和常绿灌木进一步探讨。常绿灌木根和茎的淀粉和NSC含量比常绿乔木高,但常绿灌木的可溶性糖含量在根、茎、叶3个不同的植物器官中均比常绿乔木低。乔木树种幼苗和灌木树种幼苗的NSC及其组分含量在植物根部和叶片中无显著差异,乔木和灌木树种幼苗茎的可溶性糖与NSC含量也无显著差异,但其淀粉含量和可溶性糖/淀粉在茎中的分布具有显著差异(图5)。无论是乔木还是灌木均是叶片的可溶性糖含量最高,其含量分别为88.14(±7.98)、77.34(±5.22)mg/g,其次是根部,其含量分别为71.18(±7.99)、60.83(±5.93)mg/g,茎的可溶性糖含量最少,只有61.5(±3.22 )、52.71(±2.91)mg/g。灌木的叶片可溶性糖含量与根部和茎均具有显著差异,且茎和根部两者之间无显著差异;乔木的根部和叶片的可溶性糖含量间无显著差异,叶片和茎的可溶性糖含量间存在显著差异(图5)。
乔木和灌木幼苗的根部淀粉含量最高,其含量分别为50.86(±6.74)、62.26(±11.59)mg/g,茎次之,叶片最少,其含量分别为21.99(±2.22)、17.58(±1.71)mg/g。灌木的根、茎、叶3种不同的植物器官间的淀粉含量存在显著差异;乔木的根部淀粉含量与茎和叶片间具有显著差异,但茎和叶片两者间无显著差异。乔木和灌木幼苗根部的NSC含量最高,其含量分别为122.04(±8.68)、123.09 (±10.87)mg/g,叶片次之,茎最少,其含量分别为83.91(±4.16)、93.19(±7.06)mg/g。灌木的根部NSC含量与叶片和茎的NSC含量间均存在显著差异,但叶片和茎两者之间无显著差异;乔木的根部和叶片NSC含量间无显著差异,且均与茎的NSC含量存在显著差异(图5)。
灌木的叶片可溶性糖/淀粉最高(5.27± 0.52),根部次之(2.46±0.51),茎最小(1.81± 0.23)。叶片的可溶性糖/淀粉与根部和茎均具有显著差异,根部和茎两者间差异不显著;乔木叶片的可溶性糖/淀粉最高(4.89±0.52),且与茎和根部存在显著差异,茎次之(2.96±0.16),根部的可溶性糖/淀粉最小(2.23±0.4),并且根部和茎两者的可溶性糖/淀粉间无显著差异(图5)。
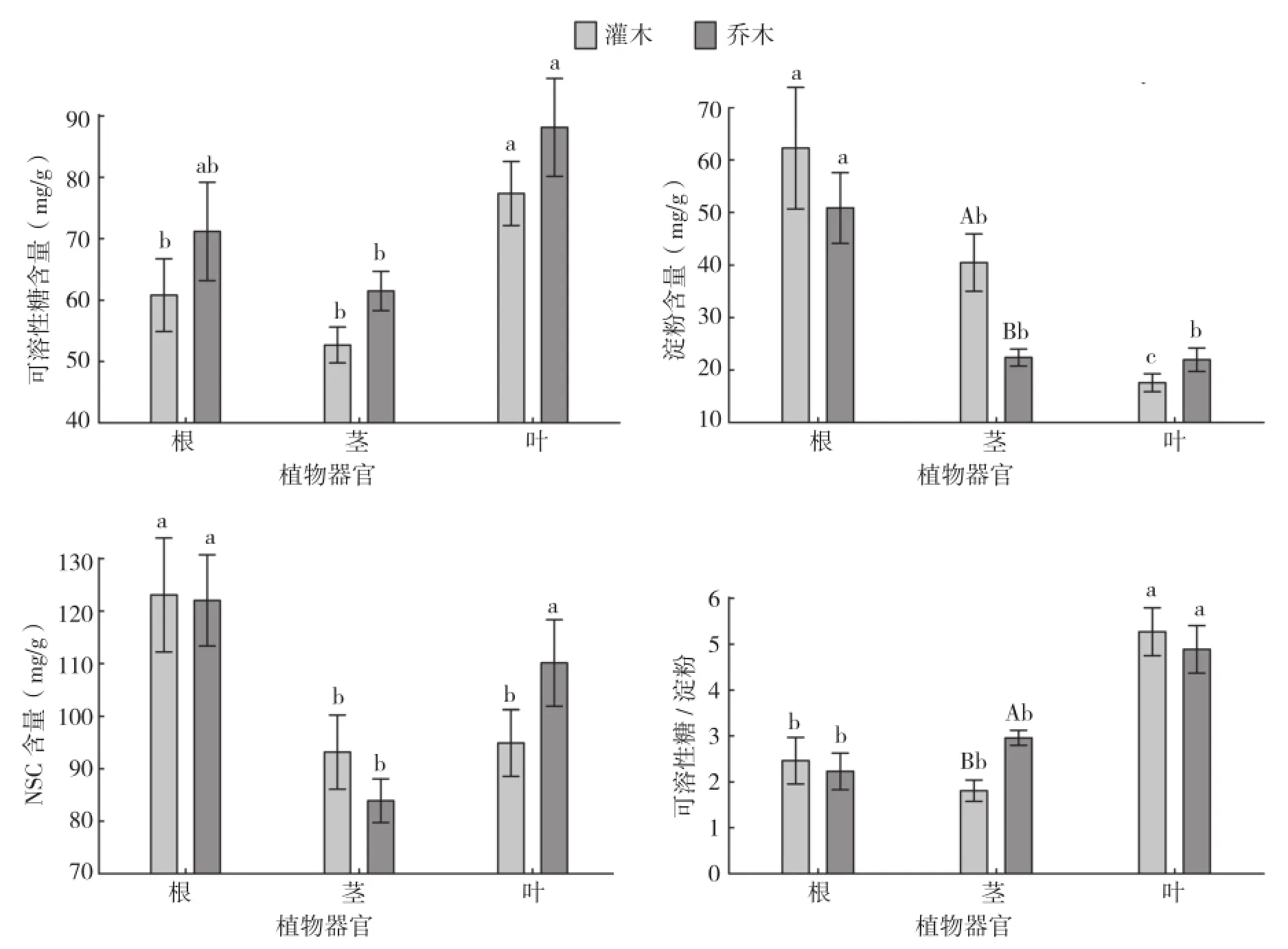
图5 不同生长型植物幼苗NSC及其组分含量在根、茎、叶中的分配差异
3 结论与讨论
本研究发现,15种常见种幼苗非结构性碳水化合物及其组分含量在植物种间均存在差异,这可能与植物本身的遗传特性和外界的环境条件有关[17]。另外,15中常见种幼苗植株茎的NSC含量为88.24 mg/g,根的NSC含量为122.53 mg/g,均低于赵镭等[18]对浙江天童幼苗报道的常绿种植株茎的NSC含量(148.7 mg/g)和根的NSC含量(408.52 mg/g),造成两者差异的原因可能是:研究区域不同,外界环境条件存在差异,以及所选择的物种不同。同时,不同生长型的植物非结构性碳水化合物储存也存在差异。常绿灌木根的淀粉和NSC含量比常绿乔木高,而常绿灌木的可溶性糖含量在根、茎、叶3个植物器官中均比常绿乔木低,但无显著差异;常绿乔木的茎可溶性糖/淀粉显著高于灌木,淀粉含量显著低于灌木。可见,常绿乔木和灌木根据各自的生长特点形成了有利于其生存的NSC储存机制,乔木在竞争中争夺阳光的能力较强,可溶性糖含量较高,有利于其争夺更多的生存空间;灌木多生活在荫蔽的环境中,光合作用效能较乔木弱,可溶性糖含量较低,但根和茎中储存较多的NSC可以转化为可溶性糖,用于其生长发育,使其在竞争中拥有更大的生存机会。
植物中储存的NSC及其组分含量因树种而异,鸭脚木的可溶性糖(132.79±38.02 mg/g)和NSC含量(159.64±39.58 mg/g)均为最高,山血丹的淀粉含量最高(76.1±55.52 mg/g),八角枫的可溶性糖/淀粉含量最大(5.82±3.29),草珊瑚的可溶性糖含量最小(41.8±8.85 mg/g),而淀粉含量较高(59.67±54.02 mg/g)。同时植物NSC及其组分含量在不同的植物器官根、茎、叶中差异显著,植株幼苗叶片的可溶性糖和可溶性糖/淀粉比值较高,根部的淀粉和NSC含量较高,说明植株叶片和根部是主要的碳储存器官。刘骏等[9]认为,厚壁毛竹地下器官的淀粉含量明显比地上器官高,叶片的可溶性糖含量较高;于丽敏等[19]认为叶和根是植物生命代谢最旺盛的器官,叶的可溶性糖含量较高,根的NSC含量较高,本研究的结果与前人的研究一致。NSC及其组分含量在植物种内不同植物器官中的分配格局体现了植物对所生活环境条件的适应,可溶性糖和淀粉互相补充从而使NSC总量保持不变,植物可以通过调节可溶性糖/淀粉比值来适应环境的变化,为其争取更大的生存机会。有关研究表明,植物在面对水分胁迫时,主要表现为可溶性糖含量增加和淀粉含量降低[20];叶片可溶性糖与淀粉比值随海拔高度的升高而增大[21]。Li等[22]对喜马拉雅林线树种的报道指出可溶性糖/淀粉比值接近3则可以保证高海拔地区植物顺利过冬。而本研究中15种常绿阔叶林常见种幼苗的可溶性糖/淀粉比值为1.15~5.82,且普遍表现为叶片的可溶性糖/淀粉比值最大。
幼苗是森林群落更新和植被恢复的关键,处于幼苗期的植物对环境较为敏感。不同的植物具有不同的利用和分配资源的能力,常绿乔木和灌木幼苗采取不同的碳储存和分配机制,且都向着有利于其生存的方向发展,反映了其对环境的适应策略。深入研究NSC的储存和分配,对揭示环境条件对植物幼苗物质积累与分配的影响具有重要作用,为生态系统植被保护,造林营林提供更多的理论依据。在建设和管理森林公园森林生态系统时,应结合植被自身的遗传特性和实际的生态环境,深入了解植物群落应对环境的策略,充分发挥森林群落的生态效能。
[1]王延平,许坛,朱婉芮,等.杨树细根碳、氮含量的季节动态及代际差异[J].应用生态学报,2015,26 (11):3268-3276.
[2]蒋思思,魏丽萍,杨松,等.不同种源油松幼苗的光合色素和非结构性碳水化合物对模拟氮沉降的短期响应[J].生态学报,2015,35(21):7061-7070.
[3]郑云普,王贺新,娄鑫,等.木本植物非结构性碳水化合物变化及其影响因子研究进展[J].应用生态学报,2014,25(4):1188-1196.
[4]于贵瑞,李娜妮,何念鹏.中国东北典型森林生态系统植物叶片非结构性碳水化合物研究[J].生态学报,2016,36(2):1-9.
[5]Yee D,Tissue D T.Relationships between non structural carbohydrate concentration and flowering in a subtropical herb,Heliconia caribaea(Heliconiaceae)[J].Caribbean Journal of Science,2005,41(2):243-249.
[6]Palacio S,Millard P,Maestro M,et al.Non-structural carbohydrates and nitrogen dynamics in mediterranean sub-shrubs:an analysis of the functional role of overwintering leaves[J].Plant Biology,2007,9(1):49-58.
[7]Bansal S,Germino M J.Temporal variation of nonstructural carbohydrates in montane conifers:similarities and differences among developmental stages,species and environmental conditions[J].Tree Physiology,2009,29(4):574-575.
[8]Latt C R,Nair P,Kang B T.Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate[J].Forest Ecology and Management,2001,146(1):145-158.
[9]刘骏,杨清培,杨光耀,等.厚壁毛竹非结构性碳水化合物分配格局[J].江西农业大学学报,2011,33 (5):924-928.
[10]周云龙.广州市流溪河森林公园植物区系的初步研究[J].热带亚热带植物学报,1994,2(2):31-40.
[11]郭志华,肖文发,周云龙.广州流溪河常绿阔叶林的植物区系地理分析[J].林业科学研究,2003,16 (5):622-627.
[12]苏开君,王光,马红岩,等.流溪河小流域针阔混交林林冠降雨截留模型研究[J].中南林业科技大学学报(自然科学版),2007,27(1):60-63.
[13]王冬云,张卓文,苏开君,等.广州流溪河流域毛竹林的水文生态效应[J].浙江林学院学报,2008,25(1):37-41.
[14]曾震军,吴汉.流溪河林场森林资源调查与分析[J].广东林业科技,2009,25(1):77-82.
[15]Wang F,Sanz A,Brenner M L,et al.Sucrose synthase,starch accumulation,and tomato fruit sink strength[J].Plant Physiology,1993,101(1):321-327.
[16]Buysse J,Merckx R.An improved colorimetric method to quantify sugar content of plant tissue[J].Journal of Experimental Botany,1993,44(10):1627-1629.
[17]李娜,孙涛,毛子军.长期极端高温胁迫对樟子松幼苗生物量及非结构性碳水化合物的影响[J].植物研究,2014,34(2):212-218.
[18]赵镭,杨海波,王达力,等.浙江天童常见种幼苗的光合特性及非结构性碳水化合物储存[J].华东师范大学学报(自然科学版),2011(4):35-44.
[19]于丽敏,王传宽,王兴昌.三种温带树种非结构性碳水化合物的分配[J].植物生态学报,2011,35 (12):1245-1255.
[20]杜尧,韩轶,王传宽.干旱对兴安落叶松枝叶非结构性碳水化合物的影响[J].生态学报,2014,34 (21):6090-6100.
[21]周永斌,吴栋栋,于大炮,等.长白山不同海拔岳桦非结构碳水化合物含量的变化[J].植物生态学报,2009,33(1):118-124.
[22]Li M,Xiao W,Wang S,et al.Mobile carbohydrates in Himalayan treeline trees I.Evidence for carbon gain limitation but not for growth limitation[J].Tree Physiology,2008,28(8):1287-1296.
(责任编辑 白雪娜)
Study on non-structural carbohydrates of common species seedlings of evergreen broad-leaved forest in Liuxihe
HUANG Run-xia1,JIA Xiao-rong1,WU Hui-jun2,RAO Shu-pei1,LUO Wan-ying1,LI Ling1,LIN Shu-hang1
(1.College of Forestry and Landscape Architecture,South China Agricultural University,Guangzhou 510642,China;2.Lechang Forest Farm of Guangdong,Shaoguan 512219,China)
In order to study the interspecific differences and allocation pattern of non-structural carbohydrate (NSC)among roots,stems and leaves and its component about common species seedlings of evergreen broadleaved forest,15 common species seedlings of evergreen broadleaved forest in Liuxihe Forest Park were investigated.The concentration of soluble sugar,starch,NSC and the ratio of soluble sugar and starch of roots,stems and leaves were studied.The results showed that significant differences were found among species and significant interaction was existed among species and plant organs (P< 0.05).15 seedlings of common species generally showed that the content of soluble sugar and the ratio of soluble sugar and starch of leaves were the highest,while the starch and NSC content of roots were the biggest.There was no significant difference of NSC and its component in roots and leaves of tree seedling and shrub seedlings.There was no significant difference of soluble sugar and NSC content in stems of tree and shrub seedlings.But the ratio of soluble sugar and starch in stems of tree was significant higher than that of shrub,while its starch content was significant lower than shrub.Interspecific differences and allocation pattern of non-structural carbohydrate for seedlings of common species in evergreen broadleaved forest reflected that leaves and roots of plants seedlings were the synthesis and storage structure of NSC,whichcould provide nutrients for the growth and development of plants.Also,it reflected their different survival strategies to deal with the environment.
seedlings;NSC;evergreen broadleaved forest;Liuxihe
S718.43
A
1004-874X(2016)06-0081-09
10.16768/j.issn.1004-874X.2016.06.015
2016-02-14
广东省林业科技创新项目(2016K JCX027);广东省科技计划项目(2013B020305008); 2014年华南农业大学大学生科技创新项目(LX010)
黄润霞(1993 -),女,在读本科生,E-mail:runxiah@163.com
贾小容(1978-),女,博士,副教授,E-mail:xiaorongj@scau.edu.cn
