基于结构方程的滇东石漠化地区土壤理化性质与酶活性研究
2016-07-26舒树淼赵洋毅胡慧蓉熊好琴
舒树淼, 赵洋毅, 胡慧蓉, 熊好琴, 段 旭
(1.西南林业大学 环境科学与工程学院, 云南 昆明 650224; 2.云南玉溪森林生态系统国家定位观测研究站, 云南 新平 653400; 3.西南林业大学 林学院, 云南 昆明 650224)
基于结构方程的滇东石漠化地区土壤理化性质与酶活性研究
舒树淼1,2, 赵洋毅1,2, 胡慧蓉1, 熊好琴1,2, 段 旭2,3
(1.西南林业大学 环境科学与工程学院, 云南 昆明 650224; 2.云南玉溪森林生态系统国家定位观测研究站, 云南 新平 653400; 3.西南林业大学 林学院, 云南 昆明 650224)
摘要:[目的] 明确滇东石漠化地区不同恢复模式效果差异以及养分对酶的影响关系,为石漠化地区恢复措施提供理论依据。[方法] 以土壤酶活性为恢复效果的衡量指标,采取基于模糊数学的土壤质量评价办法,得到不同模式下土壤酶指数,用以比较恢复模式的效果。基于土壤理化性质与酶活性相关性分析的结果,采取结构方程模型量化土壤理化环境和酶活性环境,进而分析不同土壤理化环境对不同酶环境的影响。 [结果] 酶活性大小排序为:灌草模式>针阔混交林模式>阔叶林模式>针叶林模式>裸地;由结构方程模型和有机质、全磷估计出的养分子环境对由脲酶活性、过氧化氢酶、多酚氧化酶活性构成的酶活性环境存在显著正影响。 [结论] 灌草模式和针阔混交林模式对石漠化恢复有较好作用。养分环境对酶活性环境的影响系数为0.87。
关键词:喀斯特; 石漠化; 植被恢复模式; 土壤养分; 酶活性; 结构方程模型; 滇东
文献参数: 舒树淼, 赵洋毅, 胡慧蓉, 等.基于结构方程的滇东石漠化地区土壤理化性质与酶活性研究[J].水土保持通报,2016,36(3):338-345.DOI:10.13961/j.cnki.stbctb.2016.03.058
目前对石漠化的治理一般采用生态恢复等办法,而生态恢复必须涉及土壤这一关键因子。这就包括了土壤的各项性质。其中土壤酶被认为是可以综合反映土壤肥力的指示指标[1]。这是因为土壤酶主要来源于植物残体、植物根系分泌以及土壤微生物作用[2],对土壤生态系统中养分的转化和利用有着不可替代的作用[3],具有受综合因素影响的特征。同时有研究表明,几乎所有土壤退化都伴随着不同程度土壤酶活性的丧失[4]。因此对土壤酶的研究十分必要。在不同植被模式下酶活性各有不同。如Xing等[5]的研究表明土壤脲酶和蛋白酶在杉木针叶林中活性显著低于阔叶石楠和混交林。Ushio等[6]则指出酚氧化酶在不同植被类型之间差异不显著。而一般认为土壤酶活性与土壤养分存在一定相关关系。如脲酶和磷酸酶活性与C,P,N等养分循环就有密切关系[7]。同时酶活性也受到土壤结构、pH值、微生物、微量元素等因子的影响。显然在不同植被类型中,影响酶活性的因子非常复杂。目前就理化因子与土壤酶活性关系的研究方法中,如路径分析、主成分提取等并不能同时处理多个因变量与自变量的联系[8]。这样在一定程度上就偏离了统计因果。所以若能通过某种恰当的分析,在已有的信息情况下无偏地揭示酶活性与环境的因果关系是十分有必要的。结构方程模型不仅可以处理多个自变量与因变量,还具有直观描述隐变量等特点,为探明因果关系提供了有力方法。
本文选择滇东石漠化地区典型不同植被恢复模式(以裸地为对照),分别获得过氧化氢酶、蔗糖酶、脲酶、多酚氧化酶、蛋白酶、淀粉酶在不同土壤层次的活性。在获得不同植被类型酶活性大小结果的同时,采用结构方程模型分析理化因子和酶活性之间的路径关系,以期找到在不同植被模式下影响酶活性的因果联系,并将这种联系定量化,从而对滇东地区石漠化生态环境保护和石漠化治理提供基础理论依据。
1研究区概况
研究区位于云南省东部曲靖市(103°52′E,25°45′N),海拔1 150~2 750 m,具有山地季风半湿润气特候征,年平均气温13.5 ℃,年均降水量1 300 mm,年平均蒸发量831.3 mm年均日照1 820 h。土壤类型主要为红、黄壤土为主,植被属于北亚热带中山常绿针阔混交林带。
研究地位于云南省典型的喀斯特地貌分布区内(介于103°57′—104°19′E,25°03′—25°47′N),该区石漠化土地较多,水土流失严重,石漠化面积占全区的17.23%,占全区喀斯特地貌面积的27.45%。其中,轻度石漠化占全区面积的10.04%,中度占16.51%,强度占1.84%,极强度占0.94%。试验地选取滇东高原典型石漠化地段,区域内主要以中轻度石漠化为主,以4种植被恢复方式进行治理,分别为针阔混交林(云南松Pinusyunnanensis×华山松Pinusarmandii×麻栎Quercusacutissima)、针叶林(华山松)、阔叶林(麻栎)及灌草植被(椴叶山麻杆Alchorneatiliifolia、盐肤木Rhuschinensis、白茅Imperatacylindrica、紫茎泽兰等Ageratinaadenophora)模式(表1)。
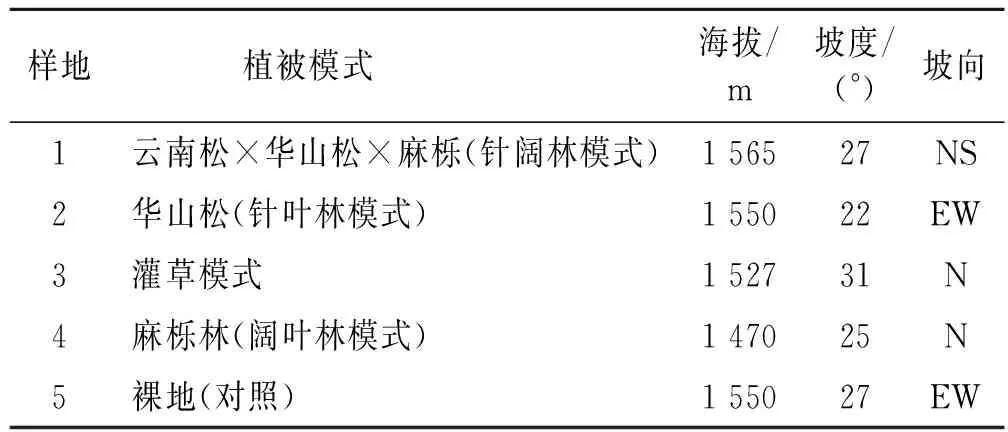
表1 不同植被模式样地基本情况
2研究方法
2.1土壤理化性质测定

2.2数据处理
(1) 土壤酶指数与统计分析。因土壤酶种类较多,各作用不同,且存在信息重叠[9],而单一的酶指标并不能全面反映整体酶活性。故使用土壤酶指数(SEI)判别各植被模式土壤酶因子综合作用。土壤酶的变化具有连续性质,因而可先采取描述连续性质的隶属度函数,并从主成分因子负荷量的正负性确定隶属度函数分布的升降型。公式如下:
SEI(xi)=(xij-ximin)/(ximax-ximin)
(1)
SEI(xi)=(ximax-xij)/(ximax-ximin)
(2)
式中:xij——土壤酶活性的值(ml/g);ximax和ximin——土壤酶活性最大最小值(ml/g)。土壤多酚氧化酶为降型分布,其他酶为升型分布。
因各个酶活性指标的单位不同,故可通过计算主成分贡献率和累计贡献率,然后利用主成分分析因子载荷得到各因子权重。
Wi=Ci/C
(3)
式中:Wi——各酶指标权重;Ci——各酶公因子方法;C——所有酶的公因子方差之和。于是加权得到土壤酶指数(SEI),以客观、全面地反映样地整体酶活性。
(4)
(2) 结构方程模型。结构方程模式是广泛应用于心理学、经济学、社会学、行为科学等领域的因果检验模型[8]。是一般线性模型的扩展。如路径分析、因素分析、多元方差分析、多元回归等都可以看做结构方程模型的特列[8]。相比于结构方程模型上述方法只能检验自变量和因变量的单一关系[8,10]。而结构方程模型不仅综合了已有的线性分析方法,还能对变量关系进行验证分析,并拥有严格的统计假设检验办法,且容纳了变量存在的测量误差,因此结构方程模型具备了一般线性分析不具有的能力,如为分析那些不可直接测量的变量(隐变量)之间的结构关系提供了可能[8]。本研究中理化环境和酶活性环境均是不可直接测量的变量。同时一般认为结构方程模型适用于较大样本(n≥100),路径关系较为复杂的结构分析中。然而结构方程模型是检验模型,必须基于一定的理论基础。故也可以适用于有明确理论依据的简单结构。在本研究中,理论基础为理化因子对酶活性存在影响。已被大量研究已经证明,即不论何种植被模式下某些养分和酶之间都存在极显著或显著相关[11-13],表明了养分和酶具有一定固有联系。事实上对于土壤生态系统而言,土壤因子之间都存在着错综复杂的联系。如脲酶同有机质可形成酶—腐殖质化合物[14],有机质对脲酶的稳定和活性具有一定作用。同时脲酶主要聚集在微团聚体上,相当于土壤粒经的黏粒部分。其活性随粒径增大有下降趋势[15]。而有机质又影响土壤结构的形成[16]。可见对于因子联系较多的系统,若将其中一个因子作为研究对象,探讨其它因子对它的影响是十分复杂的。因此为了描述这种影响,可以在逻辑上设存在一个“环境”,“环境”由不同因子构成。本文中把理化因子视为构成“土壤环境”的因子,进而影响“酶活性环境”,而“酶活性环境”则直接影响不同酶活性。因上述“土壤环境”和“酶活性环境”都不能直接观察,于是具备了结构方程模型中隐变量的特征。
根据结构方程模型,隐变量是指那些不能被直接观测的变量,往往通过显变量对其进行估计。在土壤生态系统中,如各养分指标、物理指标都是可被直接测量的指标,通过对这些显性指标的把握可以评估出土壤整体环境。因而符合结构方程模型的基本思想。结构方程模型分为测量模型部分和结构模型部分。测量模型可衡量隐变量与外显变量之间的关系。
X=∧xξ+δ
(5)
Y=∧yη+ε
(6)
式中:X——外生显性变量向量;Y——内生显性变量向量; ∧x和∧y——指标变量(X,Y)的因素负荷量;δ,ε——外生显性变量与内生显性变量的测量误差;ξ——外生潜在变量;η——内生潜在变量。结构模型可反映各潜在变量之间的关系。
η=Bη+Γξ+ζ
(7)
式中:B——内生潜在变量之间关系的结构系数矩阵; Γ——内生潜在变量与外生潜在变量之间关系的结构系数矩阵;ζ——结构模型中干扰因素或残差值。根据此模型和最大似然法便可建立结构方程模型路径图。在这里将养分因子和物理因子定义为外生显变量;它们构成的环境定义为外生隐变量;受到综合环境影响的酶活性环境定义为内生隐变量;各个酶因子的活性定义为内生显变量。使用SPSS和AMOS进行结构方程路径分析。
3结果与分析
3.1不同植被模式土壤酶指数
表2为各个样地不同层次土壤平均酶活性情况。可知酶活性几乎都随着土壤深度的增加而增加。大部分酶活性变化明显,同时也存在酶在某些样地中活性随深度变化无明显差别。如脲酶活性在样地1中变化明显,但在样地3,4的某些层中酶活性都十分相近。

表2 不同植被模式样地土壤酶活性
由表2可以看出,不同酶活性在不同样地土壤中此消彼长,为了全面综合地衡量每个样地中酶活性的大小,故采用土壤酶指数(SEI)作为衡量标准。结果如图1—2所示。

注:不同字母表示差异显著(p<0.05)。下同。
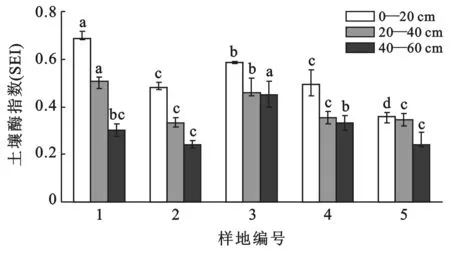
图2 各植被模式样地不同深度土壤酶指数(SEI)
各样地酶指数分别为0.498,0.352,0.499,0.392,0.315。可知样地1(针阔林模式)、样地3(灌草模式)、样地4(阔叶林模式)的土壤酶指数明显大于裸地5。而样地2(针叶林模式)和样裸地5最为接近。即:灌草模式>针阔林模式>阔叶林模式>针叶林模式>裸地。有研究表明在喀斯特地区中酶活性随植物恢复过程逐渐提高[17],同时也有研究表明微生物数量从高到低存在:灌草丛>常绿阔叶林>针阔混交林>针叶林的大小序列,而酶活性与之类似[18]。可见对于不同植被模式,酶活性大小并没有确定序列。这是因为仅从植被模式上并不能概括酶存在的具体环境。图2揭示了不同深度酶活性大小。显然对于乔木下的土壤而言,其酶活性随深度变化明显,这表明了土壤环境随深度改变明显。这可能和植物根系有关,即乔木根的活动深度较灌草强,故明显改变了深层土壤的水热环境,进而改变了酶存在环境。因麻栎是浅根系植物,故样地4在这点上表现得不如有针叶植物的模式林明显。另外裸地土壤中层酶活性较大,这表明裸地衰退的时间并不长,土壤中依然存在适宜酶存在的环境。
3.2土壤酶活性与理化指标的相关关系
表3列出了不同植被模式下土各土层典型理化指标的平均值。一般认为酶活性和这些指标有关,而这些指标也能在一定程度上反应土壤肥力。其中直径在0.5~3 mm大小的团聚体是决定土壤肥力的主要因素之一[19],为了便于操作本研究适当扩大到0.25~3 mm,并能包含更多的土壤结构信息。可以看到绝大所属理化指标随着土壤深度的增加表现出下降趋势。pH值正好相反。表4为各理化因子和6种酶person相关性分析。由结果可知含水率、团聚体和酶的相关性均不显著,而蛋白酶也不和任何理化因子表现出相关显著性。其次pH值仅与脲酶和淀粉酶显著负相关。而其余指标均相互表现出显著、极显著相关性。这说明在石漠化地区的不同植被恢复模式下,即使酶存在的环境千差万别,酶活性也仍然和某些理化因子存在着一定固有联系。且这种联系因石漠化地区从“空白”状态开始积累土壤的原因变得特别明显。事实上这些因子之间也并不孤立,同时也是相互影响后最终表现出确定状态。因此可以通过个别理化指标对土壤环境做出整体估计。同理,某些酶活性也能对酶整体活性环境做出整体估计,于是这为结构方程模型的应用提供了理论依据。

表3 不同植被模式下土壤理化指标
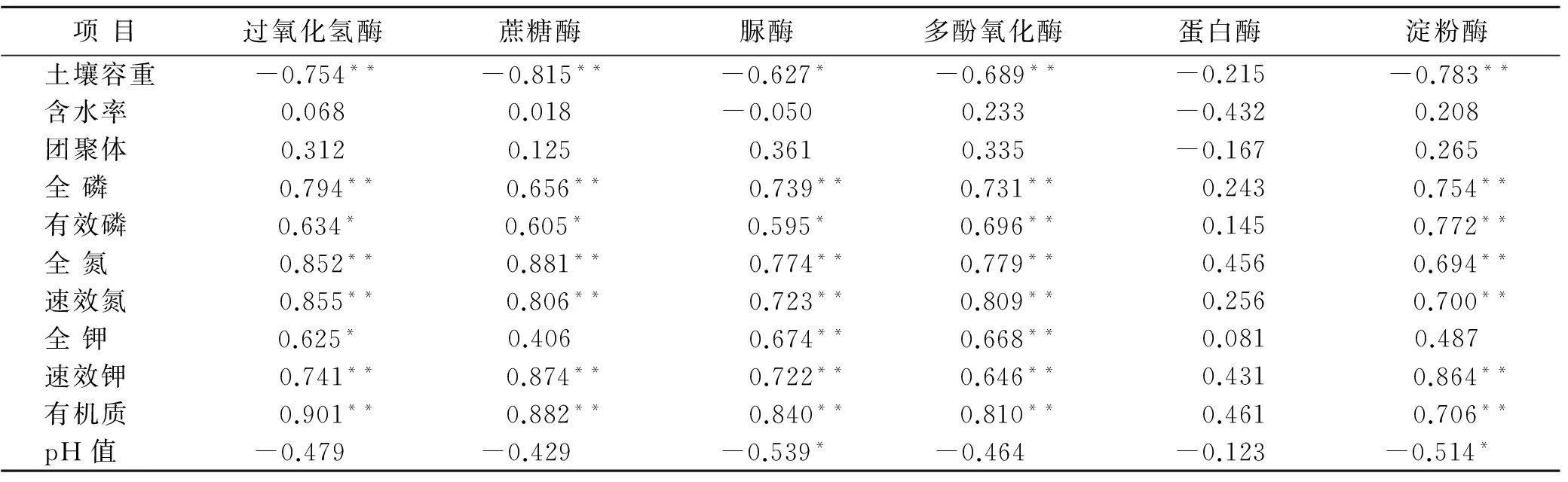
表4 土壤酶活性与土壤理化性质相关关系
注:**表示极显著相关(p<0.01);*表示显著相关(p<0.05),其中n=105。
3.3土壤理化因子与酶活性的结构方程模型
本研究所采用的结构方程模型的一般形式为图3所示。其中x表示可被观测到的理化指标,ξ称为外生隐变量表示影响酶活性的外界环境。η则为内生隐变量表示酶活性环境,由内生显变量y估计。δ,ε,ζ均为对应残差。结合上文结果选择相关显著的理化因子和酶因子通过结构方程模型来分析理化因子对酶活性影响大小。因土壤单独某酶活性对土壤整体环境的响应各不相同。如有研究表明全磷含量是影响多酚氧化酶最主要因子[20],而速效钾是影响蔗糖酶活性的最主要因素[21]。故对影响酶活性的土壤环境不能整体考虑。

图3 结构方程模型一般形式
即土壤环境中包含了影响某类酶活性的“子环境”,故分析“子环境”和某类酶活性环境的关系才能准确可靠。考虑到某些指标存在共线性影响,如有机质和全氮含量普遍存在线性关系[22],故仅考虑有机质。同时结构方程模型要求至少2个外显变量才能估计出隐变量。图4—5为土壤理化因子与土壤酶活性的最优结构方程模型。其中使用有机质(x1)、全磷(x2)、全钾(x3)、速效钾(x4)等指标对养分环境进行估计;容重(x)表示了土壤密度,在一定程度上概括了土壤物理环境,对酶活性环境起到作用且和养分环境存在相关关系。而脲酶活性(y1)、过氧化氢酶活性(y2)、多酚氧化酶活性(y3)、蔗糖酶活性(y4)、淀粉酶活性(y5)在不同“子环境”下表现不同活性,本文分成了两类。根据结构方程模型检验指标模型拟合良好,χ2/df显著度均大于0.05,接受虚无假设。表5为具体拟合结果。

图4 结构模型方程模型1

图5 结构模型方程模型2
由结构方程模型拟合结果可知,对于脲酶、过氧化氢酶、多酚氧化酶来说用有机质和全磷来估计影响上述酶活性的养分环境是可行的。其中有机质和全磷对养分环境的估计都是极显著的(p<0.01)。因子载荷量分别为0.98,0.87这意味着有机质和全磷受该种养分环境的影响较大。而3种酶对酶活性环境估计也都极显著(p<0.01),因子载荷量分别为0.92,0.99,0.91表明3种酶活性受该种酶活性环境的影响也较大。养分环境对酶活性环境的影响系数为0.872(p<0.01),较大影响的同时也极为显著。土壤容重对酶活性环境的路径系数为-0.700(p=0.742),并不显著。因此对上述酶活性的影响主要来源于由有机质和全磷估计出的养分环境,为促进作用。同理在图2中速效钾对养分环境的估计是极显著的(p<0.01),且完全受养分环境的影响。而全钾对养分环境的估计为显著(p<0.05),受养分环境的影响一般。酶活性环境对蔗糖酶的影响为极显著(p<0.01),对淀粉酶活性的影响为显著(p<0.05),同时受酶活性环境的影响都较大。然而进一步分析表明养分环境对酶活性环境的影响虽极大,影响系数为0.99,却并不显著(p=0.408)。因此使用全钾和速效钾估计养分环境还不够准确,这表明尚需要更多指标对模型进行调整。同时上述模型中土壤容重的作用都并不明显。故土壤容重并非是直接影响酶活性环境的原因。仅与养分环境表现出较大的负相关性,且模型1中为显著负相关,模型2中为非显著负相关。

表5 结构方程模型拟合结果检验
4讨论与结论
4.1讨 论
本文对滇东石漠化地区不同植被恢复模式下的土壤酶进行了分析。通过研究发现在灌草模式和针阔混交林模式下,土壤酶指数极为相近,这说明即使各土壤酶活性不同,但经过某种配比“贡献”后,两种植被模式下的综合土壤酶活性几乎相同,这与余娜等[23]对沙生植被土壤酶活性中踏浪和沙裸地的研究情况相似。若把各个土壤酶指标看作具体的观察变量,土壤酶指数看作隐变量,则这个由观察变量计算获得隐变量的过程体现了一种由外到内的方法,这正是结构方程模型的重要思想[10,21]。即:隐变量的大小虽然无法直接获得,但最终可通过若干可测外显变量进而获得隐变量,特别是在进行因果讨论的时候,当原因和结果都不能直接测量时,结构方程模型为这种讨论提供了严格准确的数学检验依据[24]。在本文中,土壤环境是作为影响土壤酶活性的原因。而这种土壤环境是一种“子环境”,存在于不同植被模式下。但是这种土壤环境并不能被直接观测,必须通过其他土壤因子来间接观测,因而符合隐变量的思想。本文主要用养分环境来代替土壤环境。这是因为对于滇东石漠化恢复地区来说,各个植被恢复模式都有着相同的恢复本底。这种情况下,养分环境与其他因素存在较为紧密的联系,所以养分环境的不同也就包含了这种差异。文中的相关性结果则符合了这一判断。事实上这与通过土壤因子来监测土壤肥力、土壤环境变化的思路相一致[25-26]。
笔者基于结构方程模型,通过极大似然法估计出了各个路径系数,借助土壤环境影响酶活性环境的因果关系,定量地得出土壤“子环境”对某类“酶活性环境”的影响大小。从而讨论土壤理化因子与酶活性的联系。许多研究表明[20,27-29],有机质、全氮、全磷是影响脲酶的主要因素,全氮是影响过氧化氢酶的直接原因,全磷含量是影响多酚氧化酶和过氧化氢酶活性的最主要因子。本文通过结构方程模型1进一步验证了脲酶、过氧化氢酶、多酚氧化酶活性的变化可为一类,而蔗糖酶、淀粉酶的活性变化可为一类;也有研究认为速效钾是影响蔗糖酶活性的最主要因素[21],在模型2中得到了体现。另外,本文并没有直接探讨各理化因子对某类具体酶活性的作用。而是通过建立隐变量之间的联系,简便直观地解析了多因子复杂耦合现象。如测定的指标里没有直接考虑温度、土壤微生物等影响因子,将它们作为了可被其他因子估计的隐变量。
结构方程模型在自然科学上应用还很少。目前有用于森林生态方面的研究[10,30]。结构方程模型被看作是探索因果理论的工具模型和检验模型,关于使用该方法研究土壤因子对土壤酶活性的影响,因参考的文献较少,文中尚有不足,而在该方法上还有待于做进一步的研究和推广。
4.2结 论
本文对滇东石漠化地区不同植被模式的样地土壤酶活性进行了研究。过氧化氢酶、蔗糖酶、脲酶、多酚氧化酶、淀粉酶与养分因子存在极显著或显著正相关,而土壤容重与上述酶活性表现出极显著负相关。不同植被模式下,酶指数大小为:灌草模式>针阔混交林模式>阔叶林模式>针叶林模式>裸地,其中灌草模式和针阔混交林模式的酶指数的大小极为相近,分别为0.499,0.498,并无明显差别。另外,酶活性在各个植被模式中,随土壤深度的变化各不相同。华山松作为深根系植物可能是对这种差别影响较大的原因。
结构方程模型分析表明即使是对于不同样地而言,由有机质和全磷估计出的养分子环境对由脲酶活性、过氧化氢酶、多酚氧化酶活性构成的酶活性环境存在显著正影响,影响系数为0.87。而由全钾、速效钾估计出的养分子环境对由蔗糖酶活性、淀粉酶活性构成的酶活性环境的影响系数为0.99,却并不显著。土壤容重虽然与酶活性存在显著相关关系,但在结构方程模型中仅与养分环境存在显著负相关,对酶活性环境影响较小,也并不显著。这表明本研究中单纯的土壤容重并不能直接的作为影响酶活性的原因。从因子载荷来看,有机质和全磷、全钾和速效钾对各自养分环境的估计都有主要显著作用。而脲酶活性、过氧化氢酶活性、多酚氧化酶活性变化很可能受土壤中同一土壤环境原因的影响,且这些酶的活性变化可为一类。即酶活性环境变化1时,则3种酶活性分别变化:0.92,0.99,0.91。
[参考文献]
[1]张咏梅,周国逸,吴宁.土壤酶学的研究进展[J].热带亚热带植物学报,2004,12(1):83-90.
[2]关松荫,张德生,张志明.土壤酶及其研究法[M].北京:农业出版社,1986.
[3]García-Ruiz R, Ochoa V, Hinojosa M B, et al. Suitability of enzyme activities for the monitoring of soil quality improvement in organic agricultural systems[J]. Soil Biology & Biochemistry, 2008,40(9):2137-2145.
[4]蒲小鹏,李春荣,白小明,等.不同生长年限的草坪土壤酶活性研究[J].甘肃农业大学学报,2008,43(4):121-123.
[5]Xing Shihe, Chenrong Chen, Zhou Biqing, et al. Soil soluble organic nitrogen and active microbial characteristics under adjacent coniferous and broadleaf plantation forests[J]. Journal of Soils & Sediments, 2010,10(4):748-757.
[6]Ushio M, Kitayama K, Balser T C. Tree species effects on soil enzyme activities through effects on soil physicochemical and microbial properties in a tropical montane forest on Mt. Kinabalu, Borneo[J]. Pedobiologia, 2010,53(4):227-233.
[7]Magnuson T S. Comparison of extracellular peroxidaseand esterase-deficient mutants ofStreptomycesviridosporusT7 A[J]. Applied & Environmental Microbiology, 1992,58(3):1070-1072.
[8]程开明.结构方程模型的特点及应用[J].统计与决策,2006(10):22-25.
[9]王群,夏江宝,张金池,等.黄河三角洲退化刺槐林地不同改造模式下土壤酶活性及养分特征[J].水土保持学报,2012,26(4):133-137.
[10]王酉石,储诚进.结构方程模型及其在生态学中的应用[J].植物生态学报,2011,35(3):337-344.
[11]杨式雄,戴教藩,陈宗献,等.武夷山不同林型土壤酶活性与林木生长关系的研究[J].福建林业科技,1994,21(4):1-12.
[12]许景伟,王卫东,李成.不同类型黑松混交林土壤微生物,酶及其与土壤养分关系的研究[J].北京林业大学学报,2000,22(1):51-55.
[13]樊军,郝明德.黄土高原旱地轮作与施肥长期定位试验研究(Ⅱ):土壤酶活性与土壤肥力[J].植物营养与肥料学报,2003,9(2):146-150.
[14]黄巧云,李学垣.粘粒矿物,有机质对酶活性的影响[J].土壤学进展,1995,23(4):12-18.
[15]周礼恺,张志明,陈恩凤.黑土的酶活性[J].土壤学报,18(2):158-166.
[16]李小刚,崔志军,王玲英,等.盐化和有机质对土壤结构稳定性及阿特伯格极限的影响[J].土壤学报,2002,39(4):550-559.
[17]邹军,喻理飞,李媛媛.退化喀斯特植被恢复过程中土壤酶活性特征研究[J].生态环境学报,2010,19(4):894-898.
[18]赵聪,李勇,杨红军.缙云山森林次生演替群落土壤微生物、酶活性和养分的研究[J].中国农学通报,2012,28(4):46-50.
[19]尹瑞玲.微生物与土壤团聚体[J].土壤学进展,1985,13(4):24-29.
[20]郑伟,霍光华,骆昱春,等.马尾松低效林不同改造模式土壤微生物及土壤酶活性的研究[J].江西农业大学学报,2010,32(4):743-751.
[21]Ranjard L, Poly F, Combrisson J, et al. Heterogeneous cell density and genetic structure of bacterial pools associated with various soil microenvironments as determined by enumeration and DNA fingerprinting approach(RISA)[J]. Microbial Ecology,2000,39(4):263-272.
[22]吴明隆.结构方程模型:AMOS的操作与应用[M].重庆:重庆大学出版社,2009.
[23]余娜,刘济明,张超,等.不同沙生植被土壤酶活性分异特征研究[J].水土保持研究,2010,17(1):77-81.
[24]侯杰泰,温忠麟,成子娟.结构方程模型及其应用[M].北京:教育科学出版社,2001.
[25]Karlen D L, Stott D E. A framework for evaluating physical and chemical indicators of soil quality[C]∥Symposium on Defining Soil Quality for a Sustainable Environment, American Society of Agronomy Annual Meeting. Minneapolis: Soil Science Society of America, 1992.
[26]刘洪鹄,赵玉明,王秀颖,等.土壤肥力评价方法探讨[J].长江科学院院报,2008,25(3):62-66.
[27]刘广深,徐冬梅,许中坚,等.用通径分析法研究土壤水解酶活性与土壤性质的关系[J].土壤学报,2003,40(5):756-762.
[28]唐海滨,廖超英,刘莉丽,等.蔬菜大棚土壤脲酶、过氧化氢酶活性与土壤养分的关系[J].干旱地区农业研究,2011,29(3):165-168.
[29]Schjonning P, Thomsen I. K, Moldrup P, et al. Linking Soil Microbial Activity to Water and Air-Phase Contents and Diffusivities[J]. Soil Science Society of America Journal, 2003,67(1):156-165.
[30]徐咪咪.异龄林林分生长的结构方程模型分析研究[D].北京:北京林业大学,2010.
收稿日期:2015-06-10修回日期:2015-07-27
通讯作者:赵洋毅(1980—)男(汉族),吉林省长春市人,博士,副教授,主要从事水土保持和生态恢复研究。E-mail:yyz301@foxmail.com。
文献标识码:A
文章编号:1000-288X(2016)03-0338-08
中图分类号:S154.2
A Study on Effect of Soil Physicochemical Properties on Enzyme Activity Using Structural Equation in Rocky Desertification Areas in Eastern Part of Yunnan Province
SHU Shumiao1,2, ZHAO Yangyi2, HU Huirong1, XIONG Haoqin1,2, DUAN Xu2,3
(1.CollegeofEnvironmentScienceandEngineering,SouthwestForestryUniversity,Kunming,Yunnan650224,China; 2.NationalStationforForestEcosysteminYuxiCity,YunnanProvince,Xingping,Yunnan653400,China; 3.CollegeofForestry,SouthwestForestryUniversity,Kunming,Yunnan650224,China)
Abstract:[Objective] To determine the effect of different vegetation types and nutrients on the enzyme activity in order to provide a theoretical basis for the restoration of rocky desertification areas in the eastern part of Yunnan Province. [Methods] Soil enzyme activity was used to evaluate the effect of restoration, and the soil quality evaluation method based on fuzzy mathematics was adopted to derive the soil enzyme index under different restoration approaches to compare the effect of each approach. Based on correlation analysis of soil physical and chemical activity and enzyme activity, structural equation model was adopted to quantify soil physical and chemical environment and enzyme activity environment, and then to analyze the influence of different soil physical and chemical environment on different enzyme environment. [Results] The order for enzyme activity was shrub and grass mode>mixed coniferous and broadleaf forest> broadleaf forest mode>coniferous forest mode>bare land; According to the principle of structural equation model, the sub-nutrient environment composed of total phosphorus and organic matter had a positive impact on enzyme environment which was composed of the activity of urease, catalase and polyphenol oxidase activity. [Conclusion] The shrub and grass mode and mixed coniferous and broadleaf forest both have a good effect on the recovery of rocky desertification. The influence coefficient of nutrient environment on enzyme activity was 0.87.
Keywords:Karst; rocky desertification; vegetation restoration model; soil nutrients; enzyme activity; structural equation; eastern part of Yunnan Province
资助项目:云南省教育厅研究项目“滇东曲靖石漠化地区不同植被恢复模式对土壤质量的影响研究”(2013Y114); 云南省高校优势特色重点学科(生态学)建设资助项目(05000511311); 国家林业局林业科技创新平台项目(2015-LYPT-DW-138); 云南省应用基础研究项目(2012FD028)
第一作者:舒树淼(1988—),男(汉族),四川省成都市人,硕士研究生,研究方向为植物群落生态学。E-mail:838291221@qq.com。
