蕨类植物修复重金属污染的应用研究进展
2016-07-23杨桂英
杨桂英
摘要:蕨类植物是一类较为低等的维管束植物,系统演化上介于苔藓植物与种子植物之间,是陆生生态系统中重要的成员。该类植物具有适应力强、耐贫瘠等独特的生态学特征,且因某些种类对砷、锑的超量吸收和积累而广受关注。此外,蕨类植物对镉、铅、铜、镍等重金属及稀土元素的吸收也有不俗表现。本文从蕨类植物对重金属污染治理的应用、修复机制及优势进行了阐述,并展望了未来蕨类植物生态修复研究的前景及可能发展的研究领域。
关键词:蕨类植物;重金属污染;砷超富集植物;植物修复
中图分类号: X171.4文献标志码: A文章编号:1002-1302(2016)05-0010-05
现存的蕨类植物绝大多数为草本植物,种类众多[2]、生态类型丰富[3]、繁衍和适应方式多样[4],是植物多样性的重要组成部分。一些种类对极端恶劣环境的适应生存能力强,如膜蕨科植物Hymenophyllum sanguinolentum耐旱能力极强,可数天忍耐-40 MPa的渗透胁迫不死[5];西欧广布种Trichomanes speciosum耐瘠、耐弱光能力极强,甚至可在火山灰上生长[6];也有种类在贫瘠土壤[7]、金属废矿堆[8]及遭受严重污染的路域[9]等地生长。许多蕨类植物因对其生存环境的特殊适应,还可作为环境变化的指示者[10-13]。近些年发现的凤尾蕨属(Pteris)植物具有富集重金属元素的作用,尤其是对砷、锑等元素的积累效应明显,在重金属污染区域的生态修复中有着巨大的潜力。
1蕨类植物在重金属污染生态修复中的应用
植物修复技术自提出以来在重金属污染治理中备受关注,近年来有关重金属超富集植物已报道了450多种,分属于45个科[14-15],绝大部分都是关于镍的超富集植物(318种)[16],蕨类超富集植物有几十种,资料显示多为砷、锑富集植物,且多隶属于凤尾蕨科(Pteridaceae)。
1.1可富集砷的蕨类植物资源及表现
砷是一种致癌的准金属元素[17],其理化性质和环境行为与重金属多有相似之处,故在讨论重金属时往往包括砷[18]。正常土壤中普通植物的含砷量一般不超过3 mg/kg[19];而一些蕨类植物可超量吸收砷且能正常生长。
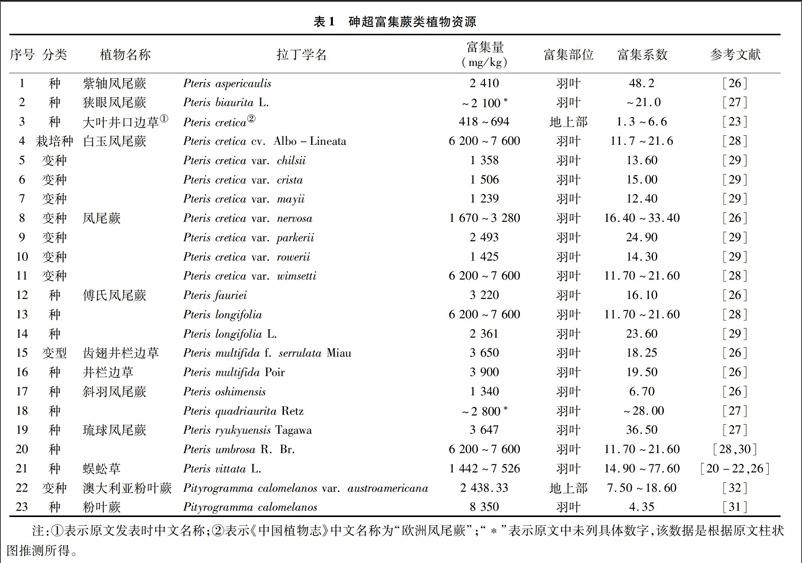
蜈蚣草是第一种被国内外学者同时期发现的砷超富集植物[20-22],其地上部累积的砷最高可达7 526 mg/kg,占地上部生物量(干质量)的2.3%,含量甚至高于植物中的磷含量。另一种砷超富集植物大叶井口边草地上部的平均含砷量为418 mg/kg(干质量),最大含砷量可达694 mg/kg;地下部(根)的平均含砷量为293 mg/kg,最大含砷量552 mg/kg,生物富集系数为1.3~4.8,转运系数(地上部砷含量与根砷含量之比)为1.0~2.6[23];其吸收的砷主要富集在羽叶的叶肉组织中[24-25]。另外,多种其他砷超富集植物被发现[26-30],多数集中在凤尾蕨科,也有一些从裸子蕨科(Hemionitedaceae)中筛选得到,如Visoottiviseth等发现泰国本土种Pityrogramma calomelanos羽叶中可富集砷8 350 mg/kg(干质量)[31];徐卫红等测定了变种澳大利亚粉叶蕨(P. calomelanos var. austroamericana)地上部砷含量达到2 438.33 mg/kg(干质量)[32]。迄今为止,国内外已发现砷超富集植物20多种(包括变种),具体如表1所示。
1.2可富集锑的蕨类植物资源及表现
锑(Sb)是地壳中的一种痕量元素[33],在植物体内,5~10 mg/kg 锑含量就能导致植物毒性的产生[19],到目前为止,仅有几种潜在的锑(超)富集植物被报道[34]。由于锑与砷具有相似的化学性质,而砷的超富集植物中许多都属于蕨类植物,故研究人员在蕨类植物中筛选锑富集植物。Tisarum等以采自美国、中国和巴西的蜈蚣草为材料,测定了Sb的富集浓度为4 192~12 000 mg/kg,且吸收的主要是三价锑[35];另一种砷超富集植物大叶井口边草对高浓度锑表现极强的耐性[36],其栽培变种——白玉凤尾蕨富集锑的最大浓度可达6 405 mg/kg[37];但富集的锑主要累积在根部,尤其在蜈蚣草中,吸收的总锑中99%以上累积在根部[35],转运系数极低。有关锑的转运方式,目前有确切证据的研究并不多,多为推测。Feng等认为锑的转运存在多种途径[38]。三价锑可以通过3价砷的转运通道,而五价锑的转运通道目前没有发现,可能是利用磷酸盐转运系统[35]。在被子植物中也有锑富集植物,早年的研究发现香叶蓍(Achillea ageratum)、长叶车前草(Plantago lanceolata)、狗筋麦瓶草(Silene vulgaris)对锑的富集浓度分别为1 367 mg/kg(基生叶)、1 150 mg/kg(根部)、1 164 mg/kg(茎部)[39];近年Affholder 等也发现,法国南部的迷迭香(Rosmarinus officinalis)在多种金属复合污染条件下栽1.3可富集其他重金属及稀土元素的蕨类植物资源及表现
现有资料显示蕨类植物对除砷、锑之外的其他重金属也具有较好的耐性和富集能力。Koller等在筛选超富集和耐性植物研究中发现,蜈蚣草和P. umbrosa除大量吸收砷外也可吸收铅、锌等重金属[41];Roccotiello等也发现蜈蚣草和Polypodium cambricum可同时吸收锌[42]。李影等研究节节草(Equisetum ramosissimum)和蜈蚣草对铜的吸收和迁移,结果表明节节草和蜈蚣草对铜具有较高的耐性和积累作用,其总积累量可达1 439. 47、398. 62 mg/kg,根系富集系数均明显大于1,可作为先锋植物来修复铜污染土壤[43-44]。
蕨类植物还能耐受多种重金属的复合污染,表现共富集特征。如水生蕨类植物槐叶苹(Salvinia natans)在富铬废水中可同时富集铬、镍、铁和镉[45],但其对镉较为敏感,半效应浓度仅为2.41mg/L[46],虽对水体镉有一定的净化作用,但极易受害;蜈蚣草在镉、铅、锰、铜和锌复合污染条件下,富集镉量在根部最大,其富集系数可达2.3[47];欧洲蕨(Pteridium aquilinum)在铬、镍复合污染条件下,羽叶中铬、镍吸收量均高于单一处理,即表现出协同现象[12],这可能与其进化程度低有关。
另外,生长在稀土矿区的蕨类植物铁芒萁(Dicranopteris linearis),其叶片积累的稀土含量高达3 263.8 mg/kg[48];芒萁(Dicranopteris dichotoma)对轻稀土元素镧、铈、钕表现出极强的积累富集能力,较生长于同一金矿环境的马尾松高出100~1 000倍[49],这一特性可用于稀土污染区域的修复。
2蕨类植物对重金属的富集/耐性机制
某些蕨类植物在重金属、类金属、稀土元素等污染环境下表现出较强的超积累、超富集、超耐性现象,其机理涉及吸收、转运、代谢、解毒等多个方面,随着蜈蚣草、粉叶蕨、白玉凤尾蕨、大叶井边兰等砷、锑超富集蕨类植物的发现,有关超富集植物的吸收、耐性和富集机制的研究越来越多。
2.1排出机制
一些蕨类植物可在分泌物中排出重金属或者积累在老叶中,通过落叶来排出多余的重金属。Tu等研究发现,蜈蚣草在砷含量>0.5 mg/kg土壤中生长时,较多的砷会在老叶中累积,并因其脱落而排出体外[50]。
2.2结合钝化作用与区隔化作用
细胞壁或细胞膜或液泡中存在与重金属等毒物结合的“结合座”,如细胞壁果胶质中的多聚糖醛酸和纤维素分子的羧酸、醛基等基团,都能与重金属结合,从而降低重金属向细胞质的运输而解毒;另外进入细胞质的重金属,还可以与小分子的有机酸(如柠檬酸、苹果酸)、氨基酸(如组氨酸),或者金属硫蛋白、植物螯合蛋白等结合。Nishizono等分析了禾秆蹄盖蕨(Athyrium yokoscense)的根细胞壁在重金属解毒中的作用,结果表明进入植物体的铜、锌、镉总量的70%~90%位于细胞壁,其中大部分以离子形式存在或结合到细胞壁的结构物质如纤维素、木质素上[51];Webb等通过X射线吸收光谱技术研究砷在蜈蚣草体内的分布,结果表明植物螯合蛋白可能对砷的大量蓄积起到积极作用[52];另外陈同斌等发现了另一种现象,即羽叶中有78%的砷分布在羽片胞液[24],这种区隔化作用(compartmentalization)是蜈蚣草解毒的重要原因[25]。
2.3抗氧化系统的作用
金属以及类金属进入植物体内会产生一些活性氧化物质(ROS),ROS的过量产生会对植物产生毒害,植物为了消除ROS会合成一些酶类以及非酶类抗氧化剂,这些抗氧化剂会清除过量的ROS,从而使正常的植物电子传递过程能够顺利进行。冯人伟研究蜈蚣草在硒胁迫下,叶片中还原型谷胱甘肽(GSH)的含量以及谷胱甘肽还原酶(GR)的活性显著增大,其耐性机理为GSH、GR起到调控超氧负自由基( O-2· )的作用,而POD、APX和CAT 3种酶仅在低浓度硒处理条件下起到清除H2O2的作用[53];Zhang等通过比较井栏边草和蜈蚣草酶的抗氧化系统,发现随着砷、铅浓度的升高,SOD和POD活性升高,GSH含量增加,减轻了金属毒害[54]。
2.4微生物的共同作用
根际微生物能促进植物对土壤中营养元素与重金属的吸收,或者通过分泌生长调节剂和保护植物的抗生素、抑菌剂或螯合素等方式以增强植物对污染环境的适应能力。在蕨类植物中,通过微生物强化污染物质吸收主要是针对砷的研究,试验所用微生物有细菌、真菌和放线菌。根际微生物绿脓假单胞菌(Pseudomonas aeruginosa)可提高欧洲凤尾蕨对砷的吸收,其机理是该菌在缺铁环境下可分泌铁载体(siderophores)于土壤中,与砷(约占总砷量的68%)结合形成铁砷复合物的形式,易被根系吸收进而转运到枝叶积累[55];另有研究发现,外源添加细菌Ts37,蜈蚣草地上部砷含量最高达837 mg/kg,施加放线菌shf2,地下部砷含量最高为427 mg/kg,分别比对照高206%、88%[56];Liu等也发现蜈蚣草接种菌根真菌Glomus mosseae后,在砷含量300 mg/kg土壤中种植,其砷累积量提高了43%[57]。
蕨类植物在长期进化过程中不断适应外界环境(包括污染的环境),在这个过程中可能发展出多种适应策略,当受污染环境胁迫时,通常是几种机制同时发挥作用。
3蕨类植物在生态修复中的优势所在
3.1起源古老,适应力强
蕨类植物是一种古老的植物,经过长期的自然选择和进化,具有广泛的适应性和顽强的生命力[6]。蜈蚣草具有极强的耐砷特性,能在砷含量为23 400 mg/kg 的矿渣上正常生长[21]。在森林植被尤其是在热带、亚热带森林植被恢复过程中,蕨类植物常成为草本层的优势种[58-59]。蕨类植物在次生迹地恢复过程中可以扮演重要的角色。
3.2具有独特的类金属砷耐性相关基因
蕨类植物具有耐贫瘠、生长迅速、繁殖能力强、抗逆性强等特点,对胁迫环境具有较强的耐性,可以弥补现有修复植物的某些缺点和不足。近年有关砷超富集蕨类植物的砷相关基因研究很多[60-62],试图从分子机理上解释超量吸收砷的原因。Indriolo等研究发现蜈蚣草具有编码亚砷酸转运蛋白的基因ACR3,在孢子体根系和配子体中的表达受砷的上调作用,其定位于液泡膜上,而该基因在被子植物中缺失[63]。可见,蕨类植物在砷污染的修复上拥有被子植物所不具有的“先天”优势。
3.3蕨类植物蓄积的无机污染物种类繁多、选择性差
蕨类植物能够蓄积多种无机污染物,选择性差[64]。一些水生蕨类植物耐污能力强,且可以富集多种重金属。如红萍(Azolla pinnata var. imbricata)(又名覆瓦状满江红)在大多数污水中均能生长,其抗酸碱能力、耐盐和耐肥能力均较强,pH值的耐受极限为2.8、12.6,具有生活能力的生理抗酸碱范围是3.5~11.7,对水环境中氮、磷耐受极限分别为175、800 mg/L[65];蕨状满江红(A. filiculoides)(又名细绿萍)对污水河中的镉、砷和锌[66]以及铅、汞[67]都有明显的吸收积累作用,并能吸收大量的氮和磷[68-69],铵态氮、硝态氮和磷的去除率最大分别可达94.04%、95.46%、99.0%以上[70];蜈蚣草在富集镉的同时,可大量吸收铅、锰、铜、锌[47];此外,满江红(A. imbircata)还能吸收放射性铀,在水培条件下对铀的去除率达到了92%~97%[71];欧洲蕨可同时积累铬和镍[12]。蕨类植物这种吸收选择性差的特性可能与其进化程度低有关,但是从植物修复的角度看,这种广谱性具有极大的应用价值。
4展望
在陆地生态系统的形成与演化过程中,蕨类植物是首先征服陆地生态环境的先锋类群,在现代植物中,蕨类植物从古生代的繁盛历经几亿年的环境变迁之后,种类数量和个体数量均锐减,与种子植物相比是个较小的类群,但作为植物界中的重要成员之一,仍然承担着陆地生态系统多样性和生态平衡的重要角色。蕨类植物具有独特的生物学和生态学特征,在受损生态系统的修复中具有较大的应用潜力。
近年来,利用蕨类植物的超富集砷的特性进行植物修复以及砷超富集机制的相关研究取得了大量成果,未来可能在以下方面具有较大的发展空间:(1)复合污染条件下,蕨类植物对砷、锑等重金属的吸收、富集过程中与其他元素的相互作用及影响规律。(2)基于现有基因的克隆和表征,通过分子设计和遗传修饰培育新的适应广、易栽培、生物量大的工程植物,用于修复重金属污染的环境。(3)蕨类植物对有机农药等新型污染物的生物降解潜力挖掘。Nesterenko-Malkovskaya等[72]曾报道凤眼莲(Eichhornia crassipes)在没有根际细菌辅助的情况下能去除废水中的萘,说明植物在降解有机污染物过程中也具有相当大的贡献,蕨类植物是否具有这种能力目前尚不清楚,有待研究。(4)具修复潜力的蕨类植物快繁新技术研究。由于蕨类植物不是通过种子繁殖,因此如何高效繁育种苗可能成为修复应用的一个瓶颈问题。虽然目前也有一些繁育方法[73-75],但成活率较低,难以满足大田作业。(5)富集植物终期风险控制和回收处理新技术的研发。蕨类植物富集重金属后如果不加控制,容易形成生态风险[76],Mathews等发现在富集砷蕨类植物种植区,植食动物等消费者体内砷含量远超对照,甚至达到杀伤的效应,若无有效的风险控制机制,很可能导致生态失衡[77-78]。如果按照一般填埋、焚烧等老方法处理,又易造成二次污染,因此高效安全的资源化利用新技术研发有着广阔的前景。
参考文献:
[1]李星学,周志炎,郭双兴. 植物界的发展和演化[M]. 北京:科学出版社,1981:50-51.
[2]陆树刚. 中国蕨类植物区系[M]//吴征镒,陈心启.中国植物志(第一卷).北京:科学出版社,2004:78-94.
[3]陆树刚,陈风. 论蕨类植物生态类型的划分问题[J]. 云南大学学报:自然科学版,2013,35(3):407-415.
[4]吴兆洪,秦仁昌. 中国蕨类科属志[M]. 北京:科学出版社,1991:4.
[5]Proctor M C F. Light and desiccation responses of some Hymenophyllaceae (filmy ferns) from Trinidad,Venezuela and New Zealand:poikilohydry in a light-limited but low evaporation ecological niche[J]. Annals of Botany,2012,109(5):1019-1026.
[6]Johnson G N,Rumsey F J,Headley A D,et al. Adaptations to extreme low light in the fern Trichomanes speciosum[J]. New Phytologist,2000,148(3):423-431.
[7]Page C N. Ecological strategies in fern evolution:a neopteridological overview[J]. Review of Palaeobotany and Palynology,2002,119(1):1-33.
[8]李影,陈明林. 节节草生长对铜尾矿砂重金属形态转化和土壤酶活性的影响[J]. 生态学报,2010,30(21):5949-5957.
[9]Oliver M J,Tuba Z,Mishler B D. The evolution of vegetative desiccation tolerance in land plants[J]. Plant Ecology,2000,151(1):85-100.
[10]Gupta M,Devi S. Uptake and toxicity of cadmium in aquatic ferns[J]. J Environ Biology,1995,16(2):131-136.
[11]Chang J S,Yoon I H,Kim K W. Heavy metal and arsenic accumulating fern species as potential ecological indicators in As contaminated abandoned mines[J]. Ecol Indic,2009,9(6):1275-1279.
[12]Kamila K,Aleksandra S C,Krzysztof K,et al. Chromium and nickel in Pteridium aquilinum from environments with various levels of these metals[J]. Environ Sci Pollut Res,2015,22(1):527-534.
[13]严岳鸿,张宪春,马克平. 中国蕨类植物多样性与地理分布[M]. 北京:科学出版社,2013:17-26.
[14]Prasad M N V,Freitas H,Fraenzle S,et al. Knowledge explosion in phytotechnologies for environmental solutions[J]. Environ Pollut,2010,158(1):18-23.
[15]Sebastian A,Prasad M N V. Cadmium minimization in rice:a review[J]. Agron Sustain,2014,34:155-173.
[16]Kramer U,Cotter-Howells J D,Charnock J M,et al. Free histidine as a metal chelator in plants that accumulate nickel[J]. Nature,1996,379(6566):634-738.
[17]Nriagu J O. Arsenic in the environment. Part Ⅱ:Human health and ecosystem effects[M]. New York:John Wiley&Sons,1994:1-5.
[18]陈怀满. 土壤中化学物质的行为与环境质量[M]. 北京:科学出版社,2002:79.
[19]Kabata-Pendias A,Pendias H. Trace elements in soils and plants[M]. 3nd ed. Boca Raton,USA:CRC Press,2001:3-20.
[20]Ma L Q,Komar K M,Tu C,et al. A fern that hyperaccumulates arsenic:a hardy,versatile,fast-growing plant helps to remove arsenic from contaminated soils[J]. Nature,2001,409(6820):579.
[21]陈同斌,韦朝阳,黄泽春,等. 砷超富集植物蜈蚣草及其对砷的富集特征[J]. 科学通报,2002,47 (3):207-210.
[22]Chen T B,Wei C Y,Huang Z C,et al. Arsenic hyperaccumulator Pteris vittata L. and its arsenic accumulation[J]. Chinese Science Bulletin,2002,47(11):902-905.
[23]韦朝阳,陈同斌,黄泽春,等. 大叶井口边草——一种新发现的富集砷的植物[J]. 生态学报,2002,22(5):777-778.
[24]陈同斌,黄泽春,黄宇营,等. 砷超富集植物中元素的微区分布及其与砷富集的关系[J]. 科学通报,2003,45(11):1163-1168.
[25]陈同斌,阎秀兰,廖晓勇,等. 蜈蚣草中砷的亚细胞分布与区隔化作用[J]. 科学通报,2005,50(24):2739-2744.
[26]Wang H B,Wong M H,Lan C Y,et al. Uptake and accumulation of arsenic by 11 Pteris taxa from southern China[J]. Environmental Pollution,2007,145(1):225-233.
[27]Srivastava M,Ma L Q,Santos J A G. Three new arsenic hyperaccumulating ferns[J]. Science of the Total Environment,2006,364(1):24-31.
[28]Zhao F J,Dunham S J,McGrath S P. Arsenic hyperaccumulation by different fern species[J]. New Phytologist,2002,156 (1):27-31.
[29]Meharg A A. Variation in arsenic accumulation-hyperaccumulation in ferns and their allies[J]. New Phytologist,2003,157(1):25-31.
[30]Koller C E,Patrick J W,Rose R J,et al. Pteris umbrosa R. Br. as an arsenic hyperaccumulator:accumulation,partitioning and comparison with the established As hyperaccumulator Pteris vittata[J]. Chemosphere,2007,66 (7):1256-1263.
[31]Visoottiviseth P,Francesconi K,Sridokchan W .The potential of Thai indigenous plant species for the phytoremediation of arsenic contaminated land[J]. Environmental Pollution,2002,118(3):453-461.
[32]徐卫红,Singh B,Kachenko A G. 砷超积累植物粉叶蕨及其对砷的吸收富集研究[J]. 水土保持学报,2009,23(2):173-177.
[33]He M,Wang X,Wu F,et al. Antimony pollution in China[J]. Sci Total Environ,2012,421/422:41-50.
[34]冯人伟,韦朝阳,涂书新. 植物对锑的吸收和代谢及其毒性的研究进展[J]. 植物学报,2012,47(3):302-308.
[35]Tisarum R,Lessl J T,Dong X,et al. Antimony uptake,efflux and speciation in arsenic hyperaccumulator Pteris vittata[J]. Environmental Pollution,2014,186:110-114.
[36]Feng R W,Wei C Y,Tu S X,et al. Simultaneous hyperaccumulation of arsenic and antimony in Cretan brake fern:evidence of plant uptake and subcellular distributions[J]. Microchem J,2011,97(1):38-43.
[37]Feng R,Wang X,Wei C,et al. The accumulation and subcellular distribution of arsenic and antimony in four fern plants[J]. International Journal of Phytoremediation,2015,17(4):348-354.
[38]Feng R W,Wei C Y,Tu S X,et al. The uptake and detoxification of antimony by plants:a review[J]. Environ Exp Bot,2013,96:28-34.
[39]Baroni F,Boscagli A,Protano G,et al. Antimony accumulation in Achillea ageratum,Plantago lanceolata and Silene vulgaris growing in an old Sb-mining area[J]. Environmental Pollution,2000,109(2):347-352.
[40]Affholder M C,Prudent P,Masotti V,et al. Transfer of metals and metalloids from soil to shoots in wild rosemary (Rosmarinus officinalis L.) growing on a former lead smelter site:human exposure risk[J]. Sci Total Environ,2013,454:219-229.
[41]Koller C E,Patrick J W,Rose R J,et al. Arsenic and heavy metal accumulation by Pteris vittata L. and P. umbrosa R. Br[J]. Bulletin of Environmental Contamination and Toxicology,2008,80(2):128-133.
[42]Roccotiello E,Manfredi A,Drava G,et al. Zinc tolerance and accumulation in the ferns Polypodium cambricum L. and Pteris vittata L.[J]. Ecotoxicology and Environmental Safety,2010,73(6):1264-1271.
[43]李影,褚磊. 节节草对Cu 的吸收和积累特性[J]. 生态学报,2008,28(4):1565-1572.
[44]李影,王友保. 4种蕨类草本植物对Cu的吸收和耐性研究[J]. 草业学报,2010,19(3):191-197.
[45]Dhir B,Sharmila P,Pardha Saradhi P,et al. Physiological and antioxidant responses of Salvinia natans exposed to chromium-rich wastewater[J]. Ecotoxicology and Environmental Safety,2009,72(6):1790-1797.
[46]徐勤松,计汪栋,杨海燕,等. 镉在槐叶苹叶片中的蓄积及其生态毒理学分析[J]. 生态学报,2009,29(6):3019-3027.
[47]何志坚,薛鸿. 蜈蚣草野外复合污染条件下镉等重金属富集量的测定[J]. 绵阳师范学院学报,2011,30(11):130-134.
[48]李凡庆,毛振伟,等. 铁芒萁植物体中稀土元素含量分布的研究[J]. 稀土,1992,13(5):16-19.
[49]苗莉,徐瑞松,马跃良,等. 河台金矿矿山土壤-植物稀土元素含量分布和迁移积聚特征[J]. 生态环境,2008,17(1):350-356.
[50]Tu C,Ma L Q. Effects of arsenic concentrations and forms on arsenic uptake by the hyperaccumulator ladder brake[J]. Journal of Environmental Quality,2002,31(2):641-647.
[51]Nishizono H,Minemura H,Suzuki S. An inducible copper-thiolate in the fern,Athyrium yokoscense:involvement in copper tolerance of the fern[J]. Plant Cell Physiol,1988,29(8):1345-1351.
[52]Webb S M,Gaillard J F,Ma L Q,et al . XAS speciation of arsenic in a hyperaccumulating fern[J]. Environ Sci Techn,2003,37(4):754-760.
[53]冯人伟. 植物对砷、硒、锑的富集及抗性机理研究[D]. 武汉:华中农业大学,2009.
[54]Zhang K M,Deng T,Fang Y M,et al. Influence of co-contamination of As and Pb on the frond physiology and ultrastructure of Pteris vittata L.[J]. Fresenius Environmental Bulletin,2011,21(8):2215-2223.
[55]Jeong S,Moon H S,Nam K. Enhanced uptake and translocation of arsenic in Cretan brake fern (Pteris cretica L.) through siderophorearsenic complex formation with an aid of rhizospheric bacterial activity[J]. Journal of Hazardous Materials,2014,280:536-543.
[56]赵根成,廖晓勇,阎秀兰,等. 微生物强化蜈蚣草累积土壤砷能力的研究[J]. 环境科学,2010,31(2):431-436.
[57]Liu Y,Zhu Y G,Chen B D,et al. Influence of the arbuscular mycorrhizal fungus Glomus mosseae on uptake of arsenate by the As hyperaccumulator fem Pteris vittata L.[J]. Mycorrhiza,2005,15(3):187-192.
[58]胡理乐,刘琪,闫伯前,等. 生态恢复后的千烟洲植物群落种类组成及结构特征[J]. 林业科学研究,2006,19(6):807-812.
[59]高雷,刘宏茂.西双版纳热带雨林下砂仁拔除后的生态恢复研究[J]. 植物生态学报,2003,27 (3):366-372.
[60]Sundaram S,Wu S,Ma L Q,et al. Expression of a Pteris vittata glutaredoxin PvGRX5 in transgenic Arabidopsis thaliana increases plant arsenic tolerance and decreases arsenic accumulation in the leaves[J]. Plant,Cell and Environment,2009,32(7):851-858.
[61]何振艳,麻密,徐文忠,等. 一种植物砷抗性相关的蛋白及编码基因及其应用:中国,ZL201010116397.2[P]. 2013-05-01.
[62]Kumar S,Dubey R S,Tripathi R D,et al. Omics and biotechnology of arsenic stress and detoxification in plants:current updates and prospective[J]. Environment International,2015,74(6):221-230.
[63]Indriolo E,Na G N,Ellis D,et al. A Vacuolar arsenite transporter necessary for arsenic tolerance in the arsenic hyperaccumulating fern Pteris vittata is missing in flowering plants[J]. The Plant Cell,2010,22(6):2045-2057.
[64]郑洁敏,唐世荣,陈子元,等. 蕨类植物对无机污染物的吸收[J]. 核农学报,2005,19(2):155-159.
[65]郎业广,赵慧琴,刘忠阳,等. 红萍净化污水的研究[C]. 福州:国际红萍利用学术讨论会,1985.
[66]齐恩山,刘期松,杨桂芬,等. 凤眼莲等水生植物对灌溉重金属污水净化作用的初步研究[J]. 生态学杂志,1984,9(1):14-18.
[67]任安芝,唐廷贵. 细绿萍对铅、汞污水的净化作用及其生物学效应[J]. 南开大学学报:自然科学版,1996,29(1):74-79.
[68]Forni C,Chen J,Tancioni L,et al. Evaluation of the fern azolla for growth,nitrogen and phosphorus removal from wastewater[J]. Water Res,2001,35(6):1592-1598.
[69]易厚燕,吴爱平,王华. 满江红在不同氮浓度中对氮、磷的吸收效果[J]. 山地农业生物学报,2013,32(2):138-142.
[70]熊集兵,常会庆,何振立,等. 低温条件下满江红对地表水氮磷的去除效应研究[J]. 水土保持学报,2007,21(6):96-99.
[71]胡南,丁德馨,李广悦,等. 五种水生植物对水中铀的去除作用[J]. 环境科学学报,2012,32(7):1637-1645.
[72]Nesterenko-Malkovskaya A,Kirzhner F,Zimmels Y,et al. Eichhornia crassipes capability to remove naphthalene from wastewater in the absence of bacteria[J]. Chemosphere,2012,87 (10):1186-1191.
[73]李日新. 几种蕨类植物无性繁殖技术研究[J]. 国土与自然资源研究,2011(2):55-56.
[74]黄碧光,陈庆山,林文雄.水蕨的农田生境调查和人工繁育探索[J]. 中国生态农业学报,2012,20(6):810-812.
[75]李和平,刘兴剑,陈艳,等. 土生蕨类植物引种栽培技术[J]. 江苏农业科学,2014,42(11):213-214.
[76]沈羽,张开梅,方炎明. 蕨类植物修复土壤与净化水体的研究进展[J]. 江苏农业科学,2014,42(1):11-14.
[77]Mathews S,Ma L Q,Bala R,et al. Arsenic reduced scale-insect infestation on arsenic hyperaccumulator Pteris vittata L.[J]. Environmental and Experimental Botany,2009,65(2/3):282-286.
[78]Jeong S,Moon H S,Nam K. Increased ecological risk due to the hyperaccumulation of As in Pteris cretica during the phytoremediation of an As-contaminated site[J]. Chemosphere,2015,122:1-7.韩晗,韦晓婷,魏昳,等. 沙门氏菌对食品的污染及其导致的食源性疾病[J]. 江苏农业科学,2016,44(5):15-20.doi:10.15889/j.issn.1002-1302.2016.05.004
