大连市皮口海域口虾蛄群体繁殖生物学特征初步研究
2016-07-22薛梅闫红伟刘海映刘奇邢坤陈雷吕海波卢羽洁张涛迟庆宏
薛梅,闫红伟,刘海映,刘奇,邢坤,陈雷,吕海波,卢羽洁,张涛,迟庆宏
(1.大连海洋大学辽宁省海洋牧场工程技术研究中心,辽宁 大连116023;2.大连海洋大学辽宁省海洋生物资源恢复与生境修复重点实验室,辽宁大连116023;3.大连海洋大学水产与生命学院,辽宁大连116023;4.大连天正实业有限公司,辽宁 大连116000;5.大连市长海县水产技术推广站,辽宁长海116500)
大连市皮口海域口虾蛄群体繁殖生物学特征初步研究
薛梅1、2,闫红伟3,刘海映1、2,刘奇1、2,邢坤1、2,陈雷1、2,吕海波1、2,卢羽洁1、2,张涛4,迟庆宏5
(1.大连海洋大学辽宁省海洋牧场工程技术研究中心,辽宁 大连116023;2.大连海洋大学辽宁省海洋生物资源恢复与生境修复重点实验室,辽宁大连116023;3.大连海洋大学水产与生命学院,辽宁大连116023;4.大连天正实业有限公司,辽宁 大连116000;5.大连市长海县水产技术推广站,辽宁长海116500)
摘要:为了解大连市皮口海域口虾蛄Oratosquilla oratoria群体的繁殖生物学特征,于2014年5月—2015年4月,每月中旬对皮口海域口虾蛄群体进行随机取样,采用组织学和电镜技术对其性腺发育规律和生殖细胞发生过程进行了初步研究。结果表明:除7月份以外,其他月份的雌、雄口虾蛄比例与期望值1∶1之间均无显著性差异 (χ2=0.500,P>0.05);口虾蛄雌、雄性比为0.79~1.34,全年雌、雄性比平均为1.04± 0.05,雌、雄性腺周年发育变化趋势类似,雄性发育略快,性成熟高峰期均有2个,雌性在5月和11月达性成熟高峰期,而雄性在4月和11月达性成熟高峰期;口虾蛄卵子发生过程分为卵原细胞、初级卵母细胞、次级卵母细胞、卵黄形成前期细胞、卵黄形成期细胞、早期成熟期卵细胞和成熟期卵细胞,卵巢发育分为未发育时期、卵母细胞期、生长前期、生长中期、生长后期、成熟前期、成熟期和恢复期;精子的发生过程分为精原细胞、精母细胞和精子,精巢发育分为精原细胞期、精母细胞期、早期精子期和精子期。本研究结果可为口虾蛄繁殖生物学的深入研究奠定理论基础,并为早日实现口虾蛄栖息地养护及人工增养殖提供参考。
关键词:口虾蛄;性腺;生殖细胞;组织学;超微结构
口虾蛄Oratosquilla oratoria隶属于节肢动物门Arthropoda、软甲纲 Malacostraca、口足目 Stomatopoda、虾蛄科 Squillidae、口虾蛄属 Oratosquilla[1],俗称虾爬子、皮皮虾等[2],在中国沿海均有分布,以黄海、渤海产量最多[3],一般生活在5~60 m深水域,营穴居生活。因其肉嫩味美、营养丰富,是一种重要的海产经济动物。近年来,由于捕捞过度及环境恶化等影响,口虾蛄资源严重衰退,需求量和市场价格不断攀升。为了保护现有资源并满足日益增长的市场需求,口虾蛄增养殖势在必行。规模化繁育人工苗种,是进行增殖放流或人工养殖的关键过程,而充分了解口虾蛄繁殖生物学规律是推动这一过程的首要前提。
性腺不仅是主要的繁殖器官,而且是性细胞发育的重要基础[4]。甲壳动物生殖细胞发生是个体发育的重要环节,研究经济甲壳动物生殖细胞发育规律可为进行人工增养殖研究奠定理论基础。有关虾蛄属繁殖生物学特征的研究国内外已有一些报道,并积累了一定的研究成果。王春琳等[5]观察了黑斑口虾蛄 Oratosquilla kempi雄性生殖系统;Yamazaki等[6]、Kodama等[7]、Ohtomi等[8]和徐善良等[9]对口虾蛄卵巢发育进行了初步研究;刘海映等[10]研究了口虾蛄生殖细胞发生;Kodama等[7]探讨了口虾蛄精巢发育。截至目前,由于缺乏系统的观察数据,口虾蛄生殖细胞的发生与繁殖规律尚不连贯,发育分期也未有统一标准,影响了对口虾蛄繁殖生物学的进一步研究。
大连市皮口海域是黄海海域口虾蛄主要产区之
1 材料与方法
1.1材料
试验用口虾蛄捕自辽宁省大连市皮口海域(图1),采样时间为2014年5月—2015年4月,每月中旬采样,除1、2月份采集样本220尾外,其余各月每次采集样本300余尾。每次采样时均记录海区水温、盐度。

图1 采样地点Fig.1 Location of sampling site
1.2方法
1.2.1组织学样品的制备 每月采样后,首先根据有无交接肢将全部样品区分,记录口虾蛄雌、雄数量,估算性比。测量所有样品的体长 (BL)、体质量 (BW)和完整性腺质量。同时从中分别随机抽取雌、雄各60尾,解剖获得精巢、卵巢,并从性腺组织中段切取小块组织,经Bouin氏液固定24 h,按常规石蜡切片方法进行脱水、透明、包埋。使用YD 355AT(China)切片机进行组织切片,切片厚度为 5~7 μm,经Harris苏木精-伊红染色后[11],于Nikkon Eclipse 80i(Japan)显微镜下观察并拍照,用目微尺测量细胞及核仁大小。此外,对成熟的卵巢使用二氧六环 [(C2H4)2O2]代替二甲苯 (C8H10)进行透明[12],以防止组织块脆化和切片时发生破碎脱落现象。
1.2.2超显微样品的制备 分别从精巢、卵巢的中段切取小块 (1 mm×1 mm×3 mm)组织,迅速用0.1 mol/L磷酸缓冲溶液配制的戊二醛和1%锇酸双重固定[13],用乙醇梯度脱水、环氧树脂包埋、超薄切片,经醋酸铀和柠檬酸铅染色后,在JEM 2100透射电镜下 (Japan)观察并拍照。
1.2.3性腺指数的计算
性腺指数=卵巢质量/体质量×100%。
1.3数据处理
用Excel 2013软件对试验数据进行常规处理,采用SPSS 22.0软件进行性腺指数单因素方差分析(One-Way ANOVA),用Turkey法检验月间差异。
此外,性别比例 (雌∶雄)以1∶1为期望值,采用卡方检验 (χ2=0.50,df=2)进行差异性分析。口虾蛄雌、雄之间的体长、体质量差异性比较采用单样本T检验。所有统计值显著性水平设为0.05,统计结果均以平均值±标准误 (mean± S.E.)表示。
2 结果与分析
2.1环境因子的季节性变化
采捕口虾蛄的海区水温及盐度的周年变化如图2所示。周年水温变化范围为4~25℃,水温从2月开始上升,8月时达到全年最高值25℃,随后逐渐降低,年平均水温为(13.50±7.03)℃,月间变化显著 (P<0.05)。周年盐度变化范围为29~33.60,年平均盐度为31.09±1.33,月间变化不显著 (P>0.05)。

图2 采样海域水温和盐度的周年变化Fig.2 Monthly change in water temperature and salinity in the sampling sites
2.2口虾蛄性比的周年变化
本次调查在大连市皮口海域全年共采集3446尾口虾蛄样品,其中雌性个体1746尾,雄性个体1700尾,雌、雄比例为1.03∶1。周年雌、雄口虾蛄性别比例变化为0.79~1.34,周年平均性比为1.04±0.05(表1)。卡方检验表明:除7月外,全年口虾蛄雌、雄比例与期望值1∶1之间均无显著性差异 (P>0.05)。
2.3体长、体质量
从表1可见:雌、雄口虾蛄体长范围分别为8.60~18.50 cm、9.90~17.40 cm;雌性口虾蛄5 月 (水温17℃)平均体长达周年最大值 (13.87 cm),之后持续降低,8月 (水温25℃)平均体长最小 (11.88 cm);雄性口虾蛄4月 (水温13℃)平均体长达周年最大值 (14.42 cm),之后降低,8月 (水温25℃)平均体长最小 (12.37 cm);一周年内,除3月、5月外,其余月份雄性口虾蛄的平均体长均大于雌性。
口虾蛄平均体质量变化规律与体长类似,雄性口虾蛄均大于雌性口虾蛄;3月 (水温12℃)雌性口虾蛄平均体质量最大 (33.34 g),11月 (水温7℃)雄性口虾蛄平均体质量达到周年最大值(39.17 g),而8月 (水温25℃)雌、雄性口虾蛄平均体质量均最小,分别为20.42 g和24.34 g(表1)。
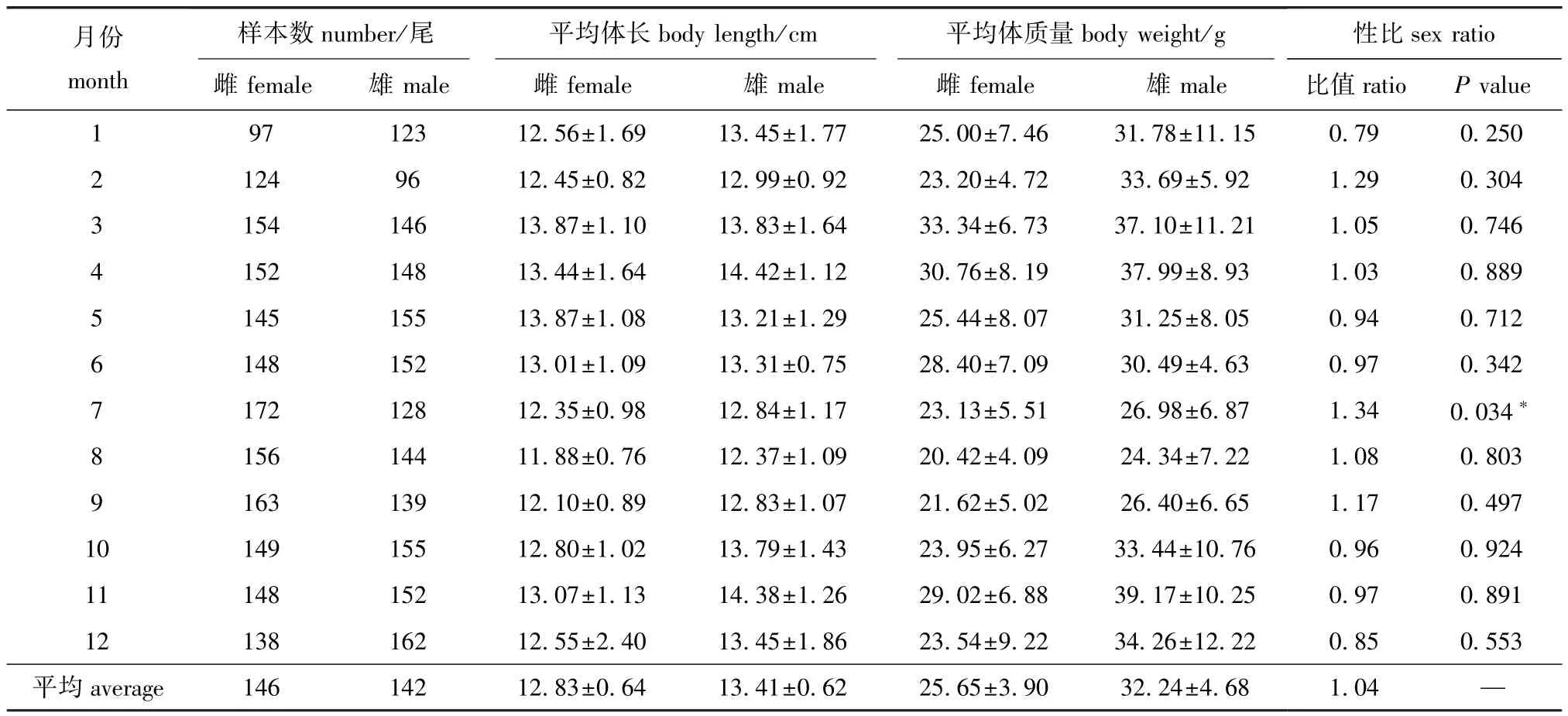
表1 大连市皮口海域口虾蛄体长、体质量和性比的周年变化 (平均值±标准误)Tab.1 Monthly change in body length,body weight and sex ratio of mantis shrimp Oratosquilla oratoria in the area of Pikou Bay,Dalian(mean±S.E.)
2.4繁殖规律
解剖样品发现,口虾蛄为雌雄异体。口虾蛄性腺位于围心窦和肝胰腺之间[14-15]。精巢质量测定显示,雄性口虾蛄性成熟高峰期全年出现2次(图3),分别发生在4月和11月。此外,采用每月雌性口虾蛄性腺指数表示当月性成熟度 (图4),结果发现:口虾蛄当年即可达性成熟,自2月(水温4℃)开始,性腺指数上升;5月 (水温17℃)达全年最高值,为9.61%,此时性腺发育成熟,肉眼可见黄色性腺占据整个背部区域,尾节处融合,末端呈黄色三角形;随后,性腺指数降低,至9月 (水温21℃)性腺指数仅为0.85%,性腺成熟度达全年最低,此时性腺退化成黑线状;之后,性腺指数再次升高,至11月 (水温7℃)性腺指数为4.28%。由此可见,大连市皮口海域雌性口虾蛄性成熟高峰期也有2个,分别在5月和11月。
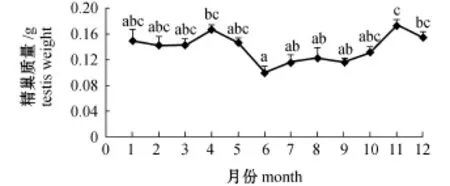
图3 大连市皮口海域口虾蛄精巢质量的周年变化Fig.3 Monthly change in testis weight of mantis shrimp Oratosquilla oratoria in the area of Pikou Bay,Dalian
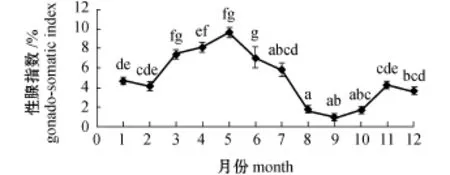
图4 大连市皮口海域雌性口虾蛄性腺指数的周年变化Fig.4 Monthly change in gonado-somatic index of female mantis shrimp Oratosquilla oratoria in the area of Pikou Bay,Dalian
2.5卵子发生和卵巢发育分期
口虾蛄有一对卵巢,呈左右对称,两个卵巢趋于愈合,仅由一层隔膜相隔,卵属于多黄卵,卵子发育由中间隔膜逐渐向两侧发育 (图5)。根据卵细胞的形态、卵黄积累和滤泡细胞的形状,卵子的发生过程分为7种配子体。
卵原细胞:细胞很小,大小仅为 (2.0±1.4)μm,来自卵巢边缘的生殖上皮细胞;细胞核位于中央。卵原细胞外围结缔组织中出现滤泡细胞(图6-A)。
初级卵母细胞:细胞增大,呈椭圆形,卵径为(2.0±8.5)μm;细胞核呈椭圆形,增大至(18.0±3.8)μm,细胞膜、核膜不明显,无核仁,细胞质少 (图6-B)。
次级卵母细胞:细胞明显增大,直径为(50.0±18.5)μm;细胞核增大至 (28.0±5.8)μm,核仁清晰,核膜明显,核中丝状的染色体出现,卵细胞周围有单层滤泡细胞围绕,其主要作用是保护卵细胞发育,并为卵细胞发育提供营养物质,此时的滤泡细胞呈卵圆形 (图6-C)。
卵黄形成前期细胞:细胞继续增大,直径为(100.0±56.9)μm;细胞核明显,为 (54.0± 10.5)μm,能看到核中有丝状的染色体分布,核仁清晰,细胞内出现网状结构,滤泡细胞变得扁长,围绕在卵细胞周围 (图6-D)。
卵黄形成期细胞:细胞大小为 (370.0±70.9)μm;细胞核大小为 (55.0±14.2)μm,此期核内丝状染色质消失,核仁明显,滤泡细胞变成长条形(图6-E)。
早期成熟期卵细胞:细胞呈不规则多边形,直径为 (400.0±82.6)μm;细胞核为 (30.0±14.2)μm,核膜消失,细胞核向细胞边缘移动,细胞质中网状结构消失,滤泡细胞拉伸 (图6-F)。
成熟期卵细胞:细胞体积达到最大,边缘呈多角结构,细胞大小为 (500.0±137.6)μm;细胞核逐渐解体、消失,卵细胞内充满卵黄,并形成较大的卵黄颗粒。组织学和超微观察均显示,此阶段的细胞还具有大小不一的油滴,滤泡细胞呈细条状,细胞核也呈类似结构 (图6-G、H、I)。
根据口虾蛄卵巢外形、颜色变化和卵细胞形态特征,将卵巢发育全过程划分为8个时期。
未发育时期:卵巢呈细小线状;左右卵巢内有大量的卵原细胞,同时存在少量的初级卵母细胞,此时的卵巢内含有营养细胞 (图7-A)。
初级卵母细胞期:卵巢扩大呈带状,存在大量初级卵母细胞及少量卵原细胞 (图7-B)。
生长前期:卵巢进一步发育,向两侧扩张,出现少许卵黄,此时卵巢呈浅黄色;卵巢中含有卵原细胞、初级卵母细胞、次级卵母细胞、滤泡细胞,且细胞排列松散 (图7-C)。
生长中期:卵巢体积明显增大,边缘凹陷明显;卵细胞排列紧密,卵细胞被单层滤泡细胞分隔包围,卵巢中含有大量次级卵母细胞 (图7-D)。
生长后期:卵巢迅速生长,两侧凹凸呈波浪状,卵黄不断累积,卵巢呈黄色;卵巢中全是卵黄,形成前期细胞,“S型”增殖区开始出现 (图7-E)。
成熟前期:卵巢饱满,卵黄迅速积累,在尾节融合,尾节中间出现黄色长条;卵巢壁变薄,细胞排列紧密,早期成熟卵细胞占据整个卵巢组织,卵巢中 “S型”区域仍然存在,其内细胞为初级卵母细胞 (图7-F)。
成熟期:卵巢极度膨大,尾节中间出现黄色三角形;此期细胞为成熟期细胞,排列紧密,肉眼可见卵细胞颗粒 (图7-G)。
恢复期:此期为排卵后,整个卵巢开始萎缩,卵细胞分布稀疏,细胞质松散,细胞核模糊;存在许多卵巢小管,部分卵巢小管包围多个萎缩的卵细胞,细胞与细胞之间出现明显间隙 (图7-H)。
2.6精子发生和精巢发育分期
口虾蛄也有一对精巢,左右对称,仅由一层膜相隔,相互连接,但不愈合。精巢由精巢管壁和精巢腔组成 (图8),未发现其他虾蟹类所具有的生精腺囊或生精细管[16]等结构。整个管状精巢内均可进行精子的发生,成年口虾蛄精巢展开长度为50~60 mm,成熟精巢质量只有成熟卵巢质量的5%~7%。精巢外部由一层薄薄的结缔组织包裹,可对精细胞起到保护和缓冲作用。
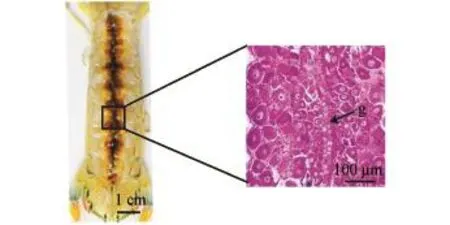
图5 口虾蛄卵巢Fig.5 The ovary of mantis shrimp Oratosquilla oratoria
通过观察生殖细胞形态、细胞内部超微结构,将口虾蛄精子发生过程分为3种配子体。
精原细胞:细胞呈不规则卵圆形,大小为(10.0±5.9)μm;细胞核近圆形,大小为 (5.0± 3.2)μm,位于细胞中央。细胞核周围有许多内质网泡,胞质内有少量线粒体 (图9-A、B)。

图6 口虾蛄卵细胞发生过程Fig.6 Oogenesis of mantis shrimp Oratosquilla oratoria
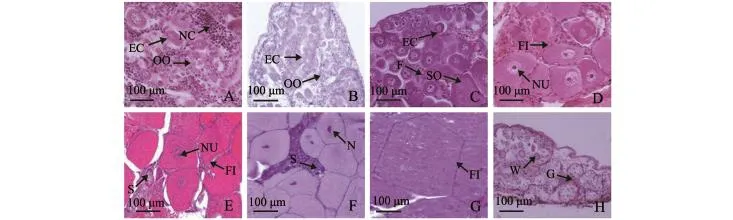
图7 口虾蛄卵巢发育分期Fig.7 Gonadal development of mantis shrimp Oratosquilla oratoria
精母细胞:细胞呈椭圆形,细胞小于精原细胞,大小为 (7.0±2.2)μm;细胞核为 (3.0± 2.8)μm,核为圆形,位于细胞一端,呈极性分布;线粒体大小不一,具双层膜,嵴少,集中分布于细胞的另一端,并有双层膜结构包裹形成 “线粒体区”。精母细胞旁有营养细胞存在,营养细胞呈多边形,较小,大小仅4 μm左右,胞质内有少量内质网、线粒体存在 (图9-C、D、E、F、G)。
精子:精子排列紧密,无鞭毛,呈圆球形或水滴形,细胞膜厚,细胞大小为 (4.0±1.3)μm;细胞核浓缩不规则,染色质致密,位于细胞一端,细胞器逐渐模糊、融合或解离,胞质中只有线粒体存在,线粒体体积由小变大,数量由少变多,嵴较多,并迁移至细胞核的末端 (图9-H)。
基于生殖细胞的类型及数目,将口虾蛄精巢发育分为4个时期。
精原细胞期:精巢未发育,只包含精原细胞(图10-A)。
精母细胞期:此期包含精原细胞、精母细胞,精原细胞区包裹着精母细胞区 (图10-B)。
早期精子期:此期包含精原细胞、精母细胞、精子,精巢从里到外依次是精子、精母细胞、精原细胞,精子比较少 (图10-C)。
精子期:精巢内大量精子存在,也有少量精母细胞存在 (图10-D)。
3 讨论
3.1口虾蛄性比的周年变化
性比是决定种群繁殖力的重要因素之一,本研究中发现,口虾蛄性比全年存在一定的波动变化,除7月份外,其余月份雌雄比例无显著性差异(P>0.05),雌雄比例接近1∶1。7月雌性口虾蛄数量明显多于雄性口虾蛄 (P=0.034),性比达全年最高值 (1.34),其原因可能与雄性口虾蛄大量交尾后死亡有关[17]。类似的,大富潤等[18]的研究同样也发现,口虾蛄性比存在周年波动,且在繁殖期前后有明显变化。因此,这种性比的波动性可能是种群对环境适应性的反应。
3.2口虾蛄繁殖特征
口虾蛄当年即可达性成熟,其生物学最小型大约为8.0 cm[19]。本试验中发现,其生长具有明显的季节特征,夏季和秋季生长最为快速。此外,皮口海域口虾蛄常年均有分布,但1、2月近岸资源量较少,采样困难。这可能与低温期口虾蛄从近岸移动到深水区,营越冬生活有关[20]。本研究中还发现,精巢和卵巢发育均具有不同步性,该现象也普遍存在于沼虾属、原螯虾属中,如日本沼虾Macrobrachium nipponense、克氏原螯虾Procambarus clarkii[21]等。
皮口海域雄性口虾蛄存在2个非常明显的性成熟高峰,基于精巢质量的周年变化推断交尾时间时发现,从3月精巢开始发育,其峰值出现在4月(水温13℃),平均精巢质量为0.166 g,随后精巢质量开始逐渐下降,可能进入交尾阶段,到6月(水温18℃),平均精巢质量最低,为0.099 g,推断此期间第1次交尾结束;此后,精巢质量又再次明显上升,至11月 (水温7℃)达到全年最高值后又开始下降,可能再次进入集中交尾阶段。故初步推测,口虾蛄交尾很可能集中发生在4—6月和11—12月。雄性口虾蛄精巢发育明显不同步性,造成无法准确通过组织切片来证实对交尾时间的推测,未来尚需通过室内人工繁育试验进一步验证。目前,国内外学者所推测的口虾蛄交尾时间主要依赖评估性腺质量的变化,且群体间的交尾时间尚无统一定论。梅文骧等[22]认为,浙江沿海口虾蛄在产卵前不久进行交尾;邓景耀等[23]认为,渤海海区口虾蛄在9—11月份交尾;Hamano[24]研究发现,日本九州口虾蛄在产卵前几个月已经完成交尾;王波等[25]研究表明,口虾蛄为广温广盐性种类,可以适应6~31℃的温度和12~35的盐度变化。当地的水温及生态条件极大地影响其繁殖发育周期。在渔业资源研究中,采样方式也会造成一定判断偏差。本研究中对皮口海域口虾蛄群体进行为期一周年的采样,尽可能规避了采样方式对试验结果所造成的影响。
性腺指数是揭示物种繁殖状况的重要指标,被广泛应用于水产动物繁殖期的判定中,如黄鳝Monopterus albus[26]、中华绒螯蟹 Eriocheir sinensis[27]等。皮口海域雌性口虾蛄性成熟高峰期也有2个,故推测产卵时间可能在5—9月和11—12月。但考虑到实际水温状况及以往口虾蛄人工育苗研究情况[28],繁殖盛期出现在5—9月是合理的,这与本试验组织学观察结果一致,也与刘海映等[10]报道的黑石礁海域口虾蛄产卵时间在6—7月类似。尽管本试验中发现,11月份所采集的口虾蛄样本中均存在性腺成熟 (成熟期)及排完卵 (恢复期)的个体,但考虑到皮口海域冬季水温低,且从当年11月份至翌年3月的5次采样中,并未采集到变态后仔虾的结果,尚无法确定皮口海域口虾蛄在冬季是否可以形成具有实际意义的繁殖种群。
本次采样发现,雌性口虾蛄的性成熟高峰期与交尾时间部分重合,暗示口虾蛄可能具有延后受精和同时受精两种交配模式。根据性腺指数和组织学观察结果,考虑到口虾蛄为2~3年生种类[28],很可能1龄以上口虾蛄在当年5—9月繁殖,未满1龄口虾蛄在冬季11—12月近岸育肥,性腺发育后,翌年5—9月才进入集中产卵期。类似的,鳌虾科克氏原螯虾的繁殖期也仅有1个 (8—9月),10月底后抱卵的雌虾由于水温降低,一直抱卵直至翌年春季才开始孵化[30]。Ohtomi等[8]对东京湾口虾蛄的研究也显示,当地口虾蛄繁殖时期仅有1个,为4—8月,而性成熟高峰期却为2个。徐善良等[31]对浙江北部口虾蛄的研究发现,口虾蛄每年也仅有1个繁殖期,性成熟高峰期分布在7—8月。在甲壳动物中,还有一些种类一年中存在2个繁殖期,如长臂虾科的脊尾白虾Exopalaemon carinicauda(4—5月和8—9月)[32]以及梭子蟹科的锯缘青蟹Scylla serrata(5—6月和8—10月)[33],这暗示甲壳动物在经历千百万年的进化后,具有适应其生存环境的不同生殖策略。
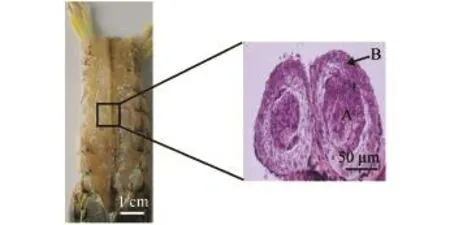
图8 口虾蛄精巢Fig.8 The testis of mantis shrimp Oratosquilla oratoria

图9 口虾蛄精子发生的超微结构观察Fig.9 Ultrastructure during spermatogenesis of mantis shrimp Oratosquilla oratoria
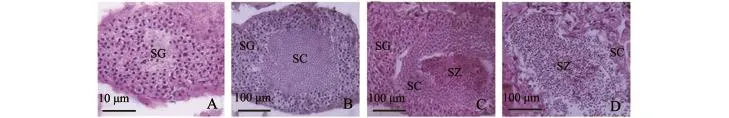
图10 口虾蛄精巢发育分期Fig.10 Testis development of mantis shrimp Oratosquilla oratoria
3.3口虾蛄卵子发生和卵巢的发育分期
生殖细胞的发生是繁殖生物学研究的重要课题,无论是在理论上还是在人工育苗中都具有重要指导意义。对于口虾蛄的卵巢分期,国内外学者意见不一。徐善良等[9]将口虾蛄卵母细胞发育分为4期,缺少未发育期和恢复期。刘海映等[10]对口虾蛄的繁殖周期和生殖细胞发育进行了研究,但未结合性腺发育过程。此外,Jennifer等[34]也做了一些工作,但由于缺乏对同一区域周年研究数据,卵子发生和卵巢发育分期尚不完备,不能完全解释口虾蛄这一特殊物种生活史中复杂的繁殖发育过程。本研究中,以皮口海域口虾蛄为研究对象,将其卵子发生过程分为7种配子体,卵巢发育分为8个时期,完整地阐述了口虾蛄卵巢从未发育至最终排卵的全部过程,首次探明了雌性口虾蛄周年发育规律。
3.4口虾蛄精子发生和精巢发育分期
口虾蛄精巢发育方式与卵巢不同,从外到内发育,且精子极小,仅有卵子的1/100,故结合组织学和透射电镜技术才能揭示整个精子的发生过程。在精子发生方面,精原细胞为卵圆形,由生殖上皮细胞迅速生长、增殖形成。精原细胞在发育过程中,细胞体积明显增大,细胞核逐渐呈卵圆形。这些特征与其他甲壳动物精原细胞的发生类似[35-36]。口虾蛄精母细胞小于精原细胞,同日本沼虾M. nipponense、三疣梭子蟹Portuns trituberculatus的精母细胞形态类似,与脊尾白虾相反。而在成熟精子方面,甲壳动物明显表现出种的特异性。如本试验中观察到的口虾蛄成熟精子呈无鞭毛的圆球形或水滴形,而日本沼虾成熟精子呈图钉形,中国龙虾Panulirus stimpsoni成熟精子呈泡囊形,克氏原螯虾成熟精子呈辐射形。与精子发生的复杂性相比,精巢分期相对简单,几乎所有甲壳种类均具有基本的精原细胞期、精母细胞期和精子期。
本试验中通过透射电镜技术发现,精子发生过程中细胞器的形态结构发生了很大变化。如从精原细胞期到精子成熟,线粒体数量由少到多、体积从小到大。这种现象可能与口虾蛄交配及受精时间不一致,为保证精子存活,需要大量线粒体储存能量有关;同时,口虾蛄精子并无鞭毛结构[37],依靠大量线粒体提供受精能量也是可能的。
综上所述,本研究中通过一周年的采样调查分析,探讨了大连市皮口海域口虾蛄的繁殖规律、生殖细胞发生和性腺发育分期,为丰富口虾蛄的繁殖生物学研究内涵提供了基础资料。
参考文献:
[1]刘瑞玉.中国海洋生物名录[M].北京:科学出版社,2008: 659.
[2]金珊,赵青松.虾蛄[J].生物学通报,2004,39(6):17.
[3]杨德渐,王永良,马绣同,等.中国北部海洋无脊椎动物[M].北京:高等教育出版社,1996:411-412.
[4]朱冬发,王桂忠,李少菁.东方扁虾卵子发生的超微结构[J].水生生物学报,2006,30(4):439-445.
[5]王春琳,朱冬发,蒋霞敏,等.黑斑口虾蛄雄性生殖系统的组织学与超微结构[J].水产学报,2002,26(5):403-410.
[6]Yamazaki M,Fuji A.Reproductive cycle of the mantis shrimp,Squilla oratoria de Haan,in Mutsu Bay[J].Bulletin of the Faculty of Fisheries Hokkaido University,1980,31(2):161-168.
[7]Kodama K,Shimizu T,Yamakawa T,et al.Reproductive biology of the female Japanese mantis shrimp Oratosquilla oratoria(Stomatopoda)in relation to changes in the seasonal pattern of larval occurrence in Tokyo Bay,Japan[J].Fisheries Science,2004,70 (5):734-745.
[8]Ohtomi J,Shimizu M.Spawning season of the Japanese mantis shrimp Oratosquilla oratoria in Tokyo Bay[J].Nippon Suisan Gakkaishi,1988,54(11):1929-1933.
[9]徐善良,王春琳,梅文骧,等.口虾蛄Oratosquilla oratoria(De Huan)性腺特征及卵巢组织学观察[J].浙江水产学院学报,1996,15(1):21-28.
[10]刘海映,谷德贤,姜玉声,等.口虾蛄繁殖周期及生殖细胞发育的研究[J].大连海洋大学学报,2013,28(3):269-272.
[11]胡盼,宫雪,韩雨哲,等.太平洋鳕仔鱼消化系统发育形态学和组织学观察[J].大连海洋大学学报,2015,30(3):243-247.
[12]王春琳,蒋霞敏,赵青松,等.黑斑口虾蛄的卵巢组织学研究[J].动物学杂志,2001,36(4):6-9.
[13]康现江,梁春光,郭明申,等.半滑舌鳎精子发生和精子形成的超微结构[J].动物学报,2008,54(2):356-365.
[14]蒋霞敏,王春琳,赵青松,等.虾蛄的人工养殖技术研究[J].东海海洋,1999,17(2):47-50.
[15]蒋霞敏,王春琳,赵青松,等.黑斑口虾蛄育苗技术研究[J].海洋科学,2000,24(5):17-20.
[16]王艺磊,张子平,李少菁.甲壳动物精子学研究概况:Ⅱ精子发生与精子的生化组成[J].动物学杂志,1998,33(4):52-57.
[17]邓景耀,韩光祖,叶昌臣.渤海对虾死亡的研究[J].水产学报,1982,6(2):119-127.
[18]大富潤,清水誠.東京湾産ツヤコの性比おょび肥満度の季節変化[J].水産増殖,1989,37(2):143-146.
[19]Hamano T,Matsuura S.Egg laying and egg mass nursing behavior in the Japanese mantis shrimp[J].Nippon Suisan Gakkaishi,1984,50(12):1969-1973.
[20]吴耀泉,张宝琳.渤海经济无脊椎动物生态特点的研究[J].海洋科学,1990(2):48-52.
[21]戴颖,巩雪洁,李兵,等.武汉地区克氏原螯虾繁殖期的研究[J].动物学杂志,2008,43(2):21-27.
[22]梅文骧,王春琳,张义浩,等.浙江沿海虾蛄生物学及其开发利用研究报告[J].浙江水产学院学报,1996,15(1):1-8.
[23]邓景耀,程济生.渤海口虾蛄渔业生物学研究[C]//甲壳动物学论文集:第三辑.青岛:青岛海洋大学出版社,1992:36-44.
[24]Hamano T.Mating behavior of Oratosquilla oratoria(de Haan,1844)(Crustacea:Stomatopoda)[J].Journal of Crustacean Biology,1988,8(2):239-244.
[25]王波,张锡烈,孙丕喜.口虾蛄的生物学特征及其人工苗种生产技术[J].黄渤海海洋,1998,16(2):64-73.
[26]刘家芳,张建勋,袁汉文.低蛋白水平饲料对黄鳝性腺指数和性腺发育的影响[J].长江大学学报:自然科学版,2012,9 (3):30-33.
[27]常国亮,成永旭,丁怀宇,等.早熟中华绒螯蟹雄蟹性腺指数、性腺生化成分及脂肪酸组成变化[J].河南师范大学学报:自然科学版,2014,42(1):120-123.
[28]刘海映,姜玉声,邢坤,等.口虾蛄池塘生态育苗方法:CN101703020A[P].2010-05-12.
[29]盛福利,曾晓起,薛莹.青岛近海口虾蛄的繁殖及摄食习性研究[J].中国海洋大学学报,2009,39(S):326-332.
[30]龚世园,吕建林,孙瑞杰,等.克氏原螯虾繁殖生物学研究[J].淡水渔业,2008,38(6):23-25.
[31]徐善良,王春琳,梅文骧,等.浙江北部海区口虾蛄繁殖和摄食习性的初步研究[J].浙江水产学院学报,1996,15(1):30-36.
[32]栗治国,张成松,李富花,等.脊尾白虾的性腺发育及组织结构观察[J].水产学报,2014,38(3):362-370.
[33]李少箐,王桂忠.锯缘青蟹繁殖生物学及人工育苗和养成技术的研究[J].厦门大学学报:自然科学版,2001,40(2):552-565.
[34]Jennifer L,Wortham-Neal J L.Reproductive morphology and biology of male and female mantis shrimp(Stomatopoda:Squillidae)[J].Journal of Crustacean Biology,2002,22(4):728-741.
[35]赵云龙,堵南山,赖伟.日本沼虾精子发生的研究[J].动物学报,1997,43(3):243-248.
[36]黄海霞,谈奇坤,郭延平.秀丽白虾精子发生的研究[J].动物学杂志,2001,36(2):2-6.
[37]堵南山.甲壳动物学:下册[M].北京:科学出版社,1993:374-398.
中图分类号:S917;Q953
文献标志码:A
DOI:10.16535/j.cnki.dlhyxb.2016.03.002
文章编号:2095-1388(2016)03-0237-09
收稿日期:2015-09-29
基金项目:辽宁省科学技术计划项目(2014203016);辽宁省教育厅一般项目 (L2015083)
作者简介:薛梅 (1989—),女,硕士研究生。E-mail:15141193742@163.com
通信作者:刘奇 (1983—),男,博士。E-mail:liuqisunson@dlou.edu.cn一,本研究中,以大连市皮口海域口虾蛄为研究对象,通过对其组织学及超微结构分析,对口虾蛄生殖细胞发生和性腺发育的周年变化规律进行了详细研究,旨在丰富口虾蛄繁殖生物学研究内涵,进而为规模化人工繁育口虾蛄苗种以及对其进行增养殖提供理论指导。
Reproductive biology of mantis shrimp Oratosquilla oratoria in Pikou Bay in coastal Dalian
XUE Mei1,2,YAN Hong-wei3,LIU Hai-ying1,2,LIU Qi1,2,XING Kun1,2,CHEN Lei1,2,LÜ Hai-bo1,2,LU Yu-jie1,2,ZHANG Tao4,CHI Qing-hong5
(1.Center for Marine Ranching Engineering Science Research of Liaoning,Dalian Ocean University,Dalian 116023,China;2.Key Laboratory of Marine Bio-resources Restoration and Habitat Reparation in Liaoning Province,Dalian Ocean University,Dalian 116023,China;3.College of Fisheries and Life Science,Dalian Ocean University,Dalian 116023,China;4.Dalian Tianzheng Co.Ltd.,Dalian 116000,China;5.Fishery Technology Extension Station of Changhai County,Changhai 116500,China)
Abstract:Germ cell genesis and gonadal development were monthly observed in mantis shrimp Oratosquilla oratoria collected in Pikou Bay in coastal Dalian from May 2014 to April 2015 by histological and ultrastructural technology to investigate the reproductive biology of the mantis shrimp population.The results showed that the ratio of female to male was similar to anticipated value 1∶1 in other months except for July(χ2=0.500,P>0.05),range from 0.79 to 1.34,with annual mean value of 1.04±0.05.There was similar tendency of gonadal development between female and male,with two peaks of maturity throughout the year,May and November in female,and April and November in male,early development in male than in female.Oogenesis of mantis shrimp was divided into 7 stages:oogonium,primary oocyte,secondary oocyte,previtellogenic oocyte,vitellogenic oocyte,early mature oocyte and mature oocyte.And ovarian development was divided into non-developmental stage,primary oocyte stage,early growth stage,middle growth stage,late growth stage,pre-maturation stage,maturation stage and finally atretic stage.Correspondingly,the spermatogenesis was divided into three sages:spermatogonium,spermatocyte and sperm,while testis development was divided into four stages:spermatogonium stage,spermatocyte phase,early sperm and sperm phase.The findings provide a first step towards understanding of the reproductive biology of mantis shrimp,benefit for stock enhancement and aquaculture of mantis shrimp in the future.
Key words:Oratosquilla oratoria;gonad;germ cell;histology;ultrastructure
