刨花楠不同相对生长速率下林木叶片碳氮磷的适应特征
2016-07-21张蕾蕾钟全林程栋梁吴永宏张中瑞王卫军
张蕾蕾, 钟全林, 程栋梁, 费 玲, 李 静, 吴永宏, 张中瑞, 王卫军
1 福建师范大学地理科学学院, 福州 350007 2 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 3 福建师范大学地理研究所, 福州 350007 4 江西省安福县谷源山林场, 安福 343200
刨花楠不同相对生长速率下林木叶片碳氮磷的适应特征
张蕾蕾1, 钟全林1,2,3,*, 程栋梁1,2,3, 费玲1, 李静1, 吴永宏1, 张中瑞1, 王卫军4
1 福建师范大学地理科学学院, 福州350007 2 湿润亚热带山地生态国家重点实验室培育基地, 福州350007 3 福建师范大学地理研究所, 福州350007 4 江西省安福县谷源山林场, 安福343200
摘要:分别对9年生与13年生刨花楠林木叶片氮磷养分之间关系及林木生物量相对生长速率与叶片碳氮磷化学计量比关系进行分析,探讨不同相对生长速率下的林木叶片N、P养分适应特征,并检验相对生长速率假说理论对刨花楠树种的适应性。结果表明:两种年龄刨花楠林木生物量相对生长速率、叶片C、N、P含量及其计量比值均存在显著差异;同一年龄的林木叶片N、P之间存在显著相关性,二者具有协同相关性;9年生林木叶片P含量及C∶P、N∶P与生物量相对生长速率呈二次曲线相关,而13年生林木叶片N、P含量及C∶N、C∶P、N∶P则与生物量相对生长速率均呈线性相关。研究表明,在能满足植物生长所需养分供给的土壤环境中,叶片N、P含量与林木相对生长速率间呈线性正相关,但当土壤中养分供应满足不了植物高速生长时,植物则会对有限的养分资源进行适应性调整。
关键词:生长速率假说;叶片氮磷比;林龄;刨花楠
N、P元素是控制陆地生态系统中植物生产力和代谢功能的关键因子[1],有机体内C∶N∶P比值及范围反映其对环境的适应机制及特征,并在一定程度上决定着有机个体或群落的稳定性[2]。生长速率是反映植物生活史对策的核心参数,并与之养分适应特征密切相关[3]。植物有机个体通过N∶P比值的动态变化反映其相对生长速率(RGR),理论基础在于N、P元素在蛋白质及核糖体中分配的内在联系[4- 5]。对水生生态系统中大量异养浮游生物[3,6]、丝状菌[7]等N∶P比值与相对生长速率关系的研究认为,P是控制有机体生长发育的最主要因子,并且这一观点构成生长速率假说理论的基本观点[3]。由于自养型生物与异养型生物获取影响生长的营养元素的途径存在差异,因此Årgen[8]针对生长速率假说理论在自养型生物方面的应用进行修正并建立理论模型,发现有机体在N素限制条件下,叶片N∶C与RGR呈线性上升;受P限制时P∶C与RGR呈曲线上升;而在受到N、P同时限制时,叶片N∶P则随RGR增大呈先上升至最大值后逐渐下降趋势。Cernusak等[9]和严正兵等[10]通过对不同陆地植物幼苗的控制施肥实验得到的关于C、N、P及其比值与RGR之间的关系结果与Årgen的研究结论基本一致。但Matzek等[11]对14种针叶树苗木的控制施肥实验得到的结果却为:叶片N∶P比值与相对生长速率之间不存在显著相关性。另外,有学者考虑到N与植物初级生产力[12]、碳水化合物合成[13]及呼吸作用[14]之间有密切联系,认为植物体内N含量决定植物的相对生长速率并与之呈线性正相关。Peng等[5]以我国亚热带地区植物为对象进行研究发现,不同物种N含量、N:C及N∶P比值均随RGR升高而增大,由此他认为在养分供应充足的情况下,有机体内N含量能更好地充当反映RGR高低的指标。土壤发育程度、气候区域性特征以及人类活动增加了陆地生态系统的复杂性,使得在对不同生活型以及生长环境差异较大的物种进行生长速率假说理论论证时,通常会产生不一致的结果。从上述相关文献可看出,以往多数学者以控制施肥实验方式对生长速率大、生长周期短的草本植物或林木幼苗进行研究,而对森林生态系统中乔木林的研究相对较少。生长速率假说理论是否适用于群落结构更为复杂的常绿阔叶林生态系统?林木是如何通过N∶P比值与相对生长速率之间的关系调控其生长发育过程?这些都还需作进一步研究。因此,论文以刨花楠人工林为对象,针对这些问题,开展相关研究。
刨花楠(Machiluspauhoi)是我国亚热带优良的常绿阔叶树种,幼年喜阴耐湿,成年喜光喜湿,适应性强,主要分布在广东、广西、江西、福建和浙江等地。本文分别以江西省永新县七溪岭林场9年生刨花楠人工林及安福县谷源山林场13年生刨花楠人工林为对象,通过对不同林龄刨花楠叶片N、P养分之间的协同关系以及RGR与叶片碳氮磷化学计量比关系进行分析,拟解决两个主要问题:1)随林龄增加,林木叶片N、P含量与相对生长速率间是否保持线性相关;2)生长在不同生长环境中的刨花楠如何调控N、P含量以满足其生长需求。研究结果旨在验证并丰富生长速率假说理论在我国亚热带常绿阔叶林中的应用实践,并为开展刨花楠人工林高效经营技术(如氮磷配方施肥技术),探讨亚热带常绿阔叶树种在生长过程中的养分需求规律等提供理论参考。
1材料与方法
1.1研究地概况
试验地分别位于江西省永新县七溪岭林场9年生刨花楠人工林(114°16′42.7″E、26°5′7.4″N)和安福县谷源山林场13年生刨花楠人工林(114°29′51.3″E、27°17′19.5″N),造林苗木均为1.5年生实生苗,两地均属于湿润亚热带季风气候。永新县年均降水量1457.15 mm,年均降雨日为166 d,年均气温约18.13 ℃,7月份平均气温27.15 ℃,1月份平均气温7 ℃。安福县年均降水量1553 mm,年均降雨日166 d,年均气温17.7 ℃,7月平均气温28.9 ℃,1月平均气温5.9 ℃。两林地土壤均为红壤,偏酸性,造林密度均为3300株/ hm2。至调查时9年生人工林现实密度2417株/ hm2,林木胸径、树高平均值分别为9.34 cm、6.59 m;13年生人工林现实密度为1375株/ hm2,林木平均胸径13.21 cm、平均树高8.21 m。
1.2样地调查与取样
2012年7月12日至14日,分别在两刨花楠人工林中各典型选设3个代表性样地(20 m×20 m),各样地间距≥50 m。将每个样地划分为4个10 m×10 m的样方。对每个样方中胸径≥3 cm刨花楠进行标号、挂树号牌,进行每木检尺,记录其胸径、树高。按照胸径大小采用上限排外法,以2 cm为一径阶(如胸径3—4.9 cm为4径阶,5—6.9 cm为6径阶,依次类推),按径阶将各样木进行分组。分别径阶各选取3株接近径阶平均木的样本(做好标记,以便后续连续采样分析),用高枝剪截取上层外圈向光面小枝(每株取3根);从每根小枝上典型摘取完全成熟的生长健康的当年生叶片5片(等间距采摘),分别样地将所摘取的同径阶样本叶进行混合、装袋,带回实验室75 ℃恒温烘干后,测定其全C、全N、全P含量。2013年1月1日至7日,对样地中刨花楠的胸径与树高进行复查,并按7月份采样方法采集原径阶样木的当年生上层叶片并测定养分含量。
另在每个样地中无干扰处典型设置3个土壤剖面,按照0—10、10—20、20—40 cm和40—60 cm的层次,环刀取土并带回室内进行养分分析(全C、全N、全P)。土壤养分特征见表1。

表1 9年生与13年生刨花楠林地土壤年均C、N、P含量及比值(平均值±标准差)
同列不同小字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)
1.3样品测定方法及数据处理
叶片、土壤全C、全N含量使用Vario ELⅢ元素分析仪测定,全P含量采用高氯酸消煮+连续流动分析仪(Skalar San++)测定。文中叶片、土壤全C、全N、全P含量及化学计量比值均为两期调查的平均值,采用SPSS 17.0统计软件对测定数据进行分析、统计(α=0.05),并用Origin 8.0软件绘图。测定生物量相对生长速率:
生物量相对生长速率=ln(Mt/M0)/t
式中,M0为7月份(初始)调查地上生物量,Mt为1月份(终期)调查地上生物量,t为二者间隔时间。本文中地上生物量根据实测树高和胸径值按文献[15]中的生物量模型进行拟合。
2结果与分析
2.1刨花楠叶片C、N、P养分含量及生物量相对生长速率
通过对9年生与13年生刨花楠林木叶片平均C、N、P含量及其比值、相对生长速率(RGR)进行T-检验得出:不同林龄叶片平均养分含量及其比值、RGR均存在极显著差异(P<0.01)(表2)。13年生林木叶片平均C、N、P含量及C∶N、C∶P均高于9年生刨花楠林木,而RGR与叶片平均N∶P低于9年生林木。
相关性分析表明,9年生与13年生林木叶片中平均N含量与P含量之间均呈极显著线性正相关(P<0.01)(图1);而两林地中,叶片平均C含量与N、P之间无显著相关性。说明在不同林龄中,刨花楠叶片N、P养分对环境的响应一致。
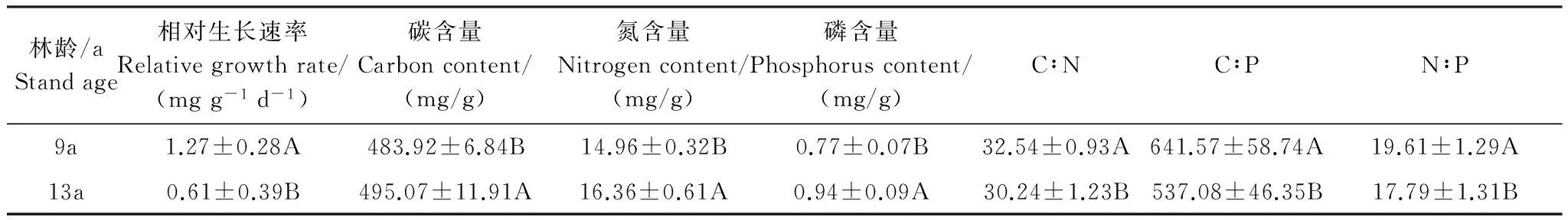
表2 不同林龄刨花楠叶片养分含量及比值(平均值±标准差)

图1 不同林龄刨花楠叶片氮、磷含量相关性 Fig.1 The correlation bewteen N and P content of Machilus pauhoi leaves in different stand ages
2.2林木生物量相对生长速率与叶片碳氮磷化学计量比关系
由图2看出,两林龄中刨花楠叶片C含量与RGR间无显著相关性,而N、P含量与RGR之间关系则呈现出差异。13年生林木叶片N、P含量均随RGR增大呈极显著线性正相关(P<0.01)。一方面可反映出林木土壤养分偏高;另一方面说明,虽13年生林木相对生长速率低但其生物量总量较大,故其生长对N、P需求量偏高。9年生林木叶片N含量与RGR间无显著相关性,P含量与RGR之间呈先上升后下降的二次抛物线关系(图2),说明植物在面对贫瘠环境时对养分利用策略进行了调整。

图2 刨花楠叶片碳、氮、磷含量与相对生长速率的关系Fig.2 The correlation bewteen leaves C,N,P content and RGR of Machilus pauhoi in different stand ages
由图3看出,13年生刨花楠叶片C∶N、C∶P及N∶P与RGR呈显著线性负相关,均随RGR增大而下降。9年生林木叶片C、N、P化学计量特征与RGR之间的关系则呈现不同,叶片N∶C与RGR间无显著相关性,而C∶P及N∶P与RGR间均呈先下降后上升的二次曲线关系。当RGR<1.8mg g-1d-1时,两林地中林木叶片C∶P、N∶P均随相对生长速率增大而减小。

图3 刨花楠叶片碳、氮、磷化学计量比与相对生长速率的关系Fig.3 The correlation bewteen leaves stoichiometric rations and RGR of Machilus pauhoi in different stand ages
3讨论
叶片N、P养分含量是反映土壤养分供给能力及植物生长状况的重要指标之一[16- 17]。两刨花楠人工林叶片平均N、P含量均低于我国753种陆生植物[18]及南北样带森林生态系统中102种优势植物[19]的平均N、P含量,反映了刨花楠叶片N、P含量相对偏低,其原因可能是受两林地土壤养分供应所限。N、P之间的关系则是由植物自身的元素生物化学过程及代谢活动决定[20],是对养分吸收和储存能力的响应。本研究中,9年生和13年生刨花楠林木叶片N、P含量间均存在显著线性正相关(图1),即N、P元素间存在良好的协同性,这与众多学者的研究结论一致[21- 23]。研究发现,9年生林木叶片N、P之间的协同作用强于13年生刨花楠林木,这在一定程度上不仅是对两林地土壤N、P含量存在差异性的响应(表1),也反映了植物不同年龄阶段其N、P养分利用策略的改变[24]。
生长速率假说理论将细胞生物化学及生理特征(如RNA与蛋白质分配对个体生长的影响)与有机体所反映的生态现象(如有机体N∶P反映的养分循环特征)紧密联系[11]。本研究中,13年生与9年生林木叶片N、P养分含量及RGR方面的显著差异性为研究不同N、P含量水平下,植物对RGR的响应机制以及在不同RGR水平下,植物如何调整和利用有限的N、P养分等提供了对比分析实证。
9年生与13年生刨花楠叶片N含量、C∶N存在显著差异,并与RGR之间的关系呈现不同。从图2(A)、图3(A)中看出,13年生叶片N含量较高,N、C∶N与相对生长速率呈线性相关;但9年生林木,在叶片N含量较低时,其N含量与RGR间无显著相关,说明在此生长阶段适当的添加N肥,提高N素供应对维持和促进刨花楠生长具有重要作用。这一结论与Peng[5]及Yu[25]等研究结果具有一致性。
叶片C∶P与林木相对生长速率之间的关系主要受P含量影响(图2B、图3B)。13年生刨花楠叶片P含量相对较高,并随RGR增大而上升,与生长假说理论一致。9年生林木叶片P含量则相对较低,与RGR之间的关系复杂。相关研究[5,9,25]均表明,在P素受限条件下提高P素供应,植物叶片P、P∶C与相对生长速率呈正比;不受P限制的情况下提高P素供应,叶片P及P∶C与RGR无显著相关性。产生后者现象的原因,一方面是供给植物生长和代谢的P元素饱和后,植物会将多余的P元素转移到其他组织中利用或储存;另一方面,当环境中P供给过量时,会引起其它元素的代谢紊乱,造成植物呼吸作用过强并消耗大量的糖分和能量,最终对植物生长产生抑制作用[26]。图2B看出,在RGR相对较低时(<1.8 mg/g/d),13年生及9年生林木叶片P含量(抛物线左侧)均随RGR增大呈上升趋势。说明在乔木生长速率较低时,即使生境对P素供给有限也能满足植物生长。当RGR持续增大时,环境对P素供给不足问题加剧,这时植物会通过调整养分利用策略以应对快速生长的要求(如,9年生叶片P含量与RGR拟合曲线的右侧)。该结果表明,在养分供应充足情况下,植物因获得充足的养分而快速生长;而在养分限制情况下,植物会通过提高单位养分利用效率以适应较高的生长速率[11]。9年生林木RGR较高、叶片P含量较低,则不仅反映其个体快速生长消耗了体内的大量P素,也反映其对P素的利用效率相对较高(与13年生刨花楠比较)。
叶片N∶P比值变化是植物内部调节机制与外界环境之间权衡的表现[9,12]。两地叶片N∶P与各自RGR的关系不同,但二者仍存在共性。RGR相对较低时(<1.8 mg/g/d),9年生与13年生林木叶片N∶P与相对生长速率均呈负相关,叶片N∶P下降与提高植物代谢速率与养分循环速率,促进植物生长有密切联系[27]。但在不同土壤养分环境及不同生长速率阶段,植物对养分的吸收是具有选择性的[5,25]。相对动物而言,植物在N、P养分贫瘠的土壤环境中具有提高养分储存的能力以支撑N∶P之间的稳定性[28],尤其是P元素在植物体内的分配方式。P元素进入细胞后,首先将P投入到磷脂、核糖核酸(RNA)、脱氧核糖核酸(DNA)以及三磷酸腺苷(ATP)等化合物的合成中,以保证植物正常的生长和代谢功能;剩余部分以磷酸盐形态储存于液泡中或运往植物其它组织中以被利用[26]。当环境中P元素供给不足时,植物会重新利用液泡中储存的P,或利用韧皮部重新分配在其它组织中的P元素。9年生林木在其RGR较大时,其叶片N∶P与林木RGR呈正相关,这正是由于植物对N、P养分调整和权衡的结果。因此,相对于环境变化而言,林木RGR与其自身养分吸收和储存能力更为密切。
植物RGR与其碳氮磷化学计量比值间的关系因生活型、生长阶段和养分条件而出现差异[5]。森林生态系统植物对复杂生境的适应能力强,植物可以通过体内养分循环调节机制以及生长与养分耐受之间的权衡[25]以适应相对贫瘠的环境,因此高等植物N∶P与RGR间的关系并不一定仅仅是简单的线性相关。本文对不同林龄刨花楠叶片N、P养分含量与相对生长速率的研究发现,研究区林木叶片N、P养分含量相对偏低(与其它区域植物比较),但N、P两营养元素含量间具良好的协同性;在能满足植物生长所需养分供给的土壤环境中,叶片N、P含量与林木相对生长速率间呈线性正相关,但当土壤中养分供应满足不了植物高速生长时,植物则会对有限的养分资源进行适应性调整。
参考文献(References):
[1]Marklein A R, Houlton B Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytologist, 2012, 193(3): 696- 704.
[2]Lovelock C E, Feller I C, Ball M C, Ellis J, Sorrell B. Testing the growth rate vs. geochemical hypothesis for latitudinal variation in plant nutrients.Ecology Letters, 2007, 10(12): 1154- 1163.
[3]Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L J. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540- 550.
[4]Hessen D O, Jensen T C, Kyle M, Elser J J. RNA responses to N- and P- limitation: reciprocal regulation of stoichiometry and growth rate inBrachionus. Functional Ecology, 2007, 21(5): 956- 962.
[5]Peng Y H, Niklas K J, Sun S C. The relationship between relative growth rate and whole- plant C: N: P stoichiometry in plant seedlings grown under nutrient- enriched conditions. Journal of Plant Ecology, 2011, 4(3): 147- 156.
[6]Hessen D O, Ventura M, Elser J J. Do phosphorus requirements for RNA limit genome size in crustacean zooplankton?. Genome, 2008, 51(9): 685- 691.
[7]Grimmett I J, Shipp K N, Macneil A, Bärlocher F. Does the growth rate hypothesis apply to aquatic hyphomycetes?.Fungal Ecology, 2013, 6(6): 493- 500.
[8]Ågren G I. The C: N: P stoichiometry of autotrophs-theory and observations. Ecology Letters, 2004, 7(3): 185- 191.
[9]Cernusak L A, Winter K, Turner B L. Leaf nitrogen to phosphorus ratios of tropical trees: experimental assessment of physiological and environmental controls. New Phytologist, 2010, 185(3): 770- 779.
[10]严正兵, 金南瑛, 韩廷申, 方精云, 韩文轩. 氮磷施肥对拟南芥叶片碳氮磷化学计量比特征的影响. 植物生态学报, 2013, 37(6): 551- 557.
[11]Matzek V, Vitousek P M. N: P stoichiometry and protein: RNA ratios in vascular plants: an evaluation of thegrowth-rate hypothesis. Ecology Letters, 2009, 12(8): 765- 771.
[12]Ågren G I. Stoichiometry and nutrition of plant growth in natural communities. Annual review of ecology, evolution, and systematics, 2008, 39: 153- 170.
[13]Vrede T, Dobberfuhl D R, Kooijman S A L M, Elser J J. Fundamental connections among organism C: N: P stoichiometry, macromolecular composition, and growth.Ecology, 2004, 85(5): 1217- 1229.
[14]Reich P B, Tjoelker M G, Pregitzer K S, Wright I J, Oleksyn J, Machado J L. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants.Ecology Letters, 2008, 11(8): 793- 801.
[15]钟全林, 张振瀛, 张春华, 周海林, 黄志强. 刨花楠生物量及其结构动态分析. 江西农业大学学报, 2001, 23(4): 533- 536.
[16]Zhang L X, Bai Y F, Han X G. Application of N: P stoichiometry to ecology studies. Acta Botanica Sinica, 2003, 45(9): 1009- 1018.
[17]Güsewell S. N: P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243- 266.
[18]Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.New Phytologist, 2005, 168(2): 377- 385.
[19]任书杰, 于贵瑞, 陶波, 王绍强. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究. 环境科学, 2007, 28(12): 2665- 2673.
[20]Jiang C M, Yu G R, Li Y N, Cao G M, Yang Z P, Sheng W P, Yu W T. Nutrient resorption of coexistence species in alpine meadow of the Qinghai-Tibetan Plateau explains plant adaptation to nutrient-poor environment.Ecological Engineering, 2012, 44: 1- 9.
[21]Reich P B, Oleksyn J, Wright I J, Niklas K J, Hedin L, Elser J J. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes.Proceedings of the Royal Society B: Biological Sciences, 2010, 277(1683): 877- 883.
[22]Baraloto C, Paine C E T, Poorter L, Beauchene J, Bonal D, Domenach A M, Hérault B, Patio S, Roggy J C, Chave J. Decoupled leaf and stem economics in rain forest trees.Ecology Letters, 2010, 13(11): 1338- 1347.
[23]Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001- 11006.
[24]Townsend A R, Cleveland C C, Asner G P, Bustamante M M C. Controls over foliar N: P ratios in tropical rain forests.Ecology, 2007, 88(1): 107- 118.
[25]Yu Q, Wu H H, He N P, Lü X T, Wang Z P, Elser J J, Wu J G, Han X G. Testing the growth rate hypothesis in vascular plants with above-and below- ground biomass.PLoS One, 2012, 7(3): e32162.
[26]陆景陵. 植物营养学(上册). 北京: 中国农业大学出版社, 2003: 23- 48.
[27]Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapln T, Cornellssen J H C, Dlemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821- 827.
[28]Elser J J, Watts T, Bitler B, Markow T A. Ontogenetic coupling of growth rate with RNA and P contents in fivespecies ofDrosophila. Functional Ecology, 2006, 20(5): 846- 856.
Biomass relative growth rate ofMachiluspauhoiin relation to leaf carbon, nitrogen, and phosphorus stoichiometry properties
ZHANG Leilei1, ZHONG Quanlin1,2,3,*, CHENG Dongliang1,2,3, FEI Ling1, LI Jing1, WU Yonghong1, ZHANG Zhongrui1, WANG Weijun4
1CollegeofGeographicalSciences,FujianNormalUniversity,Fuzhou350007,China2StateKeyLaboratoryBreedingBaseofHumidSubtropicalMountainEcology,Fuzhou350007,China3InstituteofGeography,FujianNormalUniversity,Fuzhou350007,China4GuyuanshanForestFarmofAnfuCounty,Anfu343200,China
Abstract:To explore the effects of leaf nitrogen (N) and phosphorus (P) on biomass relative growth rate (RGR) and to test the growth rate hypothesis (GRH) of plant ecological stoichiometry, two stands (9- and 13-year-old plants, respectively) of Machilus pauhoi were selected. The leaf nutrient contents (C, N, P) and biomass RGR were measured at different growth periods. The results showed that the stand age had significant effect on RGR and leaf C, N, P stoichiometry. Specifically, leaf nitrogen contents were significantly correlated with leaf phosphorus contents within the two differently aged M. pauhoi stands. Quadratic correlation was observed between the RGR and leaf P content, C∶P, and N∶P ratios in 9-year-old stand. In contrast, growth rates were linearly correlated with leaf N and P stoichiometry in 13-year-old stands. Studies had shown that in the environment that meets the needed plant nutrition, leaf N and P showed a positive linear correlation with the relative growth rate of trees; however, when the nutrient content in the environment was inadequate for the fast plant growth, the plants underwent adaptive adjustment to the limited nutrient resources.
Key Words:growth rate hypothesis; leaf N∶P; stand ages; Machilus pauhoi
基金项目:国家自然科学基金项目(31170596, 31170374, 31370589); 国家农业科技成果转化资金项目(2011GB2C400005); 福建省科技厅重大项目(2014N5008); 福建省种业创新项目(2014S1477-4)
收稿日期:2015- 01- 16; 网络出版日期:2015- 08- 26
*通讯作者
Corresponding author.E-mail: qlzhong@126.com
DOI:10.5846/stxb201501160136
张蕾蕾, 钟全林, 程栋梁, 费玲, 李静, 吴永宏, 张中瑞, 王卫军.刨花楠不同相对生长速率下林木叶片碳氮磷的适应特征.生态学报,2016,36(9):2607- 2613.
Zhang L L, Zhong Q L, Cheng D L, Fei L, Li J, Wu Y H, Zhang Z R, Wang W J.Biomass relative growth rate ofMachiluspauhoiin relation to leaf carbon, nitrogen, and phosphorus stoichiometry properties.Acta Ecologica Sinica,2016,36(9):2607- 2613.
