基于控制授粉技术的玉米弱势粒发育与库特征的关系
2016-07-14梁红伟高聚林王志刚于晓芳孙继颖胡树平振余少波李雅剑内蒙古农业大学农学院内蒙古呼和浩特010019
梁红伟 高聚林 王志刚于晓芳 孙继颖 胡树平 王 振余少波 李雅剑内蒙古农业大学农学院,内蒙古呼和浩特 010019
基于控制授粉技术的玉米弱势粒发育与库特征的关系
梁红伟 高聚林 王志刚*于晓芳 孙继颖 胡树平 王 振余少波 李雅剑
内蒙古农业大学农学院,内蒙古呼和浩特 010019
摘 要:明确弱势粒败育和灌浆受限与其库容量或库活性关系,对于探讨弱势粒调控途径、实现密植群体产量挖潜具有重要意义。本研究以典型玉米杂交种郑单 958和先玉 335为材料,在控制授粉条件下(不完全授粉IcP、完全授粉CP),比较成功发育弱势粒(IcP处理)和发育不良弱势粒(CP处理)的库容量和库活性及籽粒灌浆参数的差异。结果表明,不同控制授粉处理下,玉米弱势粒胚乳细胞增殖过程和最大胚乳细胞数无显著差异;IcP处理弱势粒可溶性酸性蔗糖转化酶(SAI)活性显著高于CP处理,平均差异和最大差异分别达12.6%和21.8%,且实测百粒重、籽粒终极生长量、最大灌浆速率和平均灌浆速率皆表现为IcP处理高于CP处理。可见,玉米果穗顶部弱势粒败育或灌浆停滞不受其库容量的限制,籽粒形成期的库活性是弱势粒败育或灌浆受限的核心限制因子。
关键词:玉米;弱势粒;控制授粉;库容量;库活性
本研究由国家自然科学基金项目(31301265,31460329),内蒙古自治区高等学校青年科技英才支持计划项目(NJYT-14-B04)和国家科技支撑计划项目(2011BAD16B13,2012BAD04B04,2013BAD07B04)资助。
This study was supported by the National Natural Science Foundation of China (31301265,31460329),the Program for Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region (NJYT-14-B04),and the National Key Technology Support Program of China (2011BAD16B13,2012BAD04B04,2013BAD07B04).
第一作者联系方式∶ E-mail∶ lxflhw2008@163.com,Tel∶ 15848179322
URL∶ http∶//www.cnki.net/kcms/detail/11.1809.S.20160321.1056.010.html
增加种植密度是生产上提高玉米产量的重要途径,但增密往往造成单株穗粒数的大幅降低而制约玉米产量的提高。研究表明,密植条件下玉米个体穗粒数的降低主要源于果穗顶部弱势粒的早期败育和中后期的灌浆受限[1]。弱势粒不能正常发育或灌浆不充实造成的玉米产量损失在 10%以上[2],促进弱势粒建成与灌浆是玉米产量挖潜的重要途径之一[2-4]。杨建昌等[5]认为,作物籽粒能否有效灌浆与作物籽粒库特征即籽粒库对同化物质的容纳和吸收能力有关。作物籽粒库特征可以用库强度表示,分为库容量和库活性两个方面[6-7]。Ho等[8]认为,库容量即胚乳细胞数和胚乳细胞大小,是物理性限制因子,与胚乳细胞分裂过程有关;库活性则是库器官卸载向其输入的同化物质的能力,在玉米籽粒形成期,参与蔗糖降解的酶主要是可溶性酸性蔗糖转化酶(SAI),可以作为库强度的重要衡量指标[9-11]。徐云姬等[12-13]对玉米果穗不同部位籽粒的研究认为,与果穗中部籽粒相比,上部籽粒轻既与胚乳细胞增值速率小,导致其胚乳细胞数少有关,也与蔗糖降解和淀粉合成的相关酶活性较低有关。但关于成功授粉的弱势籽粒仍然发生败育或灌浆停滞,其受库容量或库活性的限制尚无明确结论,回答这一问题需要对比分析败育弱势粒与成功发育弱势粒形成期的库容量和库活性。基于此,本研究在控制授粉条件下,创造成功发育弱势粒(IcP处理)和发育不良弱势粒(CP处理),并比较分析其库容量、库活性和灌浆动态的差异,旨在揭示弱势粒败育(或灌浆停滞)与其库特征的关系,为探讨弱势粒调控途径,探索密植群体产量挖潜途径提供理论依据。
1 材料与方法
1.1 试验地概况
2014年在包头市土默特右旗萨拉齐(Salaqi,40°28′ N,110°28′ E)和鄂尔多斯市达拉特(Dalate,40°40′ N,110°1′ E)试验基地同步开展两点试验。萨拉齐试验点前茬为玉米,土质为沙壤土,0~30 cm土壤含有机质24.5 g kg-1、碱解氮21.2 mg kg-1、速效磷26.7 mg kg-1、速效钾120.4 mg kg-1,pH 7.8;生育期内总降雨量为417.1 mm,≥10℃活动积温2896.5℃,太阳辐射总量3450.1 MJ m-2。达拉特试验点的前茬为玉米,土质为沙土,0~30 cm土壤含有机质15.6 g kg-1、碱解氮20.5 mg kg-1、速效磷24.8 mg kg-1、速效钾86.3 mg kg-1,pH 7.3;生育期内总降雨量为356.5 mm,≥10℃活动积温3051.9℃,太阳辐射总量3661.9 MJ m-2(图1)。
1.2 试验设计
以典型玉米杂交种郑单958和先玉335为材料,种植密度7.5万株 hm-2,行距60 cm,完全随机排列,3次重复,共6个小区,小区面积112 m2(8 m × 14 m)。各处理均以P2O5105 kg hm-2、纯氮40 kg hm-2(磷酸二铵)、K2O 40 kg hm-2(硫酸钾)作为底肥于春播前旋耕入土,于拔节期追施纯氮 150 kg hm-2(尿素),其他管理同当地大田生产。
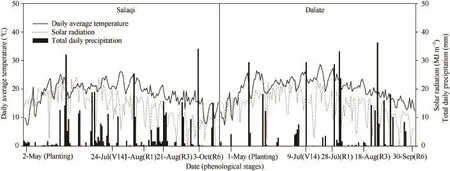
图1 2014年萨拉齐和达拉特试验点玉米生育期内降雨量、日平均温度和日辐射量Fig. 1 Total daily precipitation,daily average temperature and solar radiation during the whole growth period of maize in Salaqi and Dalate in 2014
采用冯汉宇等[14]的方法控制授粉。在玉米抽丝前,在各小区内选取生长整齐一致、有全田代表性的植株80~100株挂牌标记并将其雌穗套袋,待果穗全部花丝完全抽出后,选取一半植株的果穗作为不完全授粉处理(IcP,果穗顶部和中部小花正常授粉,下部花丝不授粉),另一半植株的果穗作为完全授粉处理(CP,玉米果穗全部花丝同步授粉),分别建成弱势籽粒和发育不良弱势籽粒(败育或灌浆停滞)。
1.3 测定指标及方法
从授粉后开始每隔6 d选取不完全授粉和完全授粉果穗各5个,按籽粒粒位分别取2~10环籽粒(从果穗最顶端数起)为上部弱势籽粒,取20~30环为中部籽粒。将各部位分别脱粒200粒左右混匀后,随机选出25粒左右,放入装有卡诺固定液(无水乙醇∶冰醋酸=3∶1溶液)的青霉素小瓶中固定,24 h后置70%乙醇溶液中4℃保存,用于胚乳细胞数的观察和计数。之后,将剩余籽粒随机选出100粒,测定其鲜重,在105℃杀青30~60 min,80℃烘干至恒重,测定百粒干重,用以计算灌浆特征参数;其余籽粒置-80℃超低温冰箱保存,用于可溶性酸性蔗糖转化酶活性测定。
参考张祖建等[15]和王晓燕等[16]的方法分离和计数胚乳细胞,参照徐云姬等[12]和周竹青等[17]方法用Richards方程[18-21]拟合籽粒灌浆和胚乳细胞的增殖动态。参照高俊凤[22]的方法测定可溶性酸性蔗糖转化酶(SAI)活性,称取1 g玉米籽粒于预冷的研钵中,加入5 mL 0.1 mol L-1的Tris-HCl (pH 7.0)缓冲液,研磨后于4℃离心10 min (8000 × g),之后取3 mL上清液于透析袋中密封好,放入25 mmol L-1Tris-HCl (pH 7.0)缓冲液中透析9 h,透析后将酶液定容至5 mL,取50 µL于具塞试管中,加入含50 mmol L-1蔗糖的0.1 mol L-1乙酸-乙酸钠反应液(pH 4.8) 0.95 mL,于30℃水浴中反应10 min,再沸水浴3 min中止反应;用失活酶液作对照,分别向各反应试管中加入1 mL 3,5-二硝基水杨酸试剂,沸水浴5 min后冷却至室温,于540 nm下测定其吸光值,通过标准曲线求其对应葡萄糖量。SAI活性(mg Glucose g-1FW h-1) = (C × Vt FW × t × Vs) × n,式中C为葡萄糖量(mg),Vt为提取酶液总体积(mL),FW为籽粒鲜重(g),t为反应时间(h),Vs为测定时取用酶液体积(mL),n为提取液稀释倍数。
1.4 数据分析
采用Microsoft Excel 2003、Origin8统计分析数据,SPSS 17.0中 One-way ANOVA进行方差分析,SigmaPlot 12.0作图。
2 结果与分析
2.1 弱势粒胚乳细胞增殖动态
由图 2可见,不同控制授粉处理下的玉米果穗顶部籽粒胚乳细胞增殖趋势一致,整体呈“S”型曲线变化。不同品种和不同试验点间无显著差异,籽粒形成期两处理的胚乳细胞数目相当,至灌浆初期IcP处理(成功发育弱势粒)的胚乳细胞数略高于 CP处理(发育不良弱势粒),但差异不显著。不同控制授粉处理下,玉米弱势粒胚乳细胞数目的增殖动态和速率、最大胚乳细胞数目(A)、最大胚乳细胞增殖速率(Gmax)、平均胚乳细胞增殖速率(Gmean)和胚乳细胞活跃增殖期(D)均表现为IcP处理略高于CP处理,但差异不显著;胚乳细胞达到最大增殖速率的时间(Tmax)总体表现为CP处理略高于IcP处理,但处理间也无显著差异(表1)。说明无论弱势粒最终发育成功与否,其籽粒形成期的胚乳细胞分裂过程都未受到显著影响,库容量不是弱势籽粒败育或灌浆停滞的制约因子。
2.2 果穗不同部位籽粒的最大库容比较
不同控制授粉处理下,果穗中部强势粒和顶部弱势粒的最大胚乳细胞数均无显著差异,但从绝对值来看,IcP处理顶部弱势粒的最大胚乳细胞数较CP处理的多14.3% (先玉335)和11.9% (郑单958)(图3)。
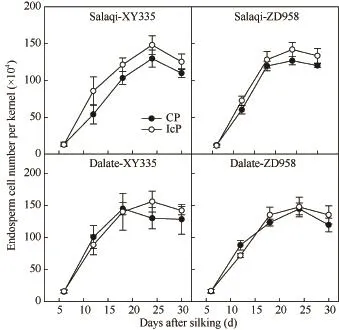
图2 不同控制授粉处理下玉米果穗弱势籽粒胚乳细胞数增殖动态Fig. 2 Dynamics of endosperm cell number of inferior kernels of maize under different pollination treatments
无论是IcP处理还是CP处理,籽粒最大胚乳细胞数均表现为中部籽粒大于上部籽粒,但不同品种间略有差异。先玉335在 IcP处理下中部籽粒最大胚乳细胞数与顶部弱势粒差异不显著,而 CP处理下中部籽粒最大胚乳细胞数显著高于顶部弱势粒,增幅达 25.9%;郑单 958在两种处理下其中部籽粒最大胚乳细胞数皆显著高于顶部弱势粒,增幅分别为22.9%和33.5%。
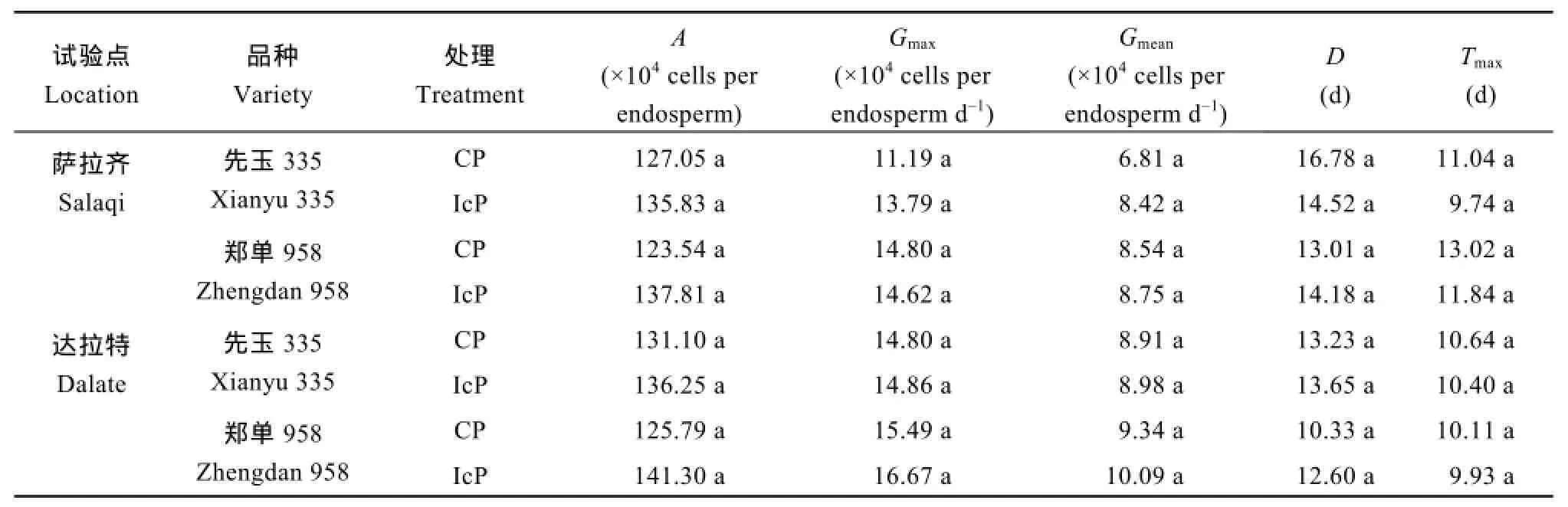
表1 不同控制授粉处理下玉米果穗弱势籽粒胚乳细胞增殖特征参数Table 1 Parameters in endosperm cell division of inferior kernels of maize ear under different pollination treatments
2.3 弱势粒库活性比较
由图 4可见,在授粉后,两试验点玉米弱势粒的可溶性酸性蔗糖转化酶(SAI)活性随生育进程皆呈单峰曲线变化,在24 d前后达到最大值。弱势粒的可溶性酸性蔗糖转化酶(SAI)活性都表现为IcP处理显著高于CP处理,平均差异达12.6%,最大差异达 21.8%,说明可溶性酸性蔗糖转化酶活性的大小与弱势粒败育或灌浆停滞与否有关,弱势粒的早期败育和后期灌浆受限是由于库活性的限制。
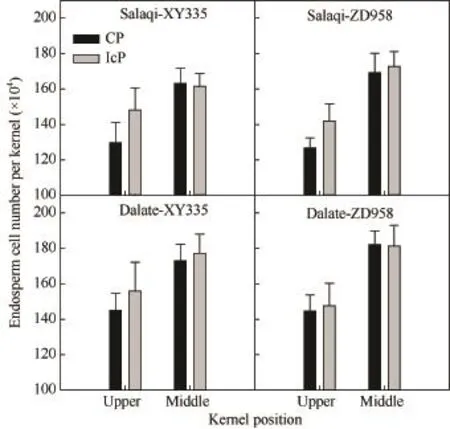
图3 不同控制授粉处理下玉米果穗不同部位籽粒的最大胚乳细胞数比较Fig. 3 Maximum endosperm cell number per kernel at different positions of maize under different pollination treatments
2.4 果穗不同部位籽粒重量比较
不同控制授粉处理下果穗不同部位籽粒的百粒干重以及由Richards方程对弱势粒灌浆动态拟合得出的籽粒灌浆特征参数分别绘于图5和列于表2。
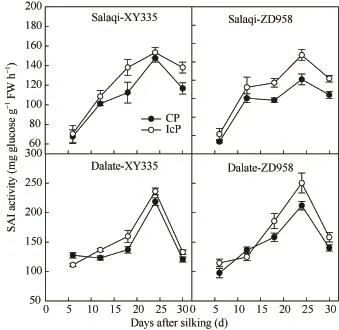
图4 不同控制授粉处理下玉米弱势粒可溶性酸性蔗糖转化酶(SAI)活性变化Fig. 4 Change of soluble acid invertase (SAI) activity ofinferior kernels of maize under different pollination treatments
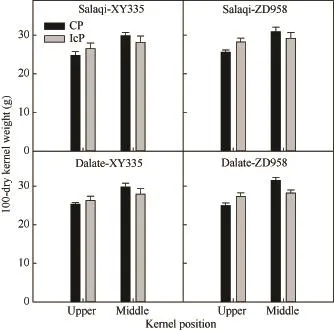
图5 不同控制授粉处理下玉米果穗不同部位籽粒的百粒干重Fig. 5 100-dry kernel weight of kernels at different positions of maize ear under different pollination treatments
不同控制授粉处理下果穗最终籽粒干重均表现为中部籽粒大于上部籽粒,IcP处理差异不显著,但CP处理差异显著,先玉335和郑单958的差异分别达20.6%和25.9%。先玉335顶部弱势粒的最终干重在IcP处理和CP处理间差异不显著,而郑单958表现为IcP处理显著高于CP处理,说明弱势籽粒的灌浆充实存在基因型差异。由表 2可见,弱势粒最终粒重(A)、最大灌浆速率(Gmax)、平均灌浆速率(Gmean)及达到最大灌浆速率的时间(Tmax)均表现为IcP处理显著高于 CP处理,但活跃灌浆期(D)在处理间差异不显著。
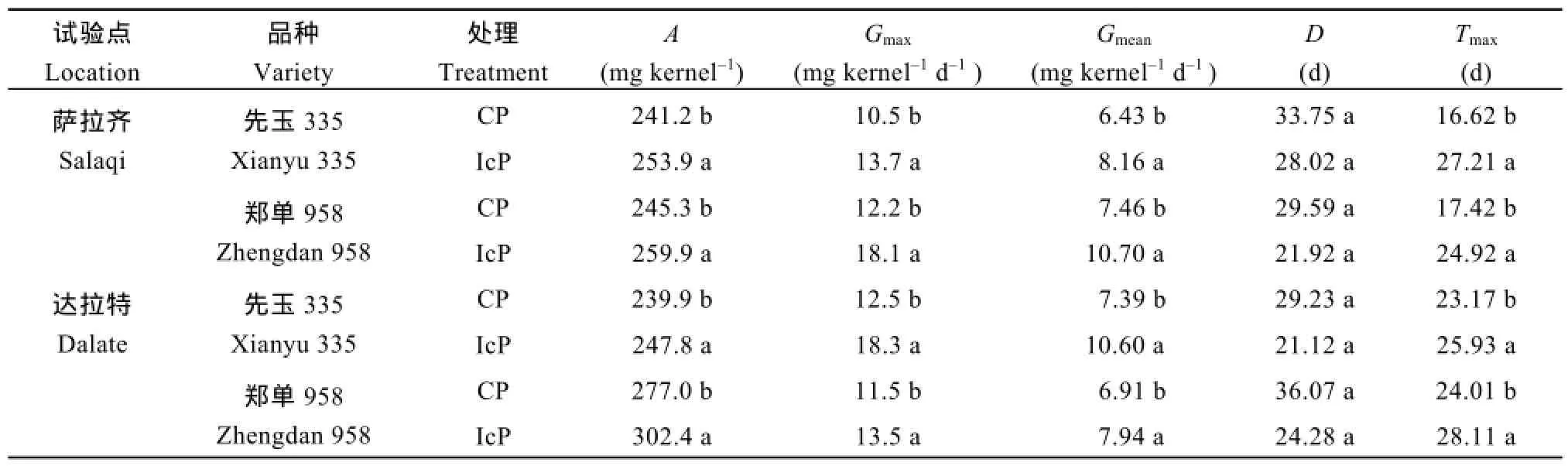
表2 不同控制授粉处理下玉米果穗弱势籽粒灌浆特征参数Table 2 Parameters of kernel-filling of inferior kernels of maize under different pollination treatments
3 讨论
玉米弱势粒败育或灌浆不良是其无法正常积累同化物质的最终结果,Reed等[23]研究表明,玉米散粉后 8~26 d,弱势粒部位穗轴中的总碳水化合物浓度要高于正常籽粒部位穗轴中的浓度,说明受库端的限制,糖分无法从穗轴向籽粒中卸载,使籽粒无法积累同化物质而最终形成无效籽粒。然而,同化物质无法向籽粒当中卸载是受籽粒库容量的限制还是受库活性的限制尚无明确结论。籽粒库容量的建成过程实际上是胚乳细胞分裂和生长的过程,主要取决于胚乳细胞数目的多少[16,24]。徐云姬等[12]在比较了不同部位籽粒的库容特征后认为,与玉米果穗中部籽粒相比上部籽粒重量轻,主要与胚乳细胞增殖速率小因而其胚乳细胞数少有关,但该研究未能回答成功授粉的弱势籽粒后期发生败育或灌浆不良与库容量或库活性的关系。本试验结果表明,无论弱势粒最终发育成功与否,其在籽粒形成阶段的胚乳细胞分裂过程和最终胚乳细胞数皆未受到显著影响,说明库容量不是籽粒败育或灌浆停滞的制约因子。研究表明,籽粒胚乳细胞的分裂与生长素(IAA)和细胞分裂素类物质(Z+ZR)密切相关[12,23],但本研究中不同发育程度的弱势粒库容量没有差异,是否与这些激素含量有关有待进一步研究。
对于玉米而言,籽粒库活性是吸收同化物质并有效转化为淀粉的能力,徐云姬等[13]研究认为,在灌浆期,籽粒中腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、淀粉合酶(StS)和淀粉分支酶(SBE)活性与籽粒灌浆密切相关;而在玉米籽粒形成期,参与蔗糖降解的酶主要是可溶性酸性蔗糖转化酶(SAI),是这一时期库强度的主要衡量指标[9-11]。本研究表明,成功发育弱势粒 SAI活性显著高于败育(或发育不良)弱势粒,且成功发育弱势粒的最大灌浆速率(Gmax)、平均灌浆速率(Gmean)、达到最大灌浆速率的时间(Tmax)和最终粒重皆高于败育(或发育不良)弱势粒,表明SAI活性影响到籽粒的灌浆速率和粒重的增长潜力,也说明弱势粒的早期败育和后期灌浆受限主要受库活性的限制,这与杨建昌等[25]在水稻上的研究结果相同。但目前影响SAI活性的机制尚不清楚。
从前人的研究结果来看,影响 SAI水平的生理原因可能主要有两种。王纪华等[9]和张凤路等[11]认为,籽粒中的内源激素比例影响库活性,当ABA水平偏高,而 GA3水平偏低时,体内可溶性酸性蔗糖转化酶(SAI)活性升高,也就是相对较高的ABA/GA3利于同化物质在籽粒库中的转化卸载,利于籽粒的正常发育。Yang等[26]对水稻弱势粒的研究认为,籽粒中多胺和乙烯的平衡关系影响糖分卸载关键酶的活性。多胺和乙烯有共同的合成前体物质S-腺苷甲硫胺酸(SAM),SAM 既可以在ACC合成酶作用下转化为乙烯的前体物质ACC,又可以在S-腺苷甲硫胺酸脱羧酶(SAMDC)及亚精胺合成酶和精胺合成酶的作用下,经丙胺基转移反应依次形成亚精胺和精胺,当多胺水平较低时,乙烯大量合成并抑制SAI活性,使糖分卸载受阻,籽粒灌浆停止,从而抑制弱势粒的发育[27]。Feng等[28]研究认为,乙烯和多胺的这种平衡关系可能受到同化物质供应量的影响,糖分供应充足时,乙烯合成受到抑制,而促进了多胺的合成。从这一结果推论,不完全授粉条件下果穗顶部弱势粒成功发育并获得较高粒重的原因,可能是不完全授粉条件下顶部弱势粒获得了相对较高的糖分供应条件,进而促进了多胺的合成,而较高的多胺水平则促进了库活性(SAI)的提高并最终促进了弱势粒的灌浆。当然,上述两种影响库活性的作用机制有待于结合弱势粒发育过程中的激素、多胺和糖分水平的分析进一步证实。
4 结论
胚乳细胞分裂过程和最大胚乳细胞数即库容量不是弱势粒败育或灌浆停滞的制约因子,籽粒形成期的库活性是弱势粒败育或灌浆受限的核心限制因子,这可能与不同发育条件或物质供应条件下弱势粒中的激素和多胺的水平有关。
References
[1] Wang Z G,Gao J L,Ma B L. Concurrent improvement in maize yield and nitrogen use efficiency with integrated agronomic management strategies. Agron J,2014,106∶ 1243-1250
[2] 孙国伟,张风路,郭江,赵久然,郭景伦. 对玉米籽粒败育原因的再认识. 玉米科学,2004,12(专刊)∶ 35-37 Sun G W,Zhang F L,Guo J,Zhao J R,Guo J L. Discuss on the cause of maize kernel abortion. J Maize Sci,2004,12(special issue)∶ 35-37 (in Chinese with English abstract)
[3] 申丽霞,王璞,张红芳,易镇邪. 施氮对夏玉米不同部位籽粒灌浆的影响. 作物学报,2005,31∶ 532−534 Shen L X,Wang P,Zhang H F,Yi Z X. Effect of nitrogen supply on grain filling at different ear position in summer maize. Acta Agron Sin,2005,31∶ 532-534 (in Chinese with English abstract)
[4] 李玉玲,台国琴. 玉米籽粒灌浆及种子萌发特性的粒位效应研究. 玉米科学,2005,13(1)∶ 60-63 Li Y L,Tai G Q. Study on the kernel position effects of grain filling and seed germination characteristics in maize. J Maize Sci,2005,13(1)∶ 60−63 (in Chinese with English abstract)
[5] Yang J C,Zhang J H,Wang Z Q,Zhu Q S. Hormones in the grains in relation to sink strength and postanthesis development of spikelets in rice. Plant Growth Regul,2003,41∶ 185−195
[6] Venkateswarlu B,Visperas R M. Source-sink relationships in crop plants. Int Rice Res Paper Ser,1987,125∶ 1−19
[7] Warren W J. Control of crop processes. In∶ Rees A R,Cockshull K E,Hand D W,Hurd R G,eds. Crop Processes in Controlled Environments. London/New York∶ Academic Press,1972. pp 7−30
[8] Ho L C. Metabolism and compartmentation of imported sugars in sink organs in relation to sink strength. Annu Rev Plant Physiol Plant Mol Biol,1988,39∶ 355−378
[9] 王纪华,王树安,赵冬梅,倪军昌. 玉米籽粒发育的调控研究(Ⅲ) 离体条件下的化学调控机理探讨. 作物学报,1996,22∶208−213 Wang J H,Wang S A,Zhao D M,Ni J C. Study on regulation of development of maize kernels∶ III. The approach to chemical regulation under conditions of tissue culture. Acta Agron Sin,1996,22∶ 208-213(in Chinese with English abstract)
[10] 唐祈林,荣廷昭. 玉米秃尖与内源激素的关系. 核农学报,2007,21∶ 366-368 Tang Q L,Rong T Z. The relationship between endogenous hormone and barren ear tip of maize. J Nucl Agric Sci,2007,21∶366−368 (in Chinese with English abstract)
[11] 张凤路,赵久然,王纪华. 离体条件下玉米籽粒发育调控研究.杂粮作物,2001,21(4)∶ 34-35 Zhang F L,Zhao J R,Wang J H. Regulating study on maize kernel development in vitro. Rain Fed Crops,2001,21(4)∶ 34−35 (in Chinese with English abstract)
[12] 徐云姬,顾道健,张博博,张耗,王志琴,杨建昌. 玉米果穗不同部位籽粒激素含量及其与胚乳发育和籽粒灌浆的关系.作物学报,2013,39∶ 1452-1461 Xu Y J,Gu D J,Zhang B B,Zhang H,Wang Z Q,Yang J C. Hormone contents in kernels at different positions on an ear and their relationship with endosperm development and kernel filling in maize. Acta Agron Sin,2013,39∶ 1452-1461 (in Chinese with English abstract)
[13] 徐云姬,顾道建,秦昊,张耗,王志琴,杨建昌. 玉米果穗不同部位籽粒碳水化合物积累与淀粉合成相关酶活性变化. 作物学报,2015,41∶ 297-307 Xu Y J,Gu D J,Qing H,Zhang H,Wang Z Q,Yang J C. Changes in carbohydrate accumulation and activities of enzymes involved in starch synthesis in maize kernels at different positions on an ear during grain filling. Acta Agron Sin,2015,41∶ 297−307 (in Chinese with English abstract)
[14] 冯汉宇,王志敏,孔凡娜,张敏洁,周顺利. 基于控制授粉技术的玉米籽粒生育特性与建成机制研究. 作物学报,2011,37∶1605-1615 Feng H Y,Wang Z M,Kong F N,Zhang M J,Zhou S L. Mechanism of maize kernel setting and characteristics of kernel growth and development under controlled pollination. Acta Agron Sin,2011,37∶ 1605−1615 (in Chinese with English abstract)
[15] 张祖建,朱庆森,王志琴. 水稻胚乳细胞计数方法研究. 江苏农学院学报,1996,17(2)∶ 7−11 Zhang Z J,Zhu Q S,Wang Z Q. A method of counting endosperm cells in rice grain. J Jiangsu Agric Coll,1996,17(2)∶ 7−11 (in Chinese with English abstract)
[16] 王晓燕,董树亭,高荣岐,张海艳. 不同类型玉米胚乳细胞增殖动态及其与粒重的关系. 华北农学报,2006,21(2)∶ 23-26 Wang X Y,Dong S T,Gao R Q,Zhang H Y. Endosperm cell proliferating and its relation to grain weight in different types of maize. Acta Agric Boreali Sin,2006,21(2)∶ 23−26 (in Chinese with English abstract)
[17] 周竹青,朱旭彤,王维金. 三种粒型小麦品种胚乳细胞增殖动态研究. 生物数学学报,2004,19∶ 123-126 Zhou Z Q,Zhu X T,Wang W J. Study on the dynamic of endosperm proliferation cell in wheat cultivars (lines) with three grain types. J Biomath,2004,19∶ 123−126 (in Chinese with English abstract)
[18] Singh B K,Jenner C F. A modified method for the determination of cell number in wheat endosperm. Plant Sci Lett,1982,26∶273−278
[19] 张海艳,董树亭,高荣岐. 不同类型玉米籽粒灌浆特性分析.玉米科学,2007,15(3)∶ 67-70 Zhang H Y,Dong S T,Gao R Q. Analysis on grain filling characteristics of different types of maize. J Maize Sci,2007,15(3)∶67−70 (in Chinese with English abstract)
[20] 杨升辉,杨恒山,唐汝友,王素阁,徐长帅,陈勇. 不同氮肥运筹下春玉米籽粒灌浆特性的分析. 玉米科学,2014,22(1)∶91−95 Yang S H,Yang H S,Tang R Y,Wang S G,Xu C S,Chen Y. Analysis on kernel filling characteristics for spring maize in different nitrogen fertilizer strategies. J Maize Sci,2014,22(1)∶91−95 (in Chinese with English abstract)
[21] 谷岩,王振明,何文安,烜梁赫,吴春胜. 不同类型玉米品种籽粒胚乳细胞增殖与籽粒建成的关系. 西北农林科技大学学报,2010,38(1)∶ 90−96 Gu Y,Wang Z M,He W A,Liang X H,Wu C S. Relationship between endosperm cell proliferation and seed formation in maize of different types. J Northwest A&F Univ,2010,38(1)∶90−96 (in Chinese with English abstract)
[22] 高俊凤. 植物生理学实验指导. 北京∶ 高等教育出版社,2006. pp 105-108 Gao J F. Plant Physiology Experiment Instruction. Beijing∶Higher Education Press,2006. pp 105−108 (in Chinese)
[23] Reed A J,Singletary G W. Roles of carbohydrate supply and phytohormones in maize kernel abortion. Plant Physiol,1989,91∶986−992
[24] 秦大鹏,刘鹏,王空军,董树亭,张吉旺,刘月娥. 高淀粉玉米籽粒胚乳细胞增殖与淀粉积累的关系. 山东农业科学,2008,8∶ 35−39 Qin D P,Liu P,Wang K J,Dong S T,Zhang J W,Liu Y E. Relationship between endosperm cell proliferation and starch accumulation in high-starch maize grain. Shandong Agric Sci,2008,8∶35−39 (in Chinese with English abstract)
[25] 杨建昌,刘立军,王志琴,郎有忠,朱庆森. 稻穗颖花开花时间对胚乳发育的影响及其生理机制. 中国农业科学,1999,32(3)∶ 44-51 Yang J C,Liu L J,Wang Z Q,Lang Y Z,Zhu Q S. Effects of flowering time of spikelets on endosperm development in rice and its physiological mechanism. Sci Agric Sin,1999,32(3)∶ 44-51
[26] Yang J C,Cao Y Y,Zhang H,Liu L J,Zhang J H. Involvement of polyamines in the post-anthesis development of inferior and superior spikelets in rice. Planta,2008,228∶ 137−149
[27] 张风路,王志敏,赵明,王树安,赵久然,郭景伦. 多胺与玉米籽粒败育关系研究. 作物学报,1999,25∶ 565−568 Zhang F L,Wang Z M,Zhao M,Wang S A,Zhao J R,Guo J L. Studies on the relationship between polyamines and kernel abortion in maize. Acta Agron Sin,1999,25∶ 565−568 (in Chinese with English abstract)
[28] Feng H Y,Wang Z M,Kong F N,Zhang M J,Zhou S L. Roles of carbohydrate supply and ethylene,polyamines in maize kernel set. J Integr Plant Biol,2011,53∶ 388−398
Relationship of Inferior Kernel Setting with Sink Strength under Controlling Pollination
LIANG Hong-Wei,GAO Ju-Lin,WANG Zhi-Gang*,YU Xiao-Fang,SUN Ji-Ying,HU Shu-Ping,WANG Zhen,YU Shao-Bo,and LI Ya-Jian
College of Agronomy,Inner Mongolia Agricultural University,Hohhot 010019,China
Abstract:Clarifying the relationship of sink capacity and/or sink activity with kernel abortion and/or filling stagnation of inferior kernels,is of great importance for closing the yield gap by improving the grain filling of inferior kernels of maize. In this present study,two typical commercial maize hybrids Zhengdan 958 and Xianyu 335 were planted at two locations. At silking stage,two pollination treatments,incomplete pollination (IcP) and complete pollination (CP),were imposed to each cultivar at each location. The objective of incomplete pollination,in which the pollinated filaments within the basal region of the ear was decreased by handing-pollinating,was to stimulate the inferior kernel set in the apical region of the ear,where the inferior kernels should be aborted or filling-stagnated in complete pollination. The sink capacity,sink activity and grain filling parameters were compared within IcP and CP treatments. The endosperm cell division process and the maximum endosperm cell number of inferior kernels showed no difference between IcP and CP treatments,while the soluble acid invertase activity of IcP was significantly higher than that of CP,by 12.6% on an average and 21.8% at the maximum. Furthermore,the actual 100-kernel weight,the stimulated final kernel weight,the maximum kernel-filling rate and the mean kernel-filling rate of IcP treatment were also higher than those of CP. The results suggested that the abortion and/or filling stagnation of inferior kernels of maize is not limited by its sink capacity;sink activity during blister stage is the primary limited factor of the abortion and/or filling stagnation of inferior kernels.
Keywords:Maize;Inferior kernel;Controlling pollination;Sink capacity;Sink activity
DOI:10.3724/SP.J.1006.2016.00917
*通讯作者(
Corresponding author)∶ 王志刚,E-mail∶ imauwzg@163.com,Tel∶ 13734813561
收稿日期Received()∶ 2015-12-01;Accepted(接受日期)∶ 2016-03-14;Published online(网络出版日期)∶ 2016-03-21.
