喙畸形研究进展
2016-07-12孙研研陈继兰
白 皓,孙研研,陈继兰
(中国农业科学院北京畜牧兽医研究所,农业部畜禽遗传资源与种质创新重点实验室,北京 100193)
喙畸形研究进展
白皓,孙研研,陈继兰*
(中国农业科学院北京畜牧兽医研究所,农业部畜禽遗传资源与种质创新重点实验室,北京 100193)
摘要:喙是鸟类上下颌包被的硬角质鞘,起到哺乳动物唇和齿的作用。喙畸形是近年来在鸟类发现并逐渐受到重视的一类骨骼疾病。喙畸形在家禽尤其是我国一些地方鸡品种中常见发生,主要表现为上下喙交错,咬合不全,呈交叉状态,严重影响个体饮水和采食,造成生长缓慢甚至死亡,给家禽生产造成巨大的经济损失,同时也威胁动物福利。目前,国内外关于鸡喙的起源、形态结构、生长发育等方面的研究相对较少,鸡喙畸形的形成机制尚不明确,而在鸟类,尤其是达尔文雀喙畸形的研究取得了较大的进展。本文综述了鸟类(包括鸡)喙的起源、形态结构、生长发育、影响因素以及其他物种唇部形态变化等相关研究,以期为人们开展鸡喙形态发育及喙畸形的分子机制研究提供借鉴。
关键词:鸡;喙畸形;鸟类;骨骼疾病
鸟类的喙由上下颌构成其骨质部分,表面被覆角质层,起到唇和齿的作用,主要用于采食、饮水、梳理羽毛以及攻击有害目标等。同时,喙的大小、色泽等也是禽类进行自由交配时的择偶条件之一[1]。在家禽生产中,喙的健康关系到个体采食和饮水质量,是家禽正常生长发育的重要保证。近年来,本研究团队在北京油鸡的世代繁育过程中发现了一定比例的喙畸形个体,主要表现为上下喙错位、咬合不全、呈交叉状态(图1)。据调查,喙畸形在文昌鸡、胡须鸡和清远麻鸡等地方鸡品种中也有1%~3%的发生率[2]。本研究团队在进行北京油鸡世代繁育过程中观察并统计发现,喙畸形鸡由于不能正常采食和饮水,生长发育受到不同程度影响,死亡率比一般鸡高出数倍,存活下来的鸡成年后能正常繁殖的比例也比正常鸡低50%以上,制约了地方鸡的产业化发展。近年来,喙畸形在多个物种中被相继报道,国际国内专家学者研究了野生鸟类和家禽喙的发育及畸形发生原因,并取得了一定的进展。本文即对近年来喙畸形的研究进展进行综述。
1喙的形成
鸟类喙的生长发育始于胚胎时期[3]。在鸟类头部发育过程中,分化和迁移的神经嵴细胞、感觉结构和颅面始基(包括鼻额突、下颌弓和舌骨弓)之间具有一定的解剖关系。喙起源于前移到鼻额突、下颌弓和舌骨弓中的神经嵴细胞[4],神经嵴对喙形态结构的形成起到关键的作用。上喙由额鼻原基和一对上颌骨原基构成,而下喙则由下颌骨原基构成。上下颌原基主要由间质和上皮两种组织构成。间质由来自神经嵴和中胚层间的叶细胞构成,上皮则由紧密连接的上皮细胞构成,它是机体与外界环境的屏障。上皮细胞和间叶细胞可同时促进喙的形成[5]。其中,间叶细胞对喙的形成起主导作用[6-7]。而上皮细胞通过诸如成纤维生长因子8(Fibroblast growth factor 8,FGF8)和音猬因子(Sonic hedgehog,SHH)之类的调节分泌因子混合物可直接调控喙的发育及相关衍生物的生成[8]。因此,上皮细胞也是喙及面部骨骼形成的必要因素。
2喙的结构、形态
喙由多种面部突起组成[5,9],额鼻骨、外侧鼻突起和上颌突起构成了上喙,下颌突起形成下喙。在不同的鸟类中,这些突起相互协调以适当的比例组成特异的喙的大小和形状。鸟类喙的结构、形态大致可以分为以下几种类型:食昆虫的鸟—尖而细长,方便取出狭窄细缝中的昆虫;食坚果的鸟—粗短成圆锥形,方便磕破坚硬的果子;食肉的鸟—粗壮、尖端锐利、成钩型弯曲,方便捕杀及撕开猎物;食鱼的鸟—细长而锐利,方便叉住游动的鱼;食浮游生物的鸟—扁平宽阔、有滤水的栉缘,方便在水中过滤浮游生物。有些个别的鸟喙也有其独特的形态。鸡喙较粗短,呈圆锥形,多与胫色一致,有硬的角质被覆喙部,分为上下两个部分,形成喙的骨质基础[10]。上喙由上颌骨、鼻骨和切齿骨组成,下喙由下颌骨组成,下颌骨与颞骨之间有特殊的方骨,开口时可将口腔拉大,便于摄食和饮水[1,11]。鸡喙的正常特征是上下颌完全闭合,向前方突出,前端呈箭头状,舌位于口腔内,在口腔顶部的两个黏膜褶壁中间有内鼻孔,口腔后部为咽部。
3禽类喙畸形的发现
20世纪,一些野生鸟类中发现的喙畸形现象引起了鸟类科学家和鸟类保护工作者的广泛重视。D.A.Rintoul[12]在赖利野生动物保护群中发现褐头牛鸟出现喙畸形个体。C.M.Handel 等[13]在阿拉斯加野生鸟黑头山雀中也发现喙畸形流行病,该病是北美野生鸟类保护区出现的新病症。C.Van Hemert 和C.M.Handel[14]在西北乌鸦中发现有多只喙畸形个体,并将其定义为一种多物种流行病。之后研究人员在其他沿海禽类中也发现喙畸形个体,认为喙畸形是海洋生物圈的怪现象。同样,喙畸形在非洲黑腹砸籽雀、泰和乌鸡、日本鹌鹑、雀形目(地雀、仙人掌雀、达尔文雀群等)以及各品种鹦鹉等物种中也相继被报道。
4喙形态发育的影响因素
导致喙形态发生变化的原因有很多,包括环境、营养、疾病等非遗传性因素和遗传因素。其中,环境和遗传是两个主要的影响因素。
4.1非遗传因素
1962年,D.E.Pomeroy[15]统计发现,野生鸟类喙畸形的发生率为0.5%,其主要原因可能是钙的营养水平不足或断喙不当。2010年,C.M.Handel等[13]统计了过去十年的数据,发现由于生态系统发生重大变化,阿拉斯加地区黑顶山雀喙畸形发生率已达到6.5%。B.P.Tangredi[16]研究发现,维生素和钙离子代谢相关营养素的缺失可导致喙畸形发生。Y.Song等[17]在对颅面骨骼维甲酸合成控制的研究中发现,外源性维生素—柠檬醛可诱导阻断类视黄醇的合成,导致鼻骨右侧细胞死亡,从而引起额鼻部形态的缺陷。同时,受伤和感染也能影响喙的发育。细菌、病毒、真菌、寄生虫感染都会引起喙的畸变[16,18];肝病[19]、肿瘤[20]等疾病也会引起喙畸形。D.Vesela等[21]研究发现,赭曲霉毒素的毒性作用对鸡的精神发育、颅脑畸形、小眼、断喙等都有影响。T.Chan等[22]研究表明,如果在孵化阶段给予鸡胚低氧环境可导致鸡在生长后期喙长明显变短,尤其是在胚胎发育前期1~6 d,对后期喙骨形成的影响最大。
4.2遗传因素
非遗传因素对于喙形态的影响是显而易见的,但是其影响一般都是后天造成的,即动物在其生长发育过程中受到的外界影响所致,可以直接或间接导致鸡喙畸形的发生。而许多情况下,喙畸形在个体发育早期(甚至胚胎期)就已经产生,是先天性的,甚至是由上一代传递给下一代的,这就是遗传因素。本研究团队通过多年的观察发现,每个世代正常公母鸡交配后代中仍有畸形个体出现,通过表型观察很难将喙畸形性状从群体中剔除。有系谱记录的喙畸形个体中,向上追溯4代,其中部分鸡可以追溯到共同祖先,连续两代都发生喙畸形的占所有喙畸形鸡的比例为62.5%,据此判断其有遗传趋势[23]。同时,本研究团队利用喙畸形公鸡与喙畸形母鸡进行交配,发现后代喙畸形的比例达到8%~10%,显著高于喙畸形公鸡与正常母鸡交配畸形后代的比例(0%~3%),也比正常群体中喙畸形比例提高5倍左右,因此可初步推测,鸡喙畸形的发生具有一定的遗传基础,相关调控基因的变异导致了喙畸形的发生。早在1998年,M.Tsudzuki等[24]报道,常染色体隐性基因(Hmm)纯合的日本雀个体出现上下喙畸形,胚胎形态畸形,并趾,骨架不正常等现象。在鸡喙研究方面,M.E.MacDonald等[25]发现,FGF8的下调表达可导致鸡喙额鼻部上皮细胞发生凋亡,从而破坏上皮细胞间质信号转导通路,最终导致鸡上喙部出现截断现象。M.R.Plant等[26]将鸡喙的形态异常分为不同的类型,通过试验进一步验证了维甲酸通路对鸡喙形态的发育起着重要的作用,即维甲酸通路可通过改变方骨的位置和转向来调节鸡喙的形态。对于骨形态发生蛋白4(Bone morphogenetic protein 4,BMP4)基因的研究越来越多,其在喙形态多样性方面起着重要作用。P.Wu等[3]在探索喙形成机制的研究中发现,BMP4基因对喙形态具有一定调节作用。该基因在喙的整个发育过程中表达的水平、时间或部位不同都会导致喙形态不一,即喙的长度、宽度、深度都会发生相应的变化。E.H.Groeneveld等[27]在骨组织形成蛋白对于人类骨骼再生的研究中表明,BMP4基因能够调控早期胚胎发育、诱导成骨、促进骨胶原生成、骨痂的形成和再塑,进一步说明BMP4基因在遗传方面可能影响喙骨的形态变化。在胚胎15HH时期,BMPs基因在上颌区域(在第一鳃弓和眼之间)的间质细胞中表达[28-29]。对15HH阶段胚胎进行短暂的提高维甲酸水平和封闭BMPs信号处理,胚胎上颌突起前端转变为额鼻骨物质。在随后的发育阶段,BMPs和维甲酸对于突起的形成具有不同的作用[30]。上述研究是基于喙基本形态变化,并未涉及交叉喙这种严重畸形情况的发生,目前有关鸡交叉喙相关的文献报道较少,相关研究才刚刚起步。K.Jaszczak等[31]在研究中发现,同源盒A1(Homeobox protein Hox-A1,HOXA1)和同源盒D3(Homeobox protein Hox-D3,HOXD3)基因的异常表达可导致雏鸡喙畸形的发生率高达16%。H.Bai等[32]通过数字基因表达谱技术对鸡喙畸形研究发现,BMP4等11个基因可作为喙畸形研究的重要候选基因。其中,白皓等[33]研究初步认为,α-半乳糖苷酶(Galactosidase,Alpha,GLA)基因多态性与北京油鸡喙畸形性状可能存在一定的关联性,其SNPs可能是喙畸形性状的潜在遗传标记。朱静[2]研究发现,类似角蛋白(LOC426217)基因在畸形喙中表达量显著高于正常喙,且具有喙组织特异性(即在鸡喙组织中表达量极显著高于其它组织),该基因可作为喙畸形性状研究的重要候选基因。罗成龙等利用鸡60K的SNP芯片对30只胡须鸡(喙畸形和正常鸡各15只)进行SNP分型,发现WNT9B附近的SNP与喙畸形性状显著相关并具有最大的OR值,且该基因与哺乳动物唇腭裂密切相关,可能是引起鸡喙畸形的重要候选基因[34]。这些遗传方面的研究值得思考和借鉴,为揭示鸡喙畸形的分子遗传机制提供了一定的参考价值和理论依据。
遗传与非遗传因素是相辅相成的,许多异常发育可能是由于两者的相互作用所造成的。比如说亲代由于非遗传因素导致了喙畸形,这种由外因导致的疾病在产生下一代的过程中可能会使下一代有喙畸形的个体存在,即表观遗传。尽管遗传选择能够主导亲代基因传递给后代,但越来越多的证据表明外界因素对基因表达具有重要的修饰作用。因此,对于喙畸形的研究,也不妨从表观遗传的角度去尝试一下,也许会有意想不到的收获。
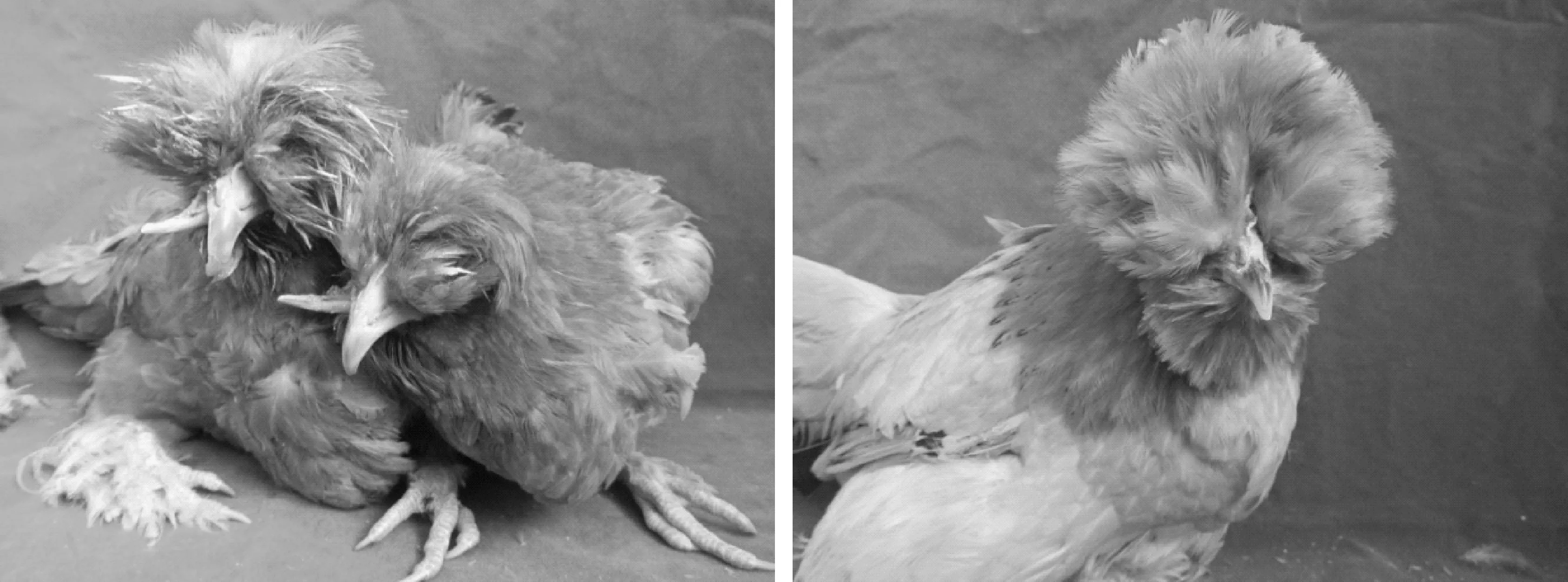
图1 北京油鸡喙畸形鸡(左)与喙正常鸡(右)Fig.1 The deformed(left) and normal(right) beaks of Beijing-You chickens
5其他物种唇部形态异常的相关研究
目前,关于唇部形态异常的研究主要集中在人和小鼠上。唇腭裂、歪嘴哭综合征、无牙症、家畜歪嘴风等症状在临床上常常出现,给人和动物带来了许多危害。如今,家养动物对于人们的生活越来越重要,给人类带来了许多经济效益。然而,这些疾病的出现,却给人类造成了严重的经济损失。
I.Satokata等[35]以小鼠为模型在进行同源异型盒基因1(Muscle Segment Homeobox 1,MSX1)基因突变研究中发现,该基因与包括唇腭裂和无牙症在内的特定的颅面畸形有一定的关联。A.C.Marcano等[36]在T盒转录因子(T-box transcription factor,TBX22)的研究中发现,该基因在腭部和舌体中都有表达,其突变可能导致人或小鼠唇腭裂的发生。O.Britanova等[37]在对新型转录因子SATB2(Special AT-rich sequence-binding protein 2)与基质结合区DNA分子特异性相互作用以及小鼠发育依赖性细胞在神经中枢表达的研究中发现,SATB2基因与非综合症型唇腭裂(NSCLP)有关,敲除小鼠的SATB2基因可能会产生唇腭裂,该基因的多态性与唇腭裂呈显著相关。近年来,人们对SUMO家族中的小类泛素修饰因子-1(SUMO-1)的关注越来越多。SUMO-1广泛参与了唇腭裂易感基因转录后蛋白质的SUMO化修饰。廖礼姝等[38]就SUMO化修饰及其在非综合征性唇腭裂发病中的意义作出了详细的阐述。T.H.Beaty等[39]关于人类唇腭裂候选基因的研究表明SUMO-1与DNA结合蛋白结合,可以作为转录因子来调控SATB2基因。近期,通过全基因组关联分析和验证研究,在人类基因组中发现一个可能会导致非综合征型唇腭裂发生的位点,该位点在人的16号染色体上,编码为rs8049367,位于CREBBP和ADCY9基因之间[40]。
其他物种,尤其是人和小鼠唇部形态异常的研究已经处于一种发展相对较快、趋于成熟和完善的阶段,而鸡喙形态变化的研究相对较少,尤其是遗传方面的研究。因此,在鸡上的研究可以借鉴哺乳动物相关研究思路和方法,找出影响鸡喙形态变化的主要遗传因素,为喙形态发育的进一步研究打下坚实的基础。
6小结
鸡喙畸形的发生影响养殖效益及动物福利,早期的研究已揭示该性状的可遗传性,但是具体形成机制尚不明确。利用先进的分子生物学技术:如重测序、高通量分型芯片、转录组测序等,采取数量遗传和分子遗传相结合的方法,挖掘导致喙畸形的重要基因或分子标记,一方面有利于在生产实践中通过分子检测手段快速准确剔除不利基因或基因型,优化群体遗传背景,减少因畸形个体出现造成的经济损失;另一方面可为鸟类喙畸形和哺乳类颅面畸形研究提供相关参考。
参考文献(References):
[1]罗克.家禽解剖学与组织学[M].福州:福建科学技术出版社,1983.
LUO K.Poultry anatomy and histology[M].Fuzhou:Fujian Science & Technology Publishing Housing,1983.(in Chinese)
[2]朱静.鸡喙组织致畸基因的筛选与验证[D].北京:中国农业科学院,2012.
ZHU J.Digital screening and identification of teratogenic genes underlying beak deformity in chickens[D].Beijing:Chinese Academy of Agricultural Sciences,2012.(in Chinese)
[3]WU P,JIANG T X,SUKSAWEANG S,et al.Molecular shaping of the beak[J].Science,2004,305(5689):1465-1466.
[4]BOUVIER M,HYLANDER W L.The effect of dietary consistency on morphology of the mandibular condylar cartilage in young macaques(Macacamulatta)[J].ProgClinBiolRes,1981,101:569-579.
[5]RICHMAN J M,LEE S H.About face:signals and genes controlling jaw patterning and identity in vertebrates[J].Bioessays,2003,25(6):554-568.
[6]SCHNEIDER R A,HELMS J A.The cellular and molecular origins of beak morphology[J].Science,2003,299(5606):565-568.
[7]MCQUALTER J L,YUEN K,WILLIAMS B,et al.Evidence of an epithelial stem/progenitor cell hierarchy in the adult mouse lung[J].ProcNatlAcadSciUSA,2010,107(4):1414-1419.
[8]ABZHANOV A,PROTAS M,GRANT B R,et al.Bmp4 and morphological variation of beaks in Darwin’s finches[J].Science,2004,305(5689):1462-1465.
[9]HELMS J A,SCHNEIDER R A.Cranial skeletal biology[J].Nature,2003,423(6937):326-331.
[10]BRAGULLA H H,HOMBERGER D G.Structure and functions of keratin proteins in simple,stratified,keratinized and cornified epithelia[J].JAnat,2009,214(4):516-559.
[11]MALLARINO R,GRANT P R,GRANT B R,et al.Two developmental modules establish 3D beak-shape variation in Darwin’s finches[J].ProcNatlAcadSciUSA,2011,108(10):4057-4062.
[12]RINTOUL D A.Beak deformity in a brown-headed cowbird,with notes on causes of beak deformities in birds[J].KansasOrnitholSocBull,2005,56:29-32.
[13]HANDEL C M,PAJOT L M,MATSUOKA S M,et al.Epizootic of beak deformities among wild birds in Alaska:An emerging disease in North America?[J].Auk,2010,127(4):882-898.
[14]VAN HEMERT C,HANDEL C M.Beak deformities in Northwestern crows:Evidence of a multispecies epizootic[J].Auk,2010,127(4):746-751.
[15]POMEROY D E.Birds with abnormal bills[J].BritishBirds,1962,55(2):49-72.
[16]TANGREDI B P.Environmental factors associated with nutritional secondary hyperparathyroidism in wild birds[J].AvianPoultBiolRev,2007,18(2):47-56.
[17]SONG Y,HUI J N,FU K K,et al.Control of retinoic acid synthesis and FGF expression in the nasal pit is required to pattern the craniofacial skeleton[J].DevBiol,2004,276(2):313-329.
[18]STEVENS V I,BLAIR R,SALMON R E,et al.Effect of varying levels of dietary vitamin D3 on turkey hen egg production,fertility and hatchability,embryo mortality and incidence of embryo beak malformations[J].PoultSci,1984,63(4):760-764.
[19]LUMEIJ J T,DE KONING J,BOSMA R B,et al.Treponemal infections in hares in The Netherlands[J].JClinMicrobiol,1994,32(2):543-546.
[20]OWEN H C,DONELEY R J,SCHMIDT R E,et al.Keratoacanthoma causing beak deformity in a budgerigar(Melopsittacusundulatus)[J].AvianPathol,2007,36(6):499-502.
[22]CHAN T,BURGGREN W.Hypoxic incubation creates differential morphological effects during specific developmental critical windows in the embryo of the chicken(Gallusgallus)[J].RespirPhysiolNeurobiol,2005,145(2-3):251-263.
[23]白皓.北京油鸡喙畸形性状候选基因的转录水平研究及SNPs分析[D].扬州:扬州大学,2013.
BAI H.The mRNA expression and SNPs analysis of candidate genes associated with beak deformity trait in Beijing-You chickens[D].Yangzhou:Yangzhou University,2013.(in Chinese)
[24]TSUDZUKI M,NAKANE Y,WADA A.Hereditary multiple malformation in Japanese quail:a possible powerful animal model for morphogenetic studies[J].JHered,1998,89(1):24-31.
[25]MACDONALD M E,ABBOTT U K,RICHMAN J M.Upper beak truncation in chicken embryos with the cleft primary palate mutation is due to an epithelial defect in the frontonasal mass[J].DevDyn,2004,230(2):335-349.
[26]PLANT M R,MACDONALD M E,GRAD L I,et al.Locally released retinoic acid repatterns the first branchial arch cartilagesinvivo[J].DevBiol,2000,222(1):12-26.
[27]GROENEVELD E H,BURGER E H.Bone morphogenetic proteins in human bone regeneration[J].EurJEndocrinol,2000,142(1):9-21.
[28]WALL N A,HOGAN B L.Expression of bone morphogenetic protein-4(BMP-4),bone morphogenetic protein-7(BMP-7),fibroblast growth factor-8(FGF-8) and sonic hedgehog(SHH) during branchial arch development in the chick[J].MechDev,1995,53(3):383-392.
[29]NIEDERREITHER K,SUBBARAYAN V,DOLLÉ P,et al.Embryonic retinoic acid synthesis is essential for early mouse post-implantation development[J].NatGenet,1999,21(4):444-448.
[30]LEE S,FU K K,HUI J N,et al.Noggin and retinoic acid transform the identity of avian facial prominences[J].Nature,2001,414(6866):909-912.
[31]JASZCZAK K,MALEWSKI T,PARADA R,et al.Expression of Hoxa1 and Hoxd3 genes in chicken embryos with exencephaly[J].JAnimFeedSci,2006,15(3):463-469.
[32]BAI H,ZHU J,SUN Y,et al.Identification of genes related to beak deformity of chickens using digital gene expression profiling[J].PLoSOne,2014,9(9):e107050.
[33]白皓,陈继兰,刘冉冉,等.北京油鸡喙畸形性状候选基因GLA多态性的研究[J].中国畜牧杂志,2013,49(15):1-6.
BAI H,CHEN J L,LIU R R,et al.Study on polymorphisms ofGLAas a candidate gene for the beak deformity trait in Beijing-You chickens[J].ChineseJournalofAnimalScience,2013,49(15):1-6.(in Chinese)
[34]罗成龙,马杰,瞿浩,等.鸡交叉喙的全基因组关联研究[C].第十三次全国畜禽遗传标记研讨会论文集,动物生物技术通讯(英文版),2012,12(1):26.
LUO C L,MA J,QU H,et al.Genome-wide association study on crossed beak in chickens[C].Proceeding of the 13th National Symposium on Animal Genetic Markers,Animal Biotechnology Bulletin,2012,12(1):26.(in Chinese)
[35]SATOKATA I,MAAS R.Msx1 deficient mice exhibit cleft palate and abnormalities of craniofacial and tooth development[J].NatGenet,1994,6(4):348-356.
[36]MARCANO A C,DOUDNEY K,BRAYBROOK C,et al.TBX22 mutations are a frequent cause of cleft palate[J].JMedGenet,2004,41(1):68-74.
[37]BRITANOVA O,AKOPOV S,LUKYANOV S,et al.Novel transcription factor Satb2 interacts with matrix attachment region DNA elements in a tissue-specific manner and demonstrates cell-type-dependent expression in the developing mouse CNS[J].EurJNeurosci,2005,21(3):658-668.
[38]廖礼姝,蒙田,郑谦,等.小遍在蛋白质样修饰物化修饰及其在非综合征性唇腭裂发病中的意义[J].国际口腔医学杂志,2009,36(1):94-97.
LIAO L S,MENG T,ZHENG Q,et al.Role of small ubiquitin like modifier in etiology of nonsyndromic cleft lip and palate[J].InternationalJournalofStomatology,2009,36(1):94-97.(in Chinese)
[39]BEATY T H,HETMANSKI J B,FALLIN M D,et al.Analysis of candidate genes on chromosome 2 in oral cleft case-parent trios from three populations[J].HumGenet,2006,120(4):501-518.
[40]SUN Y,HUANG Y,YIN A,et al.Genome-wide association study identifies a new susceptibility locus for cleft lip with or without a cleft palate[J].NatCommun,2015,6:6414.
(编辑郭云雁)
The Research Progress of Beak Deformity
BAI Hao,SUN Yan-yan,CHEN Ji-lan*
(TheKeyLaboratoryofFarmAnimalGeneticResourcesandGermplasmInnovationofMinistryofAgriculture,InstituteofAnimalScience,ChineseAcademyofAgriculturalSciences,Beijing100193,China)
Abstract:Beak is an external anatomical structure of birds.Its functions are the same as lips and teeth of mammals.Beak deformity,a new skeletal disease,was first reported in birds and gained more and more attention recently.Beak deformity(normally a crossed beak) has also been found in some indigenous chicken breeds in China.The chicken with a deformed beak has problems in eating and drinking,which could cause slow growth and early death.This causes economics loss and is harmful to the animal welfare.Until now,the origin,morphosis,growth and development of beak has less been studied.The mechanism of beak deformity in chickens is not clear yet.The study of beak deformity in Darwin’s finches has a greater progress.This article summarized these related studies in birds,aiming to provide guidance for the study of beak development and molecular mechanism of beak deformity in chickens.
Key words:chicken;beak deformity;birds;skeletal disease
doi:10.11843/j.issn.0366-6964.2016.03.001
收稿日期:2015-04-27
基金项目:国家自然科学基金青年基金项目(31501949);国家863高技术研究发展计划(2011AA100305);中央级公益性科研院所基本科研业务费专项资金(2014ywf-yb-3)
作者简介:白皓(1987-),男,江苏扬州人,博士生,主要从事家禽遗传育种研究,E-mail:bhowen1027@163.com *通信作者:陈继兰,博士,研究员,博士生导师,主要从事家禽遗传育种研究,E-mail:chen.jilan@163.com
中图分类号:S852.21
文献标志码:A
文章编号:0366-6964(2016)03-0417-06
