辽宁省老鹳草属植物种质资源遗传多样性的ISSR分析
2016-07-11孙鹏谷静逸尹海波张建逵贾晓晴邵飞
孙鹏,谷静逸,尹海波,张建逵,贾晓晴,邵飞
(1.沈阳市第七人民医院,辽宁 沈阳 110003;2.辽宁中医药大学药学院,辽宁 大连 116600)
老鹳草,始见于明代兰茂的《滇南本草》[1],是我国传统中药,来源于牻牛儿苗科植物牻牛儿苗(Erodiumstephanianum Willd)、老鹳草(Geranium wilfordii Maxim)或野老鹳草(G carolinianumL)的干燥地上部分,收载于我国2015年版《药典》[2]。我国有牻牛儿苗科植物4属,约67种,其中老鹳草属植物55种[3],其中分布于辽宁省的有7种,分别为老鹳草(Geranium wilfordii Maxim)、鼠掌老鹳草(Geranium sibiricum L)、块根老鹳草(Geranium dahuricum DC)、突节老鹳草 (Geranium Krameri Franch.et Sav Enum)、朝鲜老鹳草(Geranium franchetii R.Kunth)、长白老鹳草(Geranium baishanense Y.L.Chang)和毛蕊老鹳草(Geranium eriostemon Fish.ex DC)[3-5]。本研究利用ISSR分子标记技术,揭示了辽宁省老鹳草属植物资源的亲缘关系及群体间的遗传多样性。
1 材料
供试材料均于2014和2015年7-9月采自辽宁省东部山区的21个居群,均经大连自然博物馆张淑梅教授和辽宁中医药大学药用植物教研室尹海波教授鉴定,具体信息见表1。

表1 样品基本信息
2 方法
2.1 DNA提取
将存储在-80℃冰箱内的实验材料取出,迅速置于研钵中充分研磨。按照TIANGEN植物基因组DNA提取试剂盒的说明进行DNA的提取,用1.5%的琼脂糖凝胶电泳检测,各个电泳道均出现一条清晰的DNA条带,即所提取的DNA均完整。
2.2 ISSR引物的选择
本实验所用ISSR引物由上海生工生物工程股份有限公司合成,源自University of British Columbia(UBC大学)公布的100条ISSR引物。根据相关论文[6],结合本实验材料,从中筛选出扩增产物条带清晰、多态性较好的引物用于不同种老鹳草亲缘关系的分析。
2.3 老鹳草属植物ISSR-PCR最佳反应体系的建立
PCR反应体系的优化是ISSR技术获得较好研究结果的先决条件[7],本实验通过对退火温度、引物浓度和DNA模板量进行优化,建立了PCR的最佳反应体系。扩增产物经1.5%琼脂糖凝胶电泳,溴化乙锭(上海Sangon)染色,于Syngene凝胶成像仪观察并拍照。
2.4 数据处理
ISSR-PCR扩增产物中,将电泳图谱中较清晰的条带(包括弱带)计为“1”,无带计为“0”,利用POPGENE软件计算等位基因数、Nei′s遗传多样性指数、Shannona信息多样性指数等参数;利用软件NTsys 2.10e分析遗传相似性系数,按UPGMA法构建聚类图[8,9]。
3 结果与分析
3.1 老鹳草属植物的遗传多样性
3.1.1 扩增产物的多态性分析
采用24条随机引物对样品进行扩增筛选,其中8条引物能扩增出带形清晰、多态性较好、清晰度高的扩增产物(表2),扩增条带分布在300~3 000 bp之间(图1)。8条引物共扩增出149条条带,其中多态性条带146条,多态性比率为97.99%,揭示老鹳草属植物在物种水平上保持相当高的遗传多样性。

表2 ISSR-PCR的扩增结果
3.1.2 Shannon信息指数分析和Nei遗传多样性指数分析
利用Popgene 32软件对7种老鹳草居群进行遗传多样性分析。由表3可以看出,老鹳草各个种群的平均Shannon指数和Nei′s基因多样性指数分别在 0.236 4~0.402 9(平均 0.311 1)和 0.140 2~0.267 5(平均0.206 6)范围内,表明老鹳草属植物种群间具有较高的多态性、遗传变异和进化水平。7个种群中,遗传变异水平大小为PopK>PopB>PopM>PopT>PopS>PopL>PopC[10-12]。
3.1.3 老鹳草的遗传结构

图1 部分UBC引物对7种老鹳草样品的ISSR扩增结果 (M为标准分子量DL 5 000)
遗传结构是通过物种居群内和居群间的遗传分化来体现的,根据总的遗传多样性(Ht)和群体内遗传多样性(Hs)计算不同种群间的遗传多样性(Dst,Dst=Ht-Hs)和遗传分化水平(Gst,Gst=Dst/Ht)。7个老鹳草种群间的遗传多样性Dst=0.250 5,分化指数Gst=0.604 2,说明种群间的遗传变异占60.42%,而种群内的遗传变异为39.58%,虽然种群间变异占主要比重,但种群内的变异程度亦相当大;基因流系数Nm为0.327 7,远小于1,说明种群间存在明显的分化[13、14]。
3.2 老鹳草属植物种间亲缘关系分析
利用Ntsys 2.10 e软件计算7个种间的遗传相似系数;采用UPGMA聚类分析方法构建亲缘关系树状聚类图(图2)。结果显示,7种老鹳草在相似系数为0.52处被聚为两类,老鹳草和鼠掌老鹳草被聚为Ⅰ类,其他5种聚为Ⅱ类;在相似系数为0.55处,Ⅱ类又分为3个亚类,即块根老鹳草、突节老鹳草和长白老鹳草为一类,朝鲜老鹳草和毛蕊老鹳草各为一类,其中,块根老鹳草和突节老鹳草的亲缘关系最为接近,相似系数高达0.77。该分类结果与《中国植物志》中对组划分及对原植物形态的分类结果相一致。
4 讨论
4.1 对辽宁省老鹳草属植物的考证
据《中国植物志》记载,北方老鹳草(Geranium erianthum DC.)产于吉林省东部;《东北草本植物志》收录老鹳草属植物共9种,其中分布于辽宁的有6种,另记载北方老鹳草在吉林省多个县均有分布;《辽宁植物志》共收载老鹳草属植物7种,亦未将毛蕊老鹳草(Geranium eriostemon Fish.ex DC.)列入其中,而将Geranium erianthum DC.收录。本实验取材主要基于以下两点考证:一是就现有资料来看,《中国植物志》第43卷成书于1997年,《东北草本植物志》第6卷成书于1977年,《辽宁植物志》成书于1988年。因此笔者推测,《东北草本植物志》对辽宁省仅有6种的记载实为纰漏,成书最晚的《中国植物志》明确收载于《辽宁植物志》的Geranium erianthum DC.一种产于吉林省,该说法与《东北草本植物志》的信息一致,而Geranium erianthum DC.和Geranium eriostemon Fish.ex DC.在亲缘关系上极为相近[10],因此,初步判断《辽宁植物志》对Geranium erianthum DC.很可能为误定。二是在实地考察中,2014,2015年6-9月间我们在辽宁省调查了近200个天然山区,其中包括《辽宁植物志》中对Geranium erianthum DC.有收录的桓仁、宽甸等山区,并未发现Geranium erianthum DC.,而Geranium eriostemon Fish.ex DC.在辽宁山区尤其本溪县内普遍分布较多;后又考察了中国科学院沈阳生态所(中国科学院林业土壤研究所前身)的凭证标本,该标本现无根部、无花果部位,唯有3大3小叶片存留,虽叶裂均至3/4处,但就目前不完整的信息并不能排除是Geranium eriostemon Fish.ex DC.的可能性。此外,我们曾在该所Geranium erianthum DC.标本的标注地区桓仁老秃顶多次进行较大规模的考察,依然未能发现该种,其中同行的有大连自然博物馆张淑梅教授。综合以上两点的考证,笔者可初步断定辽宁省的老鹳草属植物共7种,分 别 为 GeraniumwilfordiiMaxim.、Geranium sibiricum L.、Geranium eranim dahuricum DC.、Geranium krameri Franch.et Sav.Enum.、Geranium koreanum Kom.in Act.、Geranium paishanense Y.L.Chang.和Geranium eriostemon Fish.ex DC.,而更正了《辽宁植物志》中的Geranium erianthum DC.。
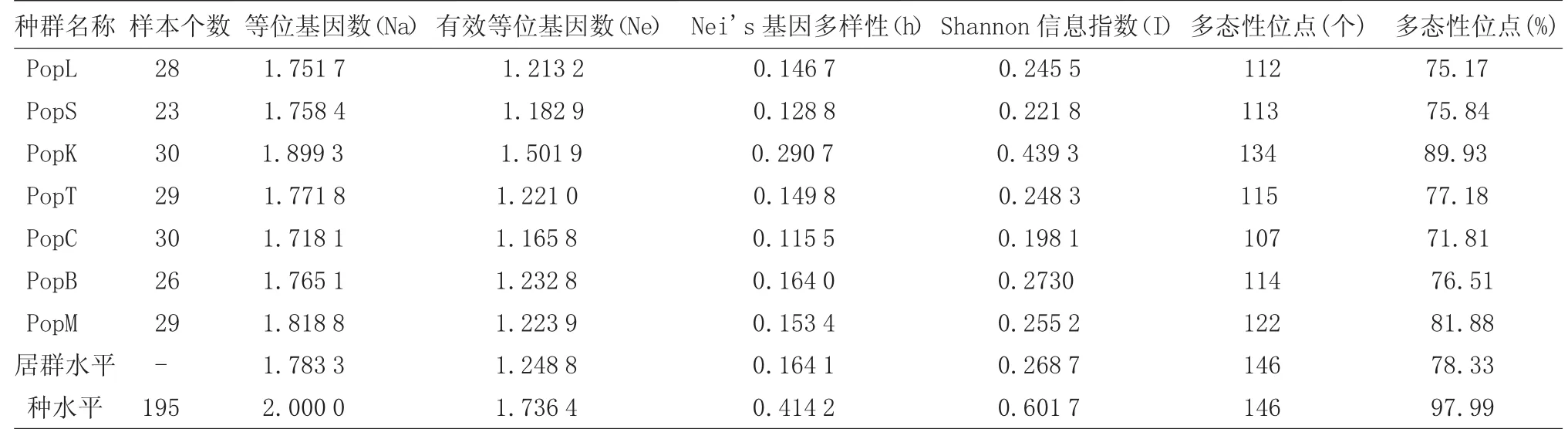
表3 195份老鹳草样品遗传多样性的ISSR分析

表4 7个种间的Nei遗传距离(斜对线上方)和相似系数(斜对线下方)

图2 7种老鹳草基于ISSR的遗传相似性UPGMA聚类图
4.2 ISSR在老鹳草属植物种间亲缘关系分析的适用性
ISSR分子标记技术能在不同的物种间通用,具有操作简单、成本较低、稳定性高、模板DNA用量少、多态性丰富等优点。采用该法对同属不同种间亲缘关系的分析屡见不鲜,而目前在老鹳草属植物中利用ISSR分子标记进行种间亲缘关系的分析尚未见报道。本实验利用ISSR引物的通用性特点,将该分子标记技术应用于辽宁省老鹳草属植物亲缘关系的考察。结果表明,该标记技术在反映老鹳草属植物资源间的亲缘关系和群体间遗传多样性中可行。由聚类结果可知,Ⅰ类均为直根系,Ⅱ类均为块根,在Ⅱ类下依次分出处于高寒地区的毛蕊老鹳草、朝鲜老鹳草和长白老鹳草,生长于草甸的块根老鹳草和突节老鹳草的亲缘关系最为接近,该结果与原植物形态、分布地区的记载完全吻合,也表明老鹳草属植物资源的遗传分化与其来源的生态环境相关联。
4.3 老鹳草属植物的遗传多样性
本研究中,8条ISSR引物的多态性为97.99%,表明该7个种群间具有相当丰富的遗传多样性。遗传变异水平大小顺序为块根老鹳草>长白老鹳草>毛蕊老鹳草>突节老鹳草>鼠掌老鹳草>老鹳草>朝鲜老鹳草,且块根老鹳草种群的Nei's基因多样性(0.290 7)远高于其他6个种群的平均水平(0.143 0)。照原植物形态来讲,块根老鹳草和大花组(直径D>2.0 cm)中的突节老鹳草极为相似,本研究亦证实二者的亲缘关系最为接近,但块根老鹳草却与Ⅰ类中的老鹳草和鼠掌老鹳草同归为小花组(直径D<2.0 cm),笔者猜测形态上的交叉可能是源于块根老鹳草丰富的遗传多样性;长白老鹳草虽多处环境较单一的高寒地区,其遗传变异水平却仅次于块根老鹳草,该结果恰恰证实了《中国植物志》中长白老鹳草是块根老鹳草原变种高寒类型的观点。
[1]刘娟,王良信.老鹳草的本草考证[J].中草药,1992,23(5):276.
[2]国家药典委员会.中华人民共和国药典·一部[S],北京:中国医药科技出版社,2015.122.
[3]中国植物志编委会.中国植物志(43卷第1册)[M].北京:科学出版社,2004.23-26.
[4]李书心.辽宁植物志(上册)[M].辽宁科学技术出版社,1988.1000-1003.
[5]辽宁省林业土壤研究所.东北草本植物志(第6卷)[M].北京:科学技术出版社,1977.11.
[6]王吉华.老鹳草品质评价及遗传多样性研究[D].辽宁中医药大学,2013.75.
[7]陈美霞,朱文宁,孙焕,等.石斛属植物ISSR扩增体系的建立与优化[J].中国农学通报,2014,30(10):177-181.
[8]李先宽,王冰,郑艳超,等.五味子特殊类型白果五味子的分子标记研究[J].中药材,2014,37(4):568-572.
[9]李猛,王平,孙吉康,等.天然蚬壳花椒种群遗传多样性的ISSR标记分析[J].中药材,2014,37(12):2159-2163.
[10]梁呈元,刘艳,李维林,等.薄荷属植物遗传多样性的ISSR分析[J].中药材,2011,34(8):1190-1193.
[11]李绍臣,李凤明,张立民,等.吉林省天然黄檗种群遗传多样性ISSR 分析[J].生态学报,2016,36(1):1-7.
[12]朱田田,晋玲,杜弢,等.甘肃不同产地中麻黄遗传关系的ISSR分析[J].中药材,2013,36(9):1397-1401.
[13] Kunar A,Rogstad SH.A hierarchical analysis of microsatelliteDNAdiversityinGambeloak(Qre ganlbelii Nutt.Fagaceae[J].Mol Ecol,1998,7:859.
[14]Slatkin M.Gene flow in natural populations[J].Ann Rev Ecol Syst,1985,16:393-430.
