长岛周边海域浮游植物分布及其环境影响因素
2016-07-11王媛媛石洪华霍元子
王媛媛,李 捷,李 芬,石洪华,霍元子,3
长岛周边海域浮游植物分布及其环境影响因素
王媛媛1,2,李捷1,李芬2,石洪华2,霍元子2,3
(1.青岛理工大学 环境与市政工程学院,山东 青岛 266033; 2.国家海洋局 第一海洋研究所,山东 青岛266061; 3.上海海洋大学 水产与生命学院,上海 201306)
摘要:2013年春季和夏季对长岛周边海域进行了环境与生物综合调查,利用多元统计分析对长岛周边海域浮游植物分布及其影响因素进行分析。根据水环境分析结果对调查海域进行聚类分析,结果表明,春季(M1、M2)与夏季(A1、A2)均可分为两大类。M1与A2为高温高营养盐区,M2与A1为低温低营养盐区。调查期间,共发现69种浮游植物。硅藻为两季主要浮游植物,春季浮游植物细胞丰度远大于夏季。春季共发现1种明显的优势种,为柔弱几内亚藻(Guinardia delicatula(Cleve)Hasle)。夏季共发现5种优势种,分别为具槽帕拉藻(Paralia sulcata)、裸甲藻(Gymnodinium sp.)、圆筛藻(Coscinodiscus sp.)、离心列海链藻(Thalassiosira eccentrica)与具齿原甲藻(Prorocentrum dentatum)。影响春季浮游植物分布的最佳解释变量组合为温度(WT)、透明度(SD)、风速(WS)和溶解总碳(DTC)组合,其中,温度影响最显著(极显著负相关); 夏季则为盐度(S)、活性磷酸盐(SRP)和 N/Si 组合,其中活性磷酸盐影响最大(显著负相关)。
关键词:浮游植物; 环境因子; 多元统计分析; 春夏季; 长岛
[Foundation: Natural Science Foundation of Shandong Province,No.ZR2013DM003; National Science and Technology Basic Work,No.2012FY112500; Special Fund for Marine Public Welfare Industry Research,No.201505001]
海岛与大陆隔离,相对独立地镶嵌在海洋中,地理位置较特殊,具有海陆二相性,是一种特殊的生态系统[1]。海岛周边海域蕴藏着丰富的渔业、港口、矿产和旅游等资源,是海岛生态系统的重要组成部分。近年来,关于海岛周边海域的研究日益受到重视[2-3]。作者针对长岛周边海域,于2013年5月和8月对其进行了环境与生物综合调查,以期对长岛周边海域有更进一步的认识。长岛纵列于渤海海峡,位于黄渤海交汇处,海流活动频繁[4],毗邻蓬莱经济区,海洋生态环境受人为活动影响显著,导致研究海域浮游植物的区域性变化复杂。从20世纪90年代起,孙军[5-9]对渤海海峡的浮游植物做了大量的工作,1998~1999年在长岛设置了站位,并对其进行了季节演替及影响因子分析[10],之后对于长岛的浮游植物研究很少。以往对于浮游植物影响因素的研究一般集中在物理水文因素和营养盐[2,11-12],且营养盐的研究一般集中在氮磷硅[13-14]对浮游植物的影响。而溶解性碳与其他环境因子的耦合的研究相对较少,本研究将溶解碳列入环境变量中,探索溶解碳对浮游植物的影响。作者通过对长岛周边海域的详细调查与分析,以探明浮游植物的分布特征及影响因素,尤其是碳对浮游植物的影响,为渤海海峡浮游植物调查作补充,并进一步加深对近海海域浮游植物的认识。
1 材料与方法
1.1站位布设与样品采集分析
样品采集于2013年5月10日和8月16日,根据各个调查站位水深情况按照《海洋调查规范》[2007]采集样品,在长岛周边海域设置21个站位(图1)。水深为5~27 m。用透明度盘测量水体透明度。水温、pH、溶解氧和盐度等环境参数用便携式多参数-水质分析仪(MS5,HACH)在现场测定。用于测定其他水质指标的水样经低温保存后带回实验室进行检测。水体悬浮物(SS)测量应用重量法。用于测定NO2-N、NO3-N、NH4-N、SRP和SiO3-Si 的水样经0.45 µm醋酸纤维滤膜过滤后,按照APHA[15]方法进行。亚硝酸盐浓度较氨氮、硝酸盐质量浓度较低,因此,将三者合并为无机氮(DIN)。用于检测浮游植物的水样立即转移至0.5L的PE(聚乙烯)瓶中,加入终浓度为5%的甲醛水溶液,于常温避光保存。浮游植物样品分析运用 Utermöhl[16]方法,依据个体大小和形态特征对照浮游植物鉴定手册进行种类鉴定。浮游植物细胞丰度用个/m3表示。
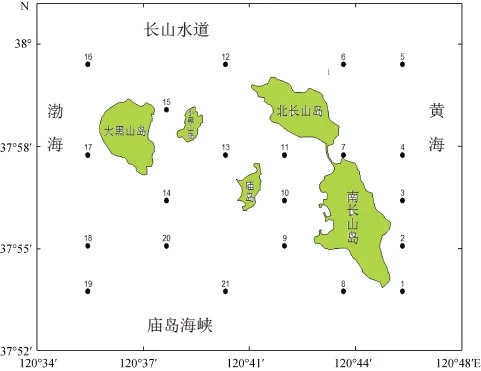
图1 长岛周边海域位置及调查站位Fig.1 Location of surrounding waters of Changdao Island and sampling stations
使用 Surfer 8.0 绘制空间分布图。数据分析在SPSS与 PRIMER v6软件包上进行,通过BIOENV分析对浮游植物丰度与环境因子的关系进行分析,找出可最佳解释群落结构的环境变量组合[18]。然后,通过 Pearson相关分析确定影响浮游植物分布的最佳环境因子组合中每一个环境因子的影响大小。
优势种由公式
2 结果与讨论
2.1水环境特征
聚类分析结果(图2)表明,根据水环境分析结果,春季与夏季均可分为两大类。春季分为M1(1、5、6、8、9、10、11、13、19、21)与M2(2、3、4、7、12、14、15、16、17、18、20),夏季分为A1(1、2、3、4、7、11、12、13、15)与A2(5、6、8、9、10、14、16、 17、18、19、20、21)。M1与M2、A1与A2之间具有显著的差异(表1)。将M1、M2与A1、A2对比发现,M1与A2所包含的区域相似,M2与A1所包含的区域相似,但经对比发现季节之间有明显的差异。
夏季营养盐浓度显著高于春季,这是因为夏季入海径流增多,带来丰富的营养盐。同时,由于降水量的增加,导致夏季调查海域pH的上升和盐度的减小(表1)。春季溶解总碳的浓度大于夏季。本次调查中溶解氧的浓度均大于 6 mg/L,大于一类海水标准,因此未考虑溶解氧的影响。
春季,M1的温度比M2较高; M1的DIN浓度范围为 0.0560~0.1230 mg/L,M2的为 0.0322~0.0484 mg/L,M1的DIN浓度显著大于M2。夏季,A2的温度比A1较高; A2的DIN质量浓度(0.1257~0.1584 mg/L)显著高于A1(0.0929~0.1380 mg/L)。M1与 A2营养盐的质量浓度(除硅酸盐)均较低。调查期间,4个区域的N/P均大于16。只有M1的N/Si大于1,其余3个区域均小于1。综上所述,可将M1与A2概括为高温高营养盐区,M2与A1为低温低营养盐区。
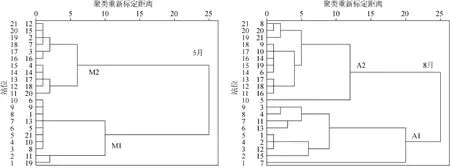
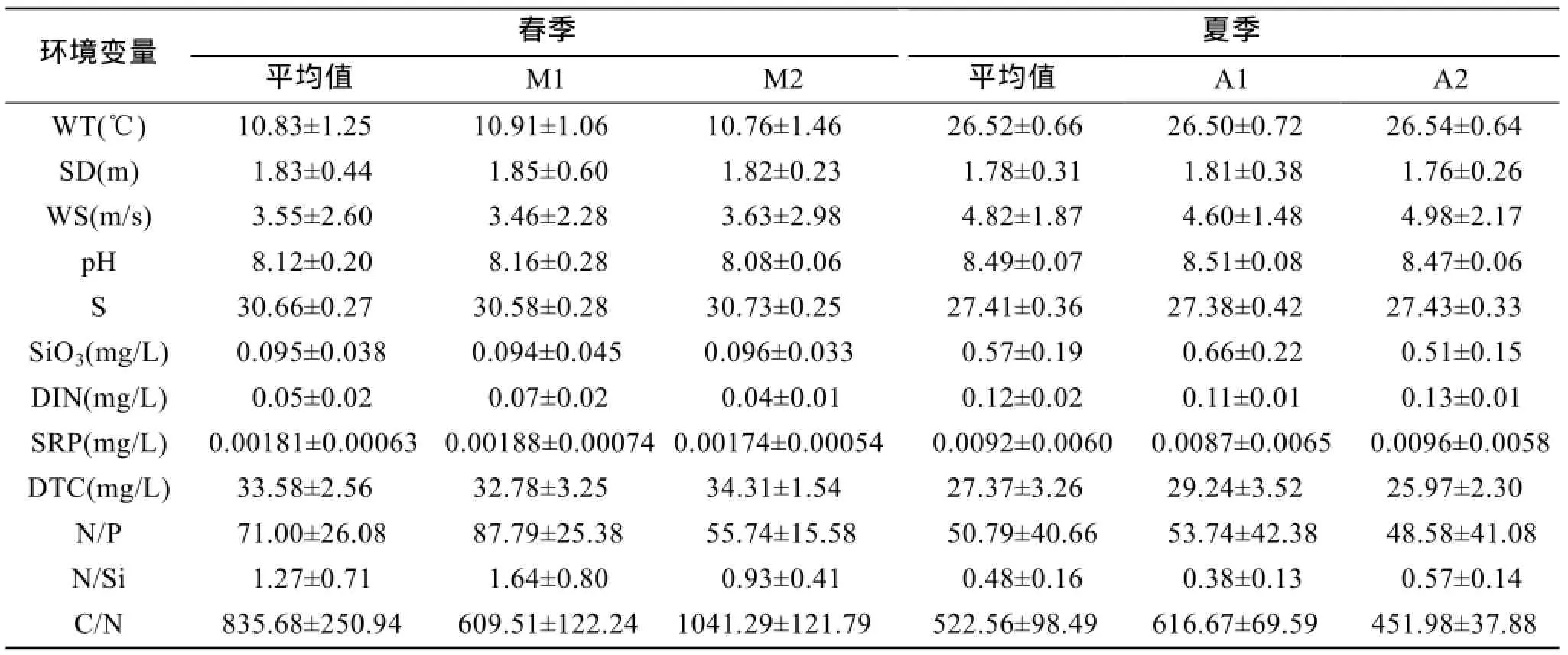
表1 长岛周边海域水环境参数平均值Tab.1 Means(and standard error)of environmental parameters in waters surrounding Changdao Island
运用组间平均连锁聚类法进行聚类
Dendrogram using average linkage(between groups)
2.2浮游植物分布特征
2.2.1浮游植物组成
调查期间,共发现69种浮游植物(浮游植物中文名和拉丁名命名参考[19]),其中,春季 41种,夏季41种。两季中硅藻所占的比例均高于 50%,春季硅藻比例(75.61%)大于夏季(51.22%)。无论是春季还是夏季,硅藻均是该海域的主要浮游植物群落。单一站位单种占该站位总数量的 5% 以上的浮游植物春季有14种,夏季有22种。
2.2.2浮游植物细胞丰度
春季,浮游植物细胞丰度为 2.47×108~2.78× 109个/m3,平均为7.11×108个/m3。硅藻细胞丰度高达总细胞丰度的99.995%。M2细胞丰度(9.19×108个/m3)远大于M1细胞丰度(4.83×108个/m3)。
夏季,浮游植物细胞丰度为 0.29×106~3.67× 106个/m3,平均为1.45×106个/m3。硅藻细胞丰度占总细胞丰度的 61.63%。A1的细胞丰度(1.450×106个/m3)与A2的细胞丰度(1.453×106个/m3)相差不大。
春季浮游植物细胞丰度远大于夏季。两季硅藻细胞丰度均占总细胞丰度的 50%以上,说明硅藻是该调查海域的重要浮游植物,尤其是春季,几乎全部均为硅藻,只有少数甲藻与金藻,金藻只有一种为小等刺硅鞭藻(Dictyocha fibula Ehrenberg)。从空间分布来看,春季差异很明显,远离海岸的海域(M2)浮游植物细胞丰度较高,靠近海岸的海域及东北角海域(M1)则较低。夏季,浮游植物分布较均匀,仅西北角海域浮游植物细胞丰度较小(图3)。

图3 浮游植物细胞丰度分布Fig.3 Distribution of phytoplankton cell abundance
2.2.3优势种
春季仅发现 1种明显的优势种,为柔弱几内亚藻(Guinardia delicatula(Cleve)Hasle)。由表2可知,柔弱几内亚藻以绝对优势成为春季优势种,无论是在 M1还是在 M2海域,柔弱几内亚藻均为优势种,但 M2海域(21.64×109个/m3)比 M1 海域(11.22×109个/m3)多(图 4)。夏季共发现 5种优势种,分别为具槽帕拉藻(Paralia sulcata)、裸甲藻(Gymnodinium sp.)、圆筛藻(Coscinodiscus sp.)、离心列海链藻(Thalassiosira eccentrica)与具齿原甲藻(Prorocentrum dentatum)(表2),占浮游植物细胞丰度的比例为74.98%。夏季,优势种主要分布在A1海域(图 4)。夏季未出现柔弱几内亚藻,而春季未出现裸甲藻和具齿原甲藻。春季,具槽帕拉藻、圆筛藻和离心列海链藻在M1与M2的分布相当,无太大差别,丰度均较小。
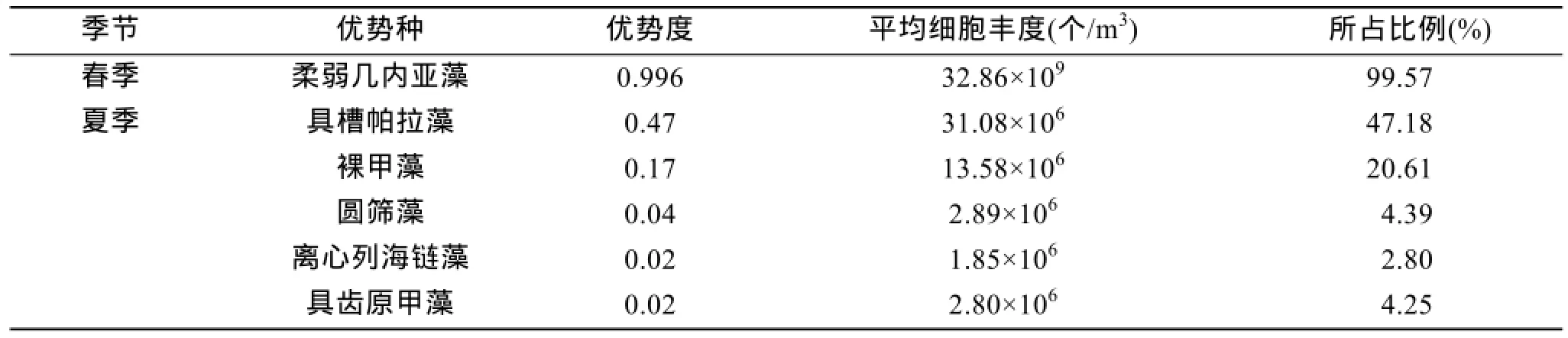
表2 浮游植物优势种及其数量Tab.2 Dominant species and amount of phytoplankton
2.3浮游植物分布与环境因子的关系
2.3.1春季浮游植物分布与环境因子的关系
通过BIOENV分析(表3)可知,温度(WT)、透明度(SD)、风速(WS)和溶解总碳(DTC)组合为影响春季浮游植物分布的最佳解释变量组合,其相关系数为0.624; 其次为温度(WT)、透明度(SD)、风速(WS)、盐度(S)和溶解总碳(DTC)组合,其相关系数为0.623。Global Test 检验的 Rho=0.624,显著度为0.01。另外,本研究对春季浮游植物优势种柔弱几内亚藻与环境因子的关系单独做了 BIOENV分析(表3)。由表3可知,影响柔弱几内亚藻分布的最佳环境因子组合与影响春季浮游植物分布的最佳环境因子组合一样,只是相关系数不同,前者的相关系数为0.573。Global Test检验的Rho=0.573,显著度为0.01。通过Pearson相关分析可知,春季浮游植物和优势种柔弱几内亚藻与最佳环境因子组合的相关分析相似,与温度均呈极显著负相关,与风速呈显著正相关,与透明度和溶解碳分别呈现一定程度的负相关和正相关,此结果从侧面反映了春季柔弱几内亚藻对整个群落的影响是很大的。
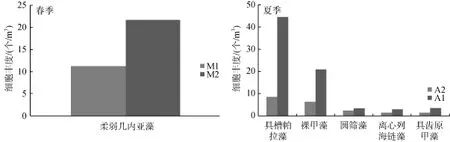
图4 优势种分布Fig.4 Distribution of dominant species

表3 春季浮游植物及优势种与环境因子的相关分析结果Tab.3 Results of BIOENV between phytoplankton(dominant species)cell abundance and environmental factors in spring
优势种是对整个群落具有控制性影响的种群,因此优势种与环境因子的关系能够代表整个群落与环境因子的关系。柔弱几内亚藻为温带近岸种,细胞个体较小,分布在我国渤海、黄海、东海和南海海域[20-21]。本研究中,柔弱几内亚藻在春季已达到赤潮浓度水平,且其丰度与温度呈显著负相关(表4)。本研究春季平均温度为10.83℃,温度较低,柔弱几内亚藻较适于生长。曾有报道指出2004年2月胶州湾也曾发生了柔弱几内亚藻赤潮[20],因此作者推测柔弱几内亚藻是低温种,且温度越低越有利于其生长。该海域水深较浅,春季风速较大,而较大的风速通过再悬浮可将底层营养盐带至表层,从而有利于浮游植物的生长[22-23]。但是营养盐的再悬浮导致海水透明度降低,从而影响浮游植物光合作用。浮游植物在光合作用过程中会释放溶解有机碳[24-25],且赵卫红等[20]的研究表明,柔弱几内亚藻与溶解有机碳呈正相关,与本研究结果一致。

表4 春季浮游植物及优势种Pearson相关分析结果Tab.4 Pearson correlation analysis of phytoplankton(dominant species)abundance and environmental factors in spring
2.3.2夏季浮游植物分布与环境因子的关系
通过BIOENV分析(表5)可知,最能对庙岛群岛南部海域浮游植物丰度分布差异做出解释的组合是盐度(S)、活性磷酸盐(SRP)和 N/Si 组合,其相关系数为0.354; 其次为透明度(SD)、盐度(S)、活性磷酸盐(SRP)和 N/Si组合,其相关系数为 0.353。Global Test检验的Rho=0.354,显著度为0.04。由 Pearson相关分析结果可知,影响夏季浮游植物分布的最佳环境因子组合中 N/Si 的影响很小,因此重点分析盐度和活性磷酸盐对浮游植物分布的影响。
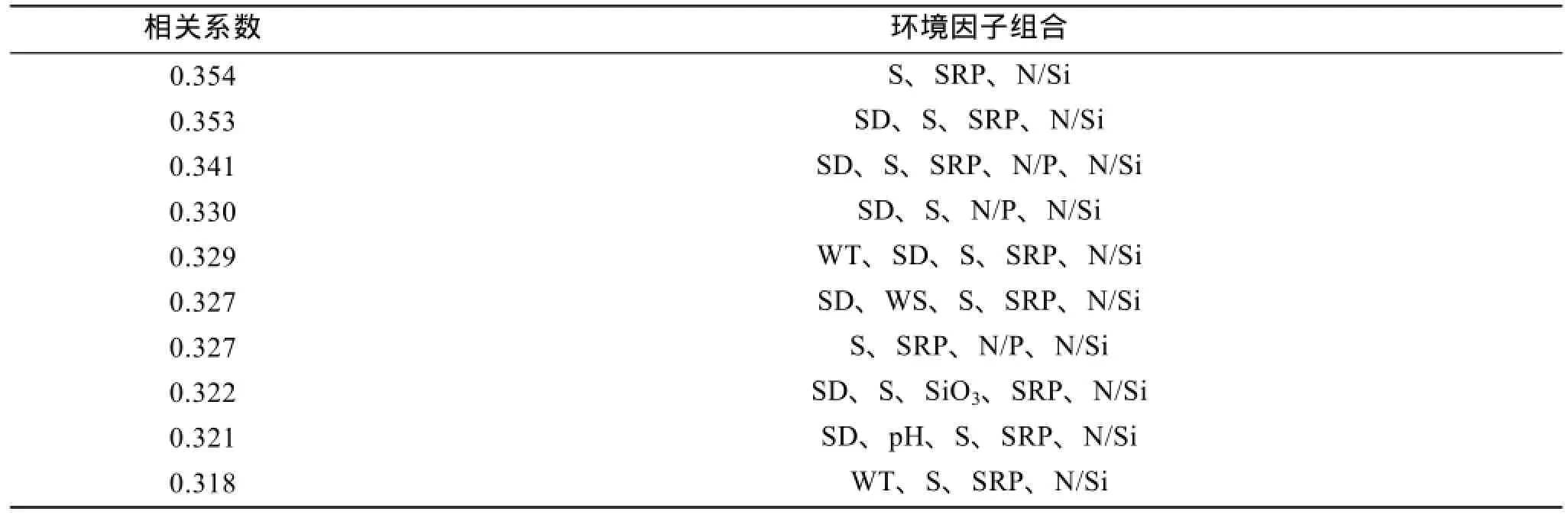
表5 夏季浮游植物与环境因子的相关分析结果Tab.5 Results of BIOENV between phytoplankton cell abundance and environmental factors in summer
表6为Pearson相关分析结果。由表6可知,夏季浮游植物与活性磷酸盐呈显著负相关。以往的研究结果[26-27]表明,当海水中Si: P>22和N: P>22时,磷酸盐为限制因子。本研究中夏季Si: P=105.81、N:P=50.79,二者比值均远大于 22,因此该海域夏季磷酸盐为其限制因子。同时由于入海径流和降雨的增多导致盐度较低,由图4可知,夏季优势种主要分布在A1海域,A1海域的盐度相对于A2较低,因此,该海域夏季浮游植物喜盐度较低的环境,从而其分布与盐度呈一定程度正相关。

表6 夏季浮游植物Pearson相关分析结果Tab.6 Pearson correlation analysis of phytoplankton abundance and environmental factors in summer
2.3.3浮游植物季节分布与环境因子的关系
夏季甲藻物种数占总物种数的 46.34%,而春季没有鉴定出甲藻种类。此结果符合温带海域浮游植物群落的一般变化趋势,即甲藻适于在温度较高的夏季生长[12,28]。本研究中的季节更替非常明显,春季适于在较低温度下生长的柔弱几内亚藻到夏季的时候基本不存在,而被适于在较高温度下生长的具槽帕拉藻、圆筛藻和离心列海链藻等硅藻与裸甲藻、具齿原甲藻等甲藻而代替。这进一步证明温度是影响浮游植物季节更替的最重要的环境因子[29-30]。
通过BIOENV分析和Pearson相关分析可知,夏季浮游植物主要受磷酸盐的限制,因此浮游植物的生长受限; 而对春季的分析结果中是物理水文因素占主导,营养盐对其影响不大。夏季虽然营养盐较春季丰富,但浮游植物是按照一定比例吸收营养盐的,这一恒定比例被称为Redfield比值[31]。Redfield比值为C: N: P=106: 16: 1[32]。尽管两季的氮磷比均大于16/1,但春季氮磷比大于夏季(表 1),而高氮磷比可促进硅藻的生长[33],因此,春季硅藻的细胞丰度远高于夏季。两季中的碳氮比均远大于106/16,而分析结果中氮不是研究海域的限制性因子,因此,此现象很有可能是因为研究海域中溶解碳的含量很高,从而可推测,该海域的生产力较高。
其次,盐度同样也会在一定程度上影响浮游植物的季节更替[34-35]。虽然春季盐度高于夏季(表 1),但两季的盐度相差不大(3.25个单位),因此,在BIOENV分析和 Pearson 相关分析中盐度并未成为显著影响因子。
3 结论
1)聚类分析结果(图 2)表明,根据水环境分析结果,春季与夏季均可分为两大类。春季分为M1(1、5、6、8、9、10、11、13、19、21)与M2(2、3、4、7、12、14、15、16、17、18、20),夏季分为A1(1、2、3、4、7、11、12、13、15)与 A2(5、6、8、9、10、14、16、17、18、19、20、21)。M1与A2为高温高营养盐区,M2与A1为低温低营养盐区。
2)调查期间共发现69种浮游植物,其中,春季41种,夏季41种。春季浮游植物细胞丰度远大于夏季。两季硅藻细胞丰度均占总细胞丰度的 50% 以上,是该海域的浮游植物群落最主要的组成类群。
3)影响春季浮游植物分布的最佳解释变量组合为温度(WT)、透明度(SD)、风速(WS)和溶解总碳(DTC)组合,其中,温度影响最显著(极显著负相关),风速次之(显著正相关); 夏季则为盐度(S)、活性磷酸盐(SRP)和 N/Si组合,其中活性磷酸盐影响最大(显著负相关)。物理水文因素对春季浮游植物的分布影响较显著,而夏季则为营养盐。
4)春季优势种仅有柔弱几内亚藻(Guinardia delicatula(Cleve)Hasle)1种。夏季共发现 5 种优势种,分别为具槽帕拉藻(Paralia sulcata)、裸甲藻(Gymnodinium sp.)、圆筛藻(Coscinodiscus sp.)、离心列海链藻(Thalassiosira eccentrica)与具齿原甲藻(Prorocentrum dentatum)(表2)。
参考文献:
[1]石洪华,郑伟,丁德文,等.典型海岛生态系统服务及价值评估[J].海洋环境科学,2009,28(6): 743-748.Shi Honghua,Zheng Wei,Ding Dewen,et al.Valuation and ecosystem services of typical island[J].Marine Environmental Science,2009,28(6): 743-748.
[2]Youg M C,Joon T Y,Jung H C,et al.Ecosystem structure and trophic level to the oceanographic conditions around the waters of Jeju Island[J].Journal of Environmental Biology,2008,29(4): 419-425.
[3]Christian S R,Christopher D H,Osmund Holm-Hansen.Influence of atmospheric teleconnections andupper circumpolar deep water on phytoplankton biomass around Elephant Island,Antarctica[J].Marine Ecology Progress Series,2009,377: 51-62.
[4]张乃星,吴凤丛,任荣珠,等.渤海海峡冬季表层海
水中溶解无机碳分布特征分析[J].海洋科学,2012,36(2): 56-61.
Zhang Naixing,Wu Fengcong,Ren Rongzhu,et al.Distribution analysis of dissolved inorganic carbon in surface water around the Bohai Strait in winter[J].Marine Sciences,2012,36(2): 56-61.
[5]孙军,刘东艳,杨世民,等.渤海中部和渤海海峡及
邻近海域浮游植物群落结构的初步研究[J].海洋与湖沼,2002,33(5): 461-471.Sun Jun,Liu Dongyan,Yang Shimin,et al.The preliminary study on phytoplankton community structure in the central Bohai Sea and the Bohai Strait and its adjacent area[J].Oceanologia Et Limnologia Sinica,2002,33(5): 461-471.
[6]孙军,刘东艳,张晨,等.渤海中部和渤海海峡及其邻近海域浮游植物粒级生物量的初步研究Ⅰ.浮游植物粒级生物量的分布特征[J].海洋学报,2003,25(5): 103-112.Sun Jun,Liu Dongyan,Zhang Chen,et al.The preliminary study on size-fractionated biomass of phytoplankton in the central Bohai Sea,the Bohai Straits and its adjacent area Ⅰ.The distribution of phytoplankton size-fractionated biomass[J].Acta Oceanologica Sinica,2003,25(5): 103-112.
[7]孙军,刘东艳,柴心玉,等.1998~1999年春秋季渤海中部及其邻近海域叶绿素a浓度及初级生产力估算[J].生态学报,2003,23(3): 517-526.Sun Jun,Liu Dongyan,Chai Xinyu,et al.The chlorophyll a concentration and estimating of primary productivity in the Bohai Sea in1998-1999[J].Acta Ecologica Sinica,2003,23(3): 517-526.
[8]孙军,刘东艳,王威,等.1998年秋季渤海中部及其邻近海域的网采浮游植物群落[J].生态学报,2004,24(8): 1643-1655.Sun Jun,Liu Dongyan,Wang Wei,et al.The netz-phytoplankton community of the central Bohai Sea and its adjacent waters in autumn,1998[J].Acta Ecologica Sinica,2004,24(8): 1643-1655.
[9]Liu D Y,Sun J,Liu Z,et al.The effects of spring-neap tide on the phytoplankton community development in the Jiaozhou Bay,China[J].Acta Oceanologica Sinica,2004,23(4): 687-697.
[10]Sun J,Liu D Y,Qian S B.Preliminary study on the seasonal succession and development pathway of phytoplankton community in the Bohai Sea[J].Acta Oceanologica Sinica,2001,20(2): 251-260.
[11]Jiang Y J,He W ,Liu W X,et al.The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake(Lake Chaohu)[J].Ecological Indicators,2014,40: 58-67.
[12]傅明珠,孙萍,孙霞,等.锦州湾浮游植物群落结构特征及其对环境变化的响应[J].生态学报,2014,34(13):3650-3660.Fu Mingzhu,Sun Ping,Sun Xia,et al.Phytoplankton community structure and its ecological responses to environmental changes in Jinzhou Bay,China.Acta Ecologica Sinica,2014,34(13): 3650-3660.
[13]Sandra M,Friso M,Melanie B,et al.Dominance of the non-indigenous diatom Mediopyxis helysia in Wadden Sea phytoplankton can be linked to broad tolerance to different Si and N supplies[J].Journal of Sea Research,2015,95: 36-44.
[14]Juliette L Y,Catharina J M P,Jacco C K.Phosphorus limitation during a phytoplankton spring bloom in the western Dutch Wadden Sea[J].Journal of Sea Research,2014,88: 109-120.
[15]APHA.Standard methods for the examination of water and wastewater,20th ed[S].Washington DC: APHAAWWA-WPCF,1998.
[16]孙军,刘东艳,钱树本.一种海洋浮游植物定量研究分析方法——Utermöhl方法的介绍及其改进[J].黄渤海海洋,2002,20(2): 105-112.Sun Jun,Liu Dongyan,Qian Shuben.A Quantative Research and Analysis Method for Marine Phytoplankton: An Introduction to Utermöhl Method and Its Modification[J].Journal of Oceanography of Huanghai & Bohai Seas,2002,20(2): 105-112.
[17]Lampitt R S,Wishner K F,Turley C M,et al.Marine snow studies in the northeast Atlantic: distribution,composition and roles as a food source for migrating plankton[J].Marine Biology,1993,116(4): 689-702.
[18]Clarke K R,Gorley R N.PRIMER v6: User Manual/Tutorial [M].Plymouth: PRIMER-E Ltd.2006.
[19]孙军,刘东艳.中国海区常见浮游植物种名更改初步意见[J].海洋与湖沼,2002,33(3): 271-286.Sun Jun,Liu Dongyan.The preliminary notion on nomenclature of common phytoplankton in China seas waters[J].Oceanologia Et Limnologia Sinica,2002,33(3): 271-286.
[20]赵卫红,崔鑫,王江涛,等.赤潮水体中胶体物质对赤潮异弯藻(Heterosigma akashiwo)和中肋骨条藻(Skeletonema costatum)生长的影响[J].生态学报,2009,29(2): 573-580.Zhao Weihong,Cui Xin,Wang Jiangtao,et al.Effect of colloidal substances in red-tide seawater on the growth of Heterosigma akashiwo and Skeletonema costatum[J].Acta Ecologica Sinica,2009,29(2): 573-580.
[21]尹翠玲,张秋丰,曹春晖,等.2012年春季渤海湾天津近岸海域网采浮游植物群落结构初探[J].海洋学研究,2013,31(4): 80-89.Yin Cuilin,Zhang Qiufeng,Cao Chunhui,et al.Netphytoplankton community in the Tianjin nearshore waters of Bohai Bay in spring of 2012[J].Journal of Marine Sciences,2013,31(4): 80-89.
[22]Xu Y,Cahill B,Wilkin J,et al.Role of wind in regulating phytoplankton blooms on the Mid-Atlantic Bight[J].Continental Shelf Research,2013,63: 26-35.
[23]孙小静,秦伯强,朱广伟,等.风浪对太湖水体中胶体态营养盐和浮游植物的影响[J].环境科学,2007,28(3): 506-511.Sun Xiaojing,Qin Boqiang,Zhu Guangwei,et al.Effect of Wind-Induced Wave on Concentration of Colloidal Nutrient and Phytoplankton in Lake Taihu[J].Environmental Science,2007,28(3): 506-511.
[24]Berman T,Holm-Hansen O.Release of photoassimilated carbon as dissolved organic matter by marine phytoplankton[J].Marine Biology,1974,28(4): 305-310.
[25]刘诚刚.中国近海浮游植物光合溶解有机碳生产研究[D].中国海洋大学,2012.Liu Chenggang.Photosynthetically produced Ddissolved organic carbon released of marine phytoplankton in china coastal seas[D].Ocean University of China,2012.
[26]Justic D,Rabalais N N,Turner R E.Stoichiometric nutrient balance and origin of coastal eutrophication[J].Marine Pollution Bulletin,1995,30(1): 41-46.
[27]Dortch Q,Whitledge T E.Does nitrogen or silicon limit phytoplankton production in the Mississippi River plume and nearby regions[J].Continental Shelf Research,1992,12: 1293-1309.
[28]王朝晖,陈菊芳,徐宁,等.大亚湾裸甲藻种群的季节变化与环境条件的关系[J].生态学报,2001,21(11):1825-1832.Wang Chaohui,Chen Jufang,Xu Ning,et al.Relationship between seasonal variations in Gymnodinium spp.population and environmental factors in Daya Bay,the South China Sea[J].Acta Ecologica Sinica,2001,21(11): 1825-1832.
[29]Santoro A E,Nidzieko N J,Van D G L.Contrasting spring and summer phytoplankton dynamics in the nearshore Southern California Bight[J].Limnology and Oceanography,2010,55(1): 21.
[30]Ke Z X,Xie P,Guo L G.Controlling factors of spring-summer phytoplankton succession in Lake Taihu(Meiliang Bay,China)[J].Hydrobiologia,2008,607(1): 41-49.
[31]王保栋.黄海和东海营养盐分布及其对浮游植物的限制[J].应用生态学报,2003,14(7): 1122-1126.Wang Baodong.Nutrient distributions and their limitation on phytoplankton in the Yellow Sea and the East China Sea[J].Chinese Journal of Applied Ecology,2003,14(7): 1122-1126.
[32]Redfield A C,Ketchum B H,Rechards F A.The influence of organisms on the composition of seawater[A].Hill M N.The Sea.Vol.2[C].New York: Interscience,1963,26-77.
[33]Silkin V A,Pautova L A,Pakhomova S V,et al.Environmental control on phytoplankton community structure in the NE Black Sea[J].Journal of Experimental Marine Biology and Ecology,2014,461: 267-274.
[34]Marijeta Č,Marina C,Frano K,et al.Controlling factors of phytoplankton seasonal succession in oligotrophic Mali Ston Bay(south-eastern Adriatic)[J].Environmental monitoring and assessment,2013,185(9):7543-7563.
[35]Melissa R.Mcquoid.Influence of salinity on seasonal germination of resting stages and composition of microplankton on the Swedish west coast[J].Marine Ecology Progress Series,2005,289(3): 151-163.
(本文编辑: 梁德海)
Phytoplankton distribution and its environmental impact factors in waters surrounding Changdao Island
WANG Yuan-yuan1,2,LI Jie1,LI Fen2,SHI Hong-hua2,HUO Yuan-zi2,3
(1.College of Environmental and Municipal Engineering,Qingdao Technology University,Qingdao 266033,China; 2.The First Institute of Oceanography,State Oceanic Administration,Qingdao 266061,China;3.College of Aquaculture and Life,Shanghai Ocean University,Shanghai 201306,China)
Received: Aug.28,2015
Key words:phytoplankton; environmental factors; multivariate statistical analysis; spring and summer; Changdao Island
Abstract:Environmental and biological investigation were conducted in waters surrounding Changdao Island in the spring and summer of 2013.The distribution of phytoplankton and its environmental impact factors were investigated in multivariate statistical analyses.According to the results of investigation and analysis of this water environment by cluster analysis,the waters surrounding Changdao Island could be divided into two categories in spring(M1and M2)and in summer(A1 and A2).A total of 69 taxa were identified.Diatom was the main phytoplankton in both seasons.The abundance of phytoplankton in spring was greater than that in summer.There was one dominant species,Guinardia delicatula(Cleve)Hasle,in spring.There were five dominant species(Paralia sulcata,Gymnodinium sp.,Coscinodiscus sp.,Thalassiosira eccentric,and Prorocentrum dentatum)in summer.The best explanations among the variables affecting the distribution of phytoplankton in spring were temperature(WT)(very significant negative correlation),transparency(SD),wind speed(WS),and total dissolved carbon(DTC); in summer,the corresponding variables were salinity(S),soluble reactive phosphate(SRP)(significant negative correlation),and N/Si.
中图分类号:Q948.8
文献标识码:A
文章编号:1000-3096(2016)03-0023-09
doi:10.11759/hykx20150603003
收稿日期:2015-08-28; 修回日期: 2015-10-10
基金项目:山东省自然科学基金项目(ZR2013DM003); 国家科技基础性工作专项项目(2012FY112500)海洋公益性行业科研专项项目(201505001)
作者简介:王媛媛(1991-),女,硕士研究生,主要从事海洋环境与生态研究,E-mail: 18766215168@163.com; 李捷,通信作者,E-mail:fstum@tom.com
