半滑舌鳎AKT-ineracting protein基因的克隆及免疫应答表达分析
2016-06-29孙璐明于孟君陈亚东陈学杰刘洋仇雪梅沙珍霞大连海洋大学水产与生命学院大连6023中国水产科学研究院黄海水产研究所农业部海洋渔业可持续发展重点实验室青岛26607青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室青岛266200上海海洋大学水产与生命学院上海20306
孙璐明于孟君陈亚东陈学杰刘 洋仇雪梅沙珍霞(.大连海洋大学水产与生命学院,大连 6023; 2.中国水产科学研究院黄海水产研究所农业部海洋渔业可持续发展重点实验室,青岛 26607; 3.青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室,青岛 266200; 4.上海海洋大学水产与生命学院,上海 20306)
半滑舌鳎AKT-ineracting protein基因的克隆及免疫应答表达分析
孙璐明1,2于孟君1,2陈亚东2,3陈学杰2,4刘 洋1仇雪梅1沙珍霞2,3
(1.大连海洋大学水产与生命学院,大连 116023; 2.中国水产科学研究院黄海水产研究所农业部海洋渔业可持续发展重点实验室,青岛 266071; 3.青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室,青岛 266200; 4.上海海洋大学水产与生命学院,上海 201306)
摘要:研究克隆了半滑舌鳎Cynoglossus semilaevis膜蛋白AKT-interacting protein(AKTIP)基因,研究了其在健康组织中的表达模式、鳗弧菌(Vibrio anguillarum)感染后免疫组织和不同病原物刺激外周血淋巴细胞中的表达特征。通过常规克隆和RACE技术,获得的半滑舌鳎AKTIP基因全长cDNA序列为1224 bp,其中包括5′-UTR为116 bp,3′-UTR为117 bp和完整的ORF序列891 bp,编码296个氨基酸,预测蛋白质的等电点(PI)为9.12,分子量是33.74 kD; 同源比对发现半滑舌鳎AKTIP的氨基酸序列在不同物种之间具有较高的保守性; 荧光实时定量PCR(qRT-PCR)检测到半滑舌鳎各个组织中均有AKTIP基因的表达,在卵巢中表达量最高; 鳗弧菌感染半滑舌鳎后,AKTIP基因在肝、鳃、血液、肠、头肾和脾中均上调表达; 病原模拟物PGN、LPS、poly I:C和WGP刺激半滑舌鳎外周血淋巴细胞均诱导AKTIP基因下调表达。研究表明半滑舌鳎AKTIP基因参与了机体免疫反应,为给半滑舌鳎免疫防御技术的研究提供理论依据。
关键词:半滑舌鳎; AKT-interacting protein基因; 基因克隆; 免疫应答; 基因表达
AKT-interacting protien(AKTIP),又称蛋白激酶B相互作用蛋白,是PI3K(磷酸肌醇3-激酶)/ PDK1(3-磷酸肌醇依赖性蛋白激酶)/ Akt信号通路中第一个被发现的支架蛋白[1,2],参与PI3K/PDK1/ Akt信号通路的调控,是介导Akt与其上游激酶PDK1相互作用的桥梁[3,4],在动物细胞信号转导中具有重要的作用,并参与许多生物学过程,如胚胎发育、细胞的增殖、分化、凋亡、免疫防御等[5]。有文献报道AKTIP也可以通过活化NF-kB 参与免疫调节[6]。到目前为止,仅有数篇关于AKTIP的研究文献,大多集中在哺乳动物和人中,但对AKTIP免疫方面的研究还不透彻,在鱼类中至今没有关于AKTIP的任何报道。
半滑舌鳎(Cynoglossus semilaevis)是我国重要的海水养殖鱼类,病害问题特别是弧菌病对半滑舌鳎养殖业造成严重的经济损失[7]。对半滑舌鳎免疫机制的研究有助于病害问题的解决。本研究拟对AKTIP基因的鉴定、表达及在免疫反应中的应答模式进行分析,以期为半滑舌鳎病害防御提供理论参考依据。
1 材料与方法
1.1 实验材料
半滑舌鳎购自日照东鑫现代渔业技术研究所,鱼为1.5龄左右,平均体重220 g,平均体长30.6 cm。实验前将鱼在24℃水箱中暂养7d,消除环境胁迫。鳗弧菌(Vibrio anguillarum)由本实验室保存,外周血淋巴细胞由本实验室分离获得。
1.2 样品采集
取3条健康半滑舌鳎进行健康组织收集,分别采集后肾、脑、胃、脾、头肾、心脏、鳃、血、肌肉、皮肤、肝、肠、卵巢13种组织,立即投入液氮中冷冻,然后转移至-80℃分别保存,以备RNA提取。
鳗弧菌感染方法参照Sha等[8]略作修改:设实验组和对照组,实验组用3.18×105CFU/g(半致死剂量,LD50)对半滑舌鳎进行腹腔注射,对照组参照实验组注射与体重相应剂量的PBS注射。在注射后0、6h、12h、24h、48h和72h各取3尾鱼,取鳃、肝、肠、头肾、脾、血液6种免疫组织,并立即投入液氮中,随后将其转移至-80℃冰箱中分别保存,用于提取总RNA。
分别用病原模拟物脂多糖(LPS)、肽聚糖(PGN)、多聚胞甘酸(poly I:C)和葡聚糖(WGP)刺激6孔细胞培养板中接种的半滑舌鳎外周血淋巴细胞,使终浓度分别为50、100、50和50 µg/mL,PBS组设为对照组,每个实验重复3次。感染30min后收集细胞悬液,1000 r/min离心5min去除包含感染源的培养基,用Hank's缓冲液反复洗涤细胞,继续用新鲜DMEM培养基重悬细胞进行培养。分别收集感染0、2h、6h、12h和24h后的淋巴细胞到相应的离心管中。离心后倒掉培养基并加入1 mL Trizol试剂(Invitrogen,美国),然后储存在-80℃冰箱中,以备提取总RNA。
1.3 总RNA提取及cDNA的合成
使用试剂盒(Tiangen,中国)提取半滑舌鳎不同组织样本的总RNA。提取方法参照试剂盒说明书。体外培养的外周血淋巴细胞RNA提取采用Trizol试剂提取。总RNA使用Agilent 2100生物分析仪(Applied Biosystems,美国)检测其质量和浓度,RNA的完整性用琼脂糖凝胶电泳法检测。按照PrimeScriptTM RT reagent Kit(Perfect Real Time)(TaKaRa,大连)试剂盒合成cDNA,-20℃保存备用。
1.4 半滑舌鳎AKTIP基因cDNA序列的获得
通过对半滑舌鳎转录组和基因组数据进行生物信息学分析,筛选了AKTIP基因部分cDNA序列,并对该序列设计了两对特异性引物P1、P2和P3、P4(表 1),以半滑舌鳎总cDNA为模板,进行PCR扩增,扩增产物用1%的琼脂糖凝胶电泳检测,纯化连接转化后,对产生的阳性克隆进行测序。在克隆获得的序列3′端设计RACE(Rapid-Amplification of cDNA Ends)引物P5(表1)和巢式特异性引物P6(表1),使用Clontech公司的SMARTTerTMRACE cDNA Amplification Kit 试剂盒获得该基因的3′端序列,将克隆的序列进行拼接获得AKTIP基因的cDNA全长序列。
1.5 生物信息学分析
对克隆获得的cDNA进行BLASTX(http:// www.ncbi.nlm.nih.gov)比对,利用ORF Finder软件(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)预测该基因的ORF,利用Expasy软件(http://www.expasy.org/)预测其氨基酸序列,蛋白的理化特性预测使用Protparam(http://au.expasy.org/tools/protparam.html),采用Clustal X软件对不同物种AKTIP 基因推导的氨基酸序列进行多序列比对分析,利用MEGA 5.1构建进化树。
1.6 半滑舌鳎AKTIP基因实时定量分析

表 1 PCR扩增所用的特异性引物Tab.1 Specific primers used in this study
采用实时定量PCR方法检测AKTIP基因的表达特征。基于AKTIP保守区设计定量特异引物P7、P8(表 1),用半滑舌鳎18S rRNA基因作为内参(表1)。在ABI PRISM 7500实时定量扩增仪上进行PCR反应,每个样品设置3个生物学重复。健康组织中是相对肝组织做的相对定量分析,采用2-△△Ct法计算,Ct值取3个平行的平均值。使用SPSS 17.0软件对实时定量PCR检测结果的数据进行单因素方差分析和Duncan多重比较统计分析,P<0.05为显著差异。
2 结果
2.1 半滑舌鳎AKTIP基因克隆及其序列特征
半滑舌鳎AKTIP基因经过几轮PCR扩增获得,将两对特异性引物P1、P2和P3、P4扩增克隆出的两段序列和RACE获得的序列经过拼接,获得AKTIP基因完整的cDNA序列,全长为1224 bp,其中包含5′-UTR为116 bp,3′-UTR为117 bp和完整的开放阅读框(ORF)序列891 bp,GC含量是47.7%,共编码296个氨基酸。预测的AKTIP蛋白质等电点(PI)为9.12,分子量是33.74 kD。
2.2 半滑舌鳎AKTIP系统进化分析
通过BLAST比对,发现半滑舌鳎AKTIP氨基酸序列与大西洋鲑(Salmo salar)(NP_00113339 4.1)、斑马鱼(Danio rerio)(NP_956399.1)、非洲爪蟾(Xenopus laevis)(NP_001086693.1)、红原鸡(Gallus gallus)(NP_001005838.1)、美洲鹫(Cathartes aura)(KFP53204.1)、大鼠(Rattus norvegicus)(NP_001011926.1)、小鼠(Mus musculus)(NP_00 1289197.1)、恒河猴(Macaca mulatta)(AFJ71 159.1)、人(Homo sapiens)(NP_001012398.1)的 AKTIP氨基酸序列相似性分别为89%、88%、77%、77%、77%、76%、76%、76%和75%。利用MEGA5.1软件对上述各个物种的AKTIP氨基酸序列进行比对构建了系统进化树(图 1)。结果表明,AKTIP被分为两个大的分支,半滑舌鳎与大西洋鲑(NP_001133394.1)、斑马鱼(NP_956399.1)AKTIP聚为一支,两栖类非洲爪蟾(NP_001086693.1)与红原鸡(NP_001005838.1)、大鼠(NP_0010 11926.1)和人(NP_001012398.1)等的AKTIP聚为一支,但哺乳动物、鸟类和两栖类又按亲缘关系远近都聚成相应的亚类。
2.3 AKTIP基因在半滑舌鳎健康组织表达特异性分析
采用实时定量PCR方法分析了AKTIP基因在半滑舌鳎不同健康组织中的表达特征(图 2)。结果显示:AKTIP基因在健康半滑舌鳎的后肾、脑、胃、脾、头肾、心脏、鳃、血、肌肉、皮肤、肝、肠、卵巢组织中均有表达,表达量由低到高,AKTIP的相对表达量在卵巢组织(17.01)中最高,其次是肠(1.24)和肝(1.02),在后肾和脑中的表达量低(后肾0.21、脑0.24)。AKTIP在不同组织中表达存在组织差异性。
2.4 AKTIP在半滑舌鳎感染鳗弧菌后免疫组织中的表达特征
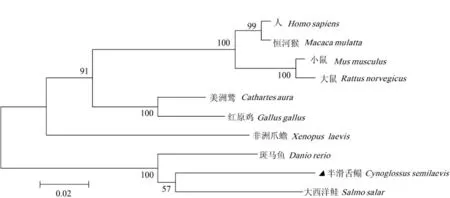
图 1 AKTIP基因家族氨基酸序列的系统进化树
鳗弧菌感染半滑舌鳎后可诱导AKTIP在免疫组织中的表达变化,在PBS对照组和感染组感染0、6h、12h、24h、48h、72h后,AKTIP在肝脏、肠、头肾、脾脏、血液和鳃6种组织中的表达水平如图 3所示。AKTIP在肠中的表达量为6h最低,随后迅速升高,48h是0的1.5倍,总体趋势为先降低又升高再降低; 在脾中表达峰值出现在鳗弧菌感染后48h,是初始表达量的1.77倍,其总体表达趋势为随着感染时间表达量先降低又升高随后下降; 在鳃中的表达变化是6h降到最低,然后一直升高,72h达到最高,其表达量是初始值的2.77倍; 在血中表达量是随着感染时间表达量表现为先下降后升高,在24h出现最高值,是0表达量的1.3倍,然后又降低;在头肾中也是随着感染时间延长表达量先降低后升高,12h表达极低然后迅速升高,在24h出现峰值后又降低,24h表达量是初始值的2.24倍; 肝中AKTIP在12h的表达量极低,最后在72h达到峰值,其表达量是初始表达量的5.33倍。以上数据表明,鳗弧菌感染半滑舌鳎后,可诱导AKTIP在肠、脾、血、鳃、肝脏和头肾中表达上调。
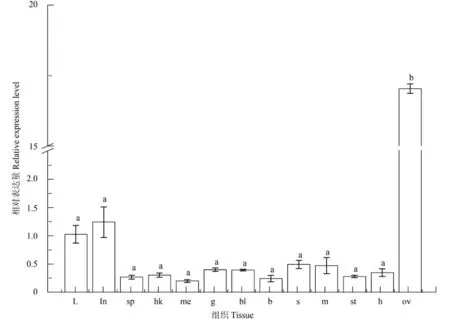
图 2 AKTIP基因在半滑舌鳎健康组织中的实时定量表达
2.5 AKTIP在半滑舌鳎外周血淋巴细胞中的表达
病原模拟物LPS、poly I:C、PGN、WGP分别刺激半滑舌鳎外周血淋巴细胞后,AKTIP基因的实时定量表达结果如图 4所示,LPS对其诱导的表达趋势为:先是持续下降,在12h稍有升高,然后再下降,24h的表达量是0 的0.28倍; 在PGN的诱导下表现2h先降低,其表达量是0的0.71倍,然后有缓慢升高,但总体是下降趋势; Poly I:C的诱导表达表现为先略有升高,2h的表达量与0相当,然后持续下降,24h 的表达量是0的0.40倍; WGP的诱导表达趋势为:先降低,从6h后趋于平缓,6h的表达量是0的0.59倍。总体上,四种病原模拟物LPS、poly I:C、PGN、WGP对AKTIP在半滑舌鳎淋巴细胞中的诱导表达趋势非常相似,都呈下调表达。
3 讨论
AKTIP是PI3K/PDK1/Akt信号通路的支架蛋白,AKTIP可直接或通过与PDK1的相互作用间接作用于蛋白激酶B(PKB/Akt),通过增强Akt的磷酸化调节Akt的活性,从而参与PI3K/PDK1/Akt信号通路的调控[4],AKTIP作为Akt活性的调控蛋白,在PI3K/PDK1/Akt信号通路的调控中起关键性作用;有研究表明AKTIP也参与动物的多种免疫防御过程[5],通过磷酸化Akt活化IkB激酶(IKKα),使NF-κB从细胞质中释放出来进入细胞核内,实现对下游基因的调控,从而参与调节大量关键的生理过程,如细胞增殖[9]、凋亡[10]、固有免疫和适应性免疫[11,12]。可见AKTIP通过调节Akt活性参与免疫防御等多种生物学过程。但是到目前为止,AKTIP的确切作用机制还没有被阐明。
本研究首先克隆获得半滑舌鳎AKTIP的cDNA全长,并利用实时定量PCR检测AKTIP在半滑舌鳎健康组织、感染后的各个组织和感染后淋巴细胞中的表达谱,以期为探讨AKTIP在半滑舌鳎机体中的免疫机制提供理论依据。研究结果表明AKTIP在半滑舌鳎各个健康组织中均有表达,在卵巢中的表达最高,在肝和肠中的表达相对其他组织也较高,推测其可能在免疫调控以及免疫细胞的发育中发挥及其重要的作用。鳗弧菌感染半滑舌鳎72h内,AKTIP在肝、肠、鳃、头肾、血和脾组织中均出现上调表达的情况,说明AKTIP在病原入侵后参与了机体的免疫应答,半滑舌鳎被鳗弧菌感染后为了维持细胞活性,可能大量合成AKTIP基因抵抗细胞凋亡; 或者磷酸化Akt激活NF-κB的信号转导途径,参与免疫反应和炎症反应各阶段许多分子的调控[13],这与Nakamura 等[14]研究结果相一致;LPS、PGN、poly I:C和WGP感染半滑舌鳎外周血淋巴细胞均诱导AKTIP下调表达,但PGN刺激后,AKTIP基因的变化幅度不大,poly I:C和WGP刺激引起的变化相当,LPS诱导AKTIP基因表达变化倍数最大,说明外周血淋巴细胞AKTIP基因对不同的免疫替代物的免疫应答具有特异性。
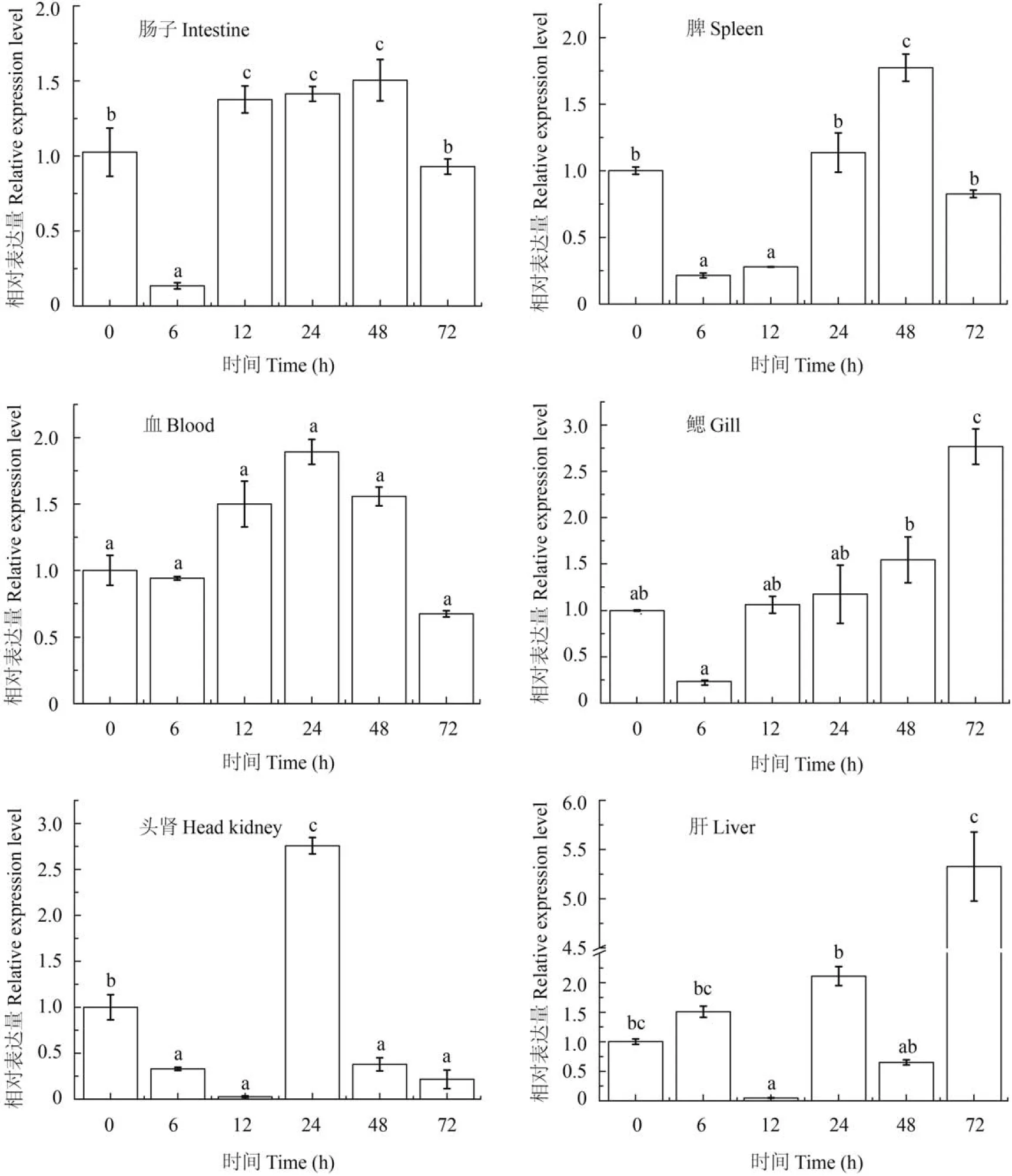
图 3 鳗弧菌感染半滑舌鳎后AKTIP在肠、脾、血、鳃、头肾和肝脏不同免疫组织不同时间点的相对表达水平
AKTIP在半滑舌鳎各个组织均有表达,病原以及革兰氏阳性菌、革兰氏阴性菌和真菌等模拟物刺激后,其在免疫组织和免疫细胞中均出现差异表达,说明AKTIP在半滑舌鳎机体免疫中具有一定的重要作用。综上所述,AKTIP参与了病原感染半滑舌鳎后机体免疫相关基因的调节,本研究为探讨AKTIP在半滑舌鳎机体内重要的免疫作用机制奠定了理论基础。但总体来说在病原和模拟物处理后AKTIP基因的表达变化并不是很显著,推测AKTIP基因在半滑舌鳎中只是间接参与了免疫应答反应,其在免疫中的作用机制有待进一步探讨研究。

图 4 AKTIP在PGN、LPS、poly I:C和WGP刺激半滑舌鳎淋巴细胞中的表达
参 考 文 献:
[1]Lesche R,Peetz A,van der Hoeven F,et al.Ftl,a novel gene related to ubiquitin-conjugating enzymes,is deleted in the Fused toes mouse mutation[J].Mammalian Genome,1997,8(12):879—883
[2]Zhen S,Zhou J F,Lu L,et al.Cloning and expression pattern and over-expression analysis of Zebrafish akt3 gene[J].Acta Hydrobiologica Sinica,2011,35(5):717—726[谌爽,周建峰,卢玲,等.斑马鱼akt3基因的克隆及其表达图谱与过表达分析.水生生物学报,2011,35(5):717—726]
[3]Remy I,Michnick S W.Regulation of apoptosis by the Ft1 protein,a new modulator of protein kinase B/Akt[J].Molecular and Cellular Biology,2004,24(4):1493—1504
[4]Woodard J,Sassano A,Hay N,et al.Statin-dependent suppression of the Akt/mammalian target of rapamycin signaling cascade and programmed cell death 4 up-regulation in renal cell carcinoma[J].Clinical Cancer Research,2008,14(14):4640—4649
[5]Jiang H,Zhang X J.PKB/Akt:A protein kinase that has a variety of functions[J].Life Science,2004,16(3):148—153[姜华,张学军.PKB/Akt:一个具有多种功能的蛋白激酶.生命科学,2004,16(3):148—153]
[6]Romashkova J A,Makarov S S.NF-κB is a target of AKT in anti-apoptotic PDGF signaling[J].Nature,1999,401(6748):86—90
[7]Sha Z X,Wang Q L,Liu Y,et al.Identification and expression analysis of goose-type lysozyme in half-smooth tongue sole(Cynoglossus semilaevis)[J].Fish & Shellfish Immunology,2012,32(5):914—921
[8]Sha Z,Gong G,Wang S,et al.Identification and characterization of Cynoglossus semilaevis microRNA response to Vibrio anguillarum infection through high-throughput sequencing[J].Developmental & Comparative Immunology,2014,44(1):59—69
[9]Fatrai S,Elghazi L,Balcazar N,et al.Akt induces β-cell proliferation by regulating cyclin D1,cyclin D2,and p21levels and cyclin-dependent kinase-4 activity[J].Diabetes,2006,55(2):318—325
[10]Datta S R,Dudek H,Tao X,et al.Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery[J].Cell,1997,91(2):231—241
[11]Nicholson K M,Anderson N G.The protein kinase B/Akt signalling pathway in human malignancy[J].Cellular signalling,2002,14(5):381—395
[12]Yamada T,Takeuchi S,Fujita N,et al.Akt kinase-interacting protein1,a novel therapeutic target for lung cancer with EGFR-activating and gatekeeper mutations[J].Oncogene,2013,32(37):4427—4435
[13]Chandrasekar B,Mummidi S,Perla R,et al.Fractalkine(CX3CL1)stimulated by nuclear factor kappaB(NF-kappaB)-dependent inflammatory signals induces aortic smooth muscle cell proliferation through an autocrine pathway[J].Biochemical Journal,2003,373:547—558
[14]Nakamura A,Naito M,Tsuruo T,et al.Freud-1/Aki1,a novel PDK1-interacting protein,functions as a scaffold to activate the PDK1/Akt pathway in epidermal growth factor signaling[J].Molecular and Cellular Biology,2008,28(19):5996—6009
AKT-INTERACTING PROTEIN GENE CLONING AND ITS EXPRESSION PROFILE IN RESPONSE TO PATHOGEN INFECTION IN HALF SMOOTH TONGUE SOLE(CYNOGLOSSUS SEMILAEVIS)
SUN Lu-Ming1,2,YU Meng-Jun1,2,CHEN Ya-Dong2,3,CHEN Xue-Jie2,4,LIU Yang1,QIU Xue-Mei1and SHA Zhen-Xia2,3
(1.College of Fishers and Life Sciences,Dalian Ocean University,Dalian 116023,China; 2.Key Laboratory for Sustainable Development of Marine Fisheries,Ministry of Agriculture,Yellow Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Qingdao 266071,China; 3.Laboratory for Marine Fisheries and Aquaculture,Qingdao National Laboratory for Marine Science and Technology,Qingdao 266200,China; 4.College of Fishers and Life Sciences,Shanghai Ocean University,Shanghai 201306,China)
Abstract:AKT-interacting protien(AKTIP)is a kind of membrane protein,involvimg in the regulation of PI3K/PDK1/Akt pathway.AKTIP gene is still unknown in fish.In the present study,AKTIP gene of Cynoglossus semilaevis was first cloned and full length of cDNA was 1224 bp in size,including 5′-untranslated region(UTR)of 116 bp,3′-UTR of 117 bp and a complete open reading frame(ORF)of 891 bp,encoding 296 amino acids.Theoretical isoelectric point(PI)of predicted protein was 9.12 and molecular weight was 33.74 kD.Homologous comparison showed that the amino acids sequence of AKTIP in C.semilaevis had a high identity with those of other species.The AKTIP gene was expressed in all tested tissues in the health C.semilaevis and the highest expression was in the ovary.In order to investigate the expression patterns of the AKTIP gene in immune response,the specific expression of AKTIP was performed after Vibrio anguillarum infection and in peripheral blood lymphocytes with different pathogens stimulated.The results showed that the expression of AKTIP gene was up-regulated in the liver,gill,blood,intestinal,head kidney and spleen after V.anguillarum infection.While the AKTIP gene expression in peripheral blood lymphocytes was down-regulated after PGN,LPS,WGP and poly I:C stimulation.The research revealed that AKTIP gene involved in C.semilaevis immune response.These results provide the evidence to explore AKTIP defense mechanism and to control C.semilaevis disease in the further study.
Key words:Cynoglossus semilaevis; AKT-interacting protein; Gene clone; Immune response; Gene expression
中图分类号:Q522
文献标识码:A
文章编号:1000-3207(2016)03-0467-07
doi:10.7541/2016.62
收稿日期:2015-09-17;
修订日期:2016-01-19
基金项目:国家“863”高技术研究发展计划(2012AA10A401-4); 国家自然科学基金(31172439)资助[Supported by the National High Technology Research and Development Program of China(863 Program)(2012AA10A401-4); the National Natural Science Foundation of China(31172439)]
作者简介:孙璐明(1990—),女,山东临沂人; 硕士研究生; 主要从事生物化学与分子生物学研究。E-mail:sunluming9966@163.com
通信作者:沙珍霞,女,研究员; 主要从事鱼类分子免疫学、基因组学和细胞生物学研究。E-mail:shazx@ysfri.ac.cn
