皖西多枝尖山区不同生活型植物种-面积关系研究
2016-06-20王志高吴甘霖段仁燕张中信
王志高, 吴甘霖, 段仁燕, 张中信
(安庆师范学院 生命科学学院, 皖西南生物多样性研究与生态保护安徽省重点实验室, 安徽 安庆 246133)
皖西多枝尖山区不同生活型植物种-面积关系研究
王志高*, 吴甘霖, 段仁燕, 张中信
(安庆师范学院 生命科学学院, 皖西南生物多样性研究与生态保护安徽省重点实验室, 安徽 安庆 246133)
摘要:根据皖西大别山多枝尖山区的植物群落调查数据,采用对数函数、幂函数和Logistic 3种模型分别拟合了乔木、灌木和草本植物的种-面积关系。结果表明,大别山多枝尖山区不同生活型植物种-面积关系的形式存在差异,其中乔木植物以Logistic模型拟合效果最好,而灌木植物和草本植物的最优模型均为对数函数模型。生境异质性、扩散限制和区域生态过程是乔木植物种-面积关系的主要影响因素,而灌木和草本植物的种-面积关系受到生长习性以及上层群落结构和生境异质性的共同作用。因而,对种-面积关系的研究需要区分物种生活型。
关键词:多枝尖; 生物型; 种-面积关系; 物种多样性
种-面积关系(species-area relationship,SAR)是对物种数目与取样面积之间关系的定量化描述,是生态学中的经典范式[1-4]。前期的大量研究表明,不同群落的种-面积曲线形式存在差异。Logistic模型、对数函数模型和幂函数模型是种-面积关系中最常见的3种形式[5-9]。其中,幂函数模型受到广泛关注,且该函数中比例因子z值的生态学意义引起了众多生态学家的兴趣。z值具有尺度依赖性,不同尺度上z值存在差异,但其生态学意义仍不明确[10-12]。种-面积关系受到物种空间分布格局的影响,物种多度分布模式和生境异质性影响种-面积关系的类型。当物种多度分布为窄对数正态分布时,种-面积关系呈指数形式;物种多度呈宽对数正态分布时,种面积关系呈幂函数分布[11-12];在局部小尺度上随着取样面积的增加,生境异质性的存在会导致物种数目增长较快[5-6]。群落中性理论认为在区域尺度上,由于受物种库大小的影响,种-面积关系呈Logistic分布[13-14]。对种-面积关系的研究有助于了解群落物种多样性的形成机制,同时在生物多样性管理、物种数目估计以及取样面积的确定等方面具有非常重要的应用价值。
不同生活型植物由于生长方式、分布格局以及扩散方式的差异,其种-面积关系可能存在不同。基于此,本文以大别山多枝尖山区植物群落调查资料为依据,分别分析了乔木、灌木和草本植物的种-面积关系,以期为进一步研究提供理论借鉴。
1研究方法
1.1研究区域概况
大别山多枝尖山区位于安徽省岳西县境内, 地理坐标为东经116°03′~ 116°09′、北纬31°00′~31°11′。多枝尖山区最高海拔1 721.5 m,是大别山的第三高峰。该区域在20世纪70—80年代遭到大规模砍伐,但自1994年被列为鹞落坪国家级自然保护区的核心区之后,次生植被得到了很好的保护和恢复。该区动植物资源丰富,典型植被类型保存较完整。保护区年降水量为1 400~2 000 mm,年日照时数为1 580~1 950 h,太阳年辐射总量为4 200~4 650 MJ/m2,多年平均蒸发量为1 444 mm,年均相对湿度在80%以上。鹞落坪自然保护区土壤具有明显的淀积黏化、生物累积、弱富铝化、水耕熟化等过程,形成了黄棕壤、山地棕壤、山区草甸土和水稻土等不同的土壤类型[13-20]。
1.2取样方法
2009年8月,采用样方法对多枝尖山区植被进行了群落调查。在1 200~1 700 m海拔范围内,在本区典型分布的黄山松林(Pinustaiwanensis)、华山松林(Pinusarmandii)、金钱松林(Pseudolarixamabilis)、枹栎林(Quercusserrata)和黄山栎林(Quercusstewardii)中随机设置了25个20 m×20 m的乔木样方,每个乔木样方中设置5个2 m×2 m的灌木样方和5个1 m×1 m的草本样方。对乔木进行每木调查(胸径1 cm以上),记录种名、胸径和坐标;记录灌木层和草本层植物的种名、株数、高度、盖度,同时记录各乔木样方的坡度、坡向和海拔等环境数据。详细的样方特征参见文献[19]。1.3种-关系面积的构建及模型选择
本研究采用组合样方的方式进行取样面积的累加。即对面积相同的样方进行随机组合,形成不同面积的样方组合系列,以不同样方组合中出现的物种数构建种-面积关系,面积由所包含的样地数目决定[21]。共进行1 000次样方随机组合(1 000次重复)。
分别采用以下3个模型拟合乔木、灌木和草本植物的种-面积关系:
1)Arrhennius的幂函数模型为
S=cAz。
2)对数函数模型为
S=zlnA+C。
3)Logistic模型为

上述模型中:S为物种数目,A为取样面积,C为常数,a、c和z为模型参数。
1.4模型拟合优度检验

式中:n为取样数目,k为模型的参数数量,RSS为残差平方和,TSS为总平方和。
2结果与分析
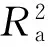
3讨论
大别山多枝尖山区地带性植被是在20世纪70年代遭破坏后恢复形成的天然次生林,目前植被恢复良好,其植被在高海拔区域以及山脊上为黄山松纯林和小面积的黄山栎林,中海拔地带为黄山松和枹栎组成的针阔混交林以及少量人工种植的金钱松和华山松林,乔木层植物种类简单[15-18]。我们前期对该区域的研究表明,多枝尖山区灌木和草本植物的种间联结松散,受到生境异质性和种间相互作用的影响[19-20]。本文对乔木、灌木和草本植物种-面积关系的研究表,乔木植物以Logistic模型拟合效果最好,而灌木植物和草本植物的最优模型均为对数函数模型。
种-面积关系的形式与构建群落结构的生态过程密切相关[6,13]。在相对均质的地段,取样面积的增加意味着包含更多的个体[22]。生态位分化理论认为物种适应于不同的生境,生境异质性的增加会提高群落物种多样性[6]。取样面积的增加伴随着生境异质性的增加,因而物种数量也逐步增加[5]。在局域尺度上,随着取样面积的进一步增大,生境异质性不会无限增加,因此种-面积关系呈幂函数分布模式[23]; 在区域尺度上还有可能包含一些只发生在大尺度的进化或生态过程[24],物种数目将会逐渐接近于区域种库的限制。由于扩散限制的存在,植物群落中的物种大多数呈聚集分布,而生境异质性也会导致物种的聚集分布,因此生境异质性与扩散限制的影响难以区分,都会对种-面积关系的形式产生影响[25-27]。本研究中乔木植物的种-面积关系呈Logistic分布,进一步说明了群落物种多样性维持机制具有尺度依赖性特征[10],局域性的生态位分化和随机扩散过程以及区域尺度上的进化过程均影响群落的物种多样性。
不同的生活型植物对环境的适应方式不同,本文中灌木和草本植物的种-面积关系呈对数函数分布,表明随着取样面积的增加,新物种的数目增加速度较快。林下生境受到生物因子(上层群落结构)与非生物因子(如地形和土壤)的共同影响[19-20]。相同的地形条件下,上层植物群落结构的差异也会导致林下生境的不同,因此随着取样面积的增加,灌木和草本植物新出现物种的概率大于乔木植物。另外,前人的研究表明林下灌木和草本植物多成斑块状分布[28],这些因素都会影响种-面积关系的形式。灌木和草本植物的物种数目随着取样面积的增加,新物种增加的速度比幂函数模型快[5,25],而用对数函数模型描述较为合适。因此,灌木和草本植物的种-面积关系受到生境异质性的影响外,还与其生长习性有关。种-面积关系模式常用来估计群落物种数目、确定取样面积和探讨群落物种多样性形成机制。本研究表明,模拟种-面积关系时应考虑物种的生活型,这仍需要更多的研究结果来证实。
4结论
多枝尖山区乔木植物的种-面积关系呈Logistic分布,灌木和草本植物的种-面积关系符合对数函数分布。我们建议,在使用种-面积关系模型探讨群落物种多样性形成机制时,需要对不同生活型植物分别进行模拟。
参考文献:
[1] ARRHENIUS O. Species and area[J].Journal of Ecology, 1921, 9(1):95-99.
[2] CAIN S. The species area curve[J]. The American Naturalist, 1938, 19:573-581.
[3] GLEASON H. On the relation between species and area[J]. Ecology, 1922, 3(2): 158-162.
[4] GLEASON H .Species and area[J]. Ecology, 1925, 6(1):66-74.
[5] HE F, LEGENDRE P. On species area relations[J]. The American Naturalist, 1996, 148:719-737.
[6] HE F, LEGENDRE P. Species diversity patterns derived from species area models[J].Ecology, 1996, 83(5):1185-1198.
[7] SCHOEREDER J, GALBIATI C, RIBAS C, et al. Should we use proportional sampling for species area studies[J]. Journal of Biogeography, 2004, 31(8): 1219-1226.
[8] TJ∅RVE E. Shapes and functions of species area curves (Ⅱ): a review of new models and parameterizations[J].Journal of Biogeography, 2009, 36(8): 1435-1445.
[9] HENSTRIDGE J. Species area functions revisited[J]. Journal of Biogeography, 2009, 36(10): 1994-2004.
[10] CRAWLEY M, HARRAL J E. Scale dependence in plant biodiversity[J]. Science, 2001,291(5505): 864-868.
[11] KEELEY J, FOTHERINGHAM C J. Species-area relationships in Mediterranean-climate plant communities[J]. Journal of Biogeography, 2003,30(11): 1629-1657.
[12] KEELEY J, FOTHERINGHAM C J. Plot shape effects on plant species diversity measurements[J]. Journal of Vegetation Science, 2005, 16(2): 249-256.
[13] HUBBELL S. The unified neutral theory of biodiversity and biogeography (MPB-32)[M]. Princeton: Princeton University Press, 2001.
[14] BELL G. Neutral macroecology[J]. Science, 2001, 293(5539): 2413-2418.
[15] 谢中稳,吴国芳.安徽大别山多枝尖山区植物区系的研究[J].华东师范大学学报(自然科学版),1993(1):102-110.
[16] 谢中稳,吴国芳.安徽省鹞落坪自然保护区植被类型及其分布[J].华东师范大学学报(自然科学版), 1995 (3): 93-102.
[17] 高林,徐如松.鹞落坪国家级自然保护区植被类型的初步探讨[J].生物学杂志,2002,19(6):32-42.
[18] 单人驊, 刘方勋.安徽大别山的植被及其地理分布纪要[J].植物生态学及地植物学丛刊, 1964,2(1):93-102.
[19] 吴甘霖,王志高,段仁燕,等.安徽大别山多枝尖山区植物物种多样性[J].林业科学,2010,46(6): 128-132.
[20] 王志高,张中信,段仁燕,等. 大别山多枝尖山区林下植物种间联结性研究[J].生态环境学报,2010,19(9): 2066-2071.
[21] 唐志尧, 乔秀娟, 方精云.生物群落的种-面积关系[J], 生物多样性,2009, 17 (6): 549-559.
[22] DURRETT R, LEVIN S. Spatial models for species-area curves[J].Journal of Theoretical Biology, 1996, 179(2): 119-127.
[23] MACARTHUR R, WILSON E. An equilibrium theory of insular zoogeography[J], Evoluotion, 1963, 17:373-387.
[24] LOSOS J, SCHLUTER D. Analysis of an evolutionary species area relationship[J]. Nature, 2000, 408(6814):847-850.
[25] TJ∅RVE E, KATHLEEN M, TJ∅RVE C. The species area relationship, self-similarity, and the true meaning of thez-value[J]. Ecology, 2008, 89(12):3528-3533.
[26] ROSINDELL J, CORNELL S J. Species area curves, neutral models, and long-distance dispersal[J]. Ecology, 2009, 90(7): 1743-1750.
[27] TURNER W, TJ∅RVE E.Scale-dependence in species-area relationships [J]. Ecography, 2005, 28: 721-730.
[28] GRACIA M, MONTAN F, PIQUÉ J, et al. Overstory structure and topographic gradients determining diversity and abundance of understory shrub species in temperate forests in central pyrenees(NE Spain)[J]. Forest Ecology and Management, 2007, 242: 391-397.
〔责任编辑王勇〕
Relationships between different life form species and its possessive area in Duozhijian, Dabieshan mountain, west Anhui pronvince
WANG Zhigao*, WU Ganlin, DUAN Renyan, ZHANG Zhongxin
(School of Life Science, Anqing Normal College, the Province Key Laboratory of the Biodiversity and Ecological Conservation in Southwest Anhui, Anqing 246133, Anhui, China)
Abstract:Using the data from Duozhijian, Dabieshan Mountain in west Anhui province, species-area relationships of arbor, shrub and herb species were modeled by logarithmic, power and logistic model, respectively. The results showed that the canopy species fitted logistic model best, while the shrub and herb species fitted logarithmic model. Different types of species area relationship curves indicated the different assemblies of arbor, shrub and herb species. Habitat heterogeneity,dispersal limitation and regional ecological process contributes to the species-area relationship of arbor species, while the species-area relationships of shrub and herb species are influenced by the ecological habits and the jointly effects of habitat heterogeneity and upper community structure. Different life forms and comprehensively analyses were suggested to explore the mechanism of species diversity maintaining using species-area relationships.Keywords: Duozhijian; life form; species-area relationship; species diversity
文章编号:1672-4291(2016)03-0091-04
doi:10.15983/j.cnki.jsnu.2016.03.336
收稿日期:2015-11-30
基金项目:中国科学院战略先导科技专项子专题(XDA0505020401-2);安徽省高校省级自然科学基金(KJ2010B087);安徽省自然科学基金(1408085MC64);安庆师范学院博士科研启动费。
*通信作者:王志高,男,副教授,博士。E-mail:zg_wang@126.com
中图分类号:Q948.1
文献标志码:A
