西南喀斯特地区不同龄级野生板栗光合日动态差异
2016-06-14王健宋淑珍杨青孟庆杰霍如雪
王健+宋淑珍+杨青+孟庆杰+霍如雪

摘要:为了研究西南喀斯特地区野生板栗不同龄级的光合特性差异,利用英国Lcpro+光合测定仪对西南喀斯特地区的3龄苗和6龄苗野生板栗的光合日变化特征进行了测定。结果表明:3龄苗野生板栗的日均净光合速率为7.3 μmol/(m2·s)(以CO2计)、蒸腾速率为4.26 mmol/(m2·s)(以H2O计)、水分利用效率1.90 μmol/mmol;6龄苗野生板栗的日均净光合速率为6.61 μmol/(m2·s)(以CO2计)、蒸腾速率为3.47 mmol/(m2·s)(以H2O计)、水分利用效率为2.02 μmol/mmol。3龄苗由于苗种生理发育尚未完全,光合作用受外界环境影响较为敏感,尤其是光合有效辐射是3龄苗板栗净光合速率的主要限制因子。6龄苗的净光合速率则与胞间CO2浓度密切相关,胞间CO2浓度作为植物叶片光合作用的物质来源,对挂果期来说十分重要。3龄苗的蒸腾速率与大气温度密切相关,说明野生板栗幼苗期的蒸腾速率严格受到大气温度的控制。6龄苗的蒸腾速率首先受到大气温度的严格制约,其次受到光照度的影响,比3龄苗更符合植物进化演替规律,也更加趋向植物体的成熟发展。
关键词:石漠化;野生板栗;净光合速率;日动态
中图分类号: S664.201
文献标志码: A
文章编号:1002-1302(2016)04-0237-03
植物体中超过90%的干物质都来自叶片的光合同化作用,研究植物叶片的光合作用特性,对制定合理的栽培措施、提高产量和品质均有十分重要的意义[1]。植物的叶片结构特征、光合特性与其所处的环境密切相关,植物会根据生境条件改变表现出不同的适应机制,均可以通过植物光合特性、叶片性状及它们之间的关系反映出来[2-7]。西南喀斯特生态系统是受喀斯特环境制约的生态系统[8],受地下、地上2层地质地貌环境的影响,喀斯特环境地表极易发生干旱,该地区植被多表现出旱生、石生和嗜钙等适应机制。为此,研究该区域潜在的野生经济林木要特别考虑环境因素对植物光合特性的影响。
板栗(Castanea mollissima)为壳斗科栗属植物,是我国重要的经济林树种,是世界著名干果,也是农区重要的果品产出,栽培及经营历史悠久,分布非常广泛,营养价值与经济价值较高,深受国内外广大消费者的欢迎和产区农民群众喜爱;同时,板栗的木材还是上等家具与工艺品的原材料,其开发利用价值已引起国内外的广泛关注[9]。
同一种植物,因其树龄、品种、种源地的差异会表现出光合特性差异。目前,国内外对野生板栗的研究多集中于板栗的微观生理如真菌、遗传等,以及品质与资源开发等[10-18],对不同龄级的光合差异研究尚属空白。本研究主要分析了西南喀斯特地区不同龄级野生板栗的光合生理日变化特征,以期为我国喀斯特地区野生板栗资源的开发与科学研究提供一定依据。
1 材料与方法
1.1 研究区自然概况
试验地研究位于贵州野生板栗广泛分布的贵州省毕节市撒拉溪镇喀斯特石漠化山区(105°02′~105°08′E,27°11′~27°16′N),属于长江流域乌江水系六冲河支流上游,为喀斯特高原山地轻中度石漠化分布区,高中山地貌基岩裸露严重,土层浅薄,水土流失严重。该地区气候属亚热带季风气候,年平均日照时间为1 360 h,年平均降水量为984.4 mm。原始植被以板栗、火棘[Pyracantha fortuneana (Maxim.) L.]为主。
1.2 试验材料
选取3、6龄苗的野生板栗作为研究材料。
1.3 试验方法
2014年4月,于晴天进行光合测定的田间试验,在08:00—18:00时段内,取中间时段,每隔2 h测定1次,共测6次;2种试验材料随机选取3株,每株选取植株3张向阳叶片进行试验,取均值。光合测定仪器采用英国ADC Bioscientific公司生产的Lcpro+光合仪,测定叶室采用红蓝光源叶室,测定叶片瞬时净光合速率[Pn,μmol/(m2·s)(以CO2计)]、蒸腾速率[Tr,mmol/(m2·s)(以H2O计)]、CO2气孔导度[Gs,mol/(m2·s),以H2O计]、细胞间CO2浓度(Ci,μmol/mol)、大气温度(T,℃)、光合有效辐射[PAR,μmol/(m2·s)]等指标。水分利用效率 WUE=Pn/Tr(μmol CO2/mmol H2O),主要反映植物体吸收CO2和蒸腾水分消耗的关系。利用SPSS 20软件进行数据统计分析,Excel 2007软件进行绘图。
2 结果与分析
2.1 大气参数日的变化特征
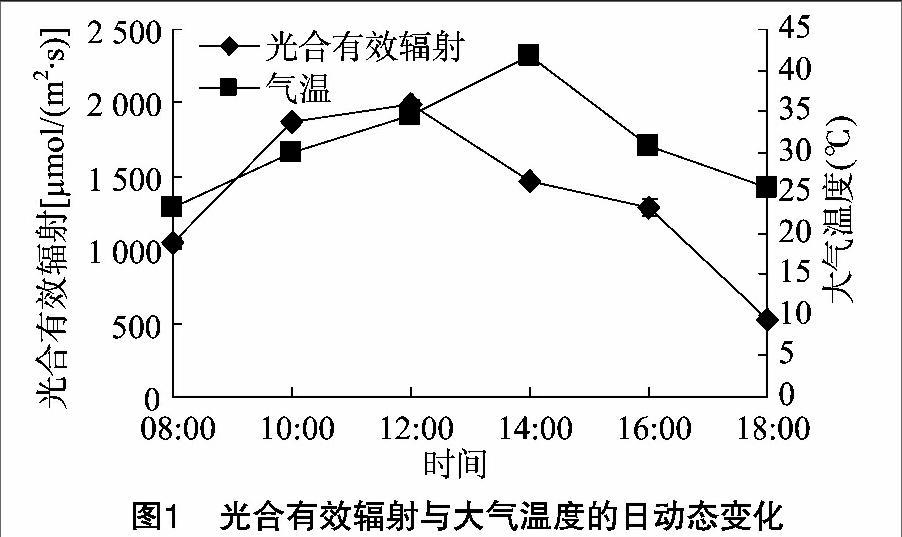
由图1可见,试验日光合有效辐射PAR在08:00为1 044.17 μmol/(m2·s),随后逐渐升高,在12:00为1 998.08 μmol/(m2·s);在18:00为1 d中最低值,为525.92 μmol/(m2·s),日均值(1 365.97±18.34) μmol/(m2·s)。大气温度在08:00为23.15 ℃,14:00出现最高值41.82 ℃,18:00降低到25.68 ℃,日均值为(30.99±0.02) ℃。
2.2 净光合速率的日变化特征
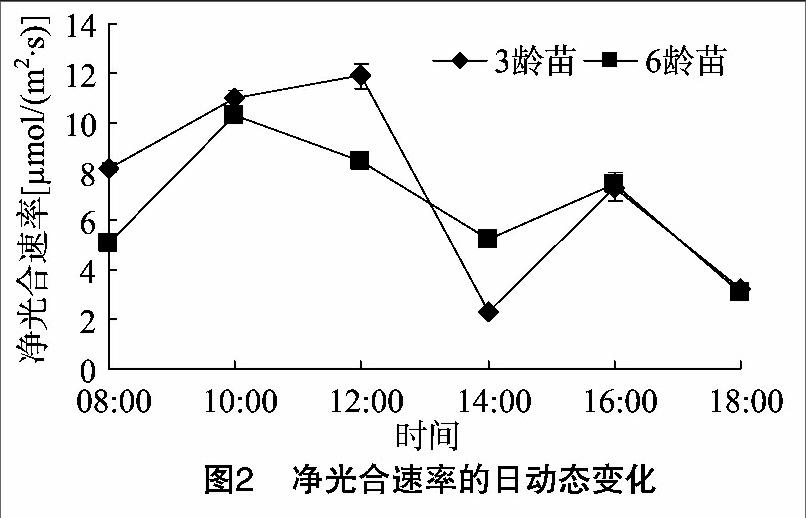
3、6龄苗的净光合速率日均值分别为(7.31±0.27)、(6.61±0.11) μmol/(m2·s),3龄苗的日均净光合速率要高于6龄苗。由图2可见,3龄苗与6龄苗的净光合速率日变化曲线变化趋势较为相似,均为“双峰”型曲线。3龄苗的净光合速率从08:00开始增长,在12:00达到峰值,在14:00出现光合“午休”谷值,在16:00略有上升,形成次峰值;光合“午休”现象推迟到14:00,说明3龄苗对强光照敏感反应较为迟缓。6龄苗的净光合速率曲线日变化变化幅度较3龄苗小,在08:00—10:00之间净光合速率上升迅速,10:00出现峰值,在14:00出现次峰值,出现光合“午休”谷值低于6龄苗,光合“午休”现象不如3龄苗明显。
2.3 蒸腾速率的日变化特征
3、6龄苗野生板栗的蒸腾速率日均值分别为(4.26±0.13)、(3.47±0.02) mmol/(m2·s),3龄苗的日均蒸腾速率同样高于6龄苗。从图3可见,3、6龄苗的蒸腾速率日变化曲线趋势大致相同,均为单峰型曲线,在14:00出现最大值,18:00出现最低值;略有区别的是3龄苗的最大峰值要明显高于6龄苗,且14:00以后3龄苗蒸腾作用更为强烈。
2.4 水分利用效率日变化特征
3、6龄苗野生板栗的水分利用效率日均值分别为(1.91±0)、(2.02±0)μmol/mmol。由图4可见,3龄苗与6龄苗的水分利用效率日变化曲线变化趋势接近但也存在一定差异,尤其是08:00—10:00阶段,2者水分利用效率峰值均出现在08:00。龄苗次峰值出现在14:00,6龄苗出现在18:00,谷值均出现在14:00;6龄苗日变化幅度小于3龄苗,但3龄苗在12:00之前则更为平缓。
2.5 气孔导度的日变化特征
由图5可知,3、6龄苗的气孔导度曲线走势一致,变化幅度有所差异,2者均在08:00出现最大值,14:00出现午间低值,18:00出现最低值,8:00、16:00差异较大,日均值分别为(0.14±0.01)、(0.10±0) mol/(m2·s)。
2.6 胞间CO2浓度的日变化特征
3、6龄苗的胞间CO2浓度日均值分别为(245.94±5.84)、(225.17±2.12)μmol/mol。由图6可知,6龄苗野生板栗的胞间CO2浓度曲线变化较为平缓,1 d间差异不明显,3龄苗则幅度变化明显,具有明显的谷值与峰值;两者最大值均出现在08:00,3龄苗次峰值出现在14:00,6龄苗则出现在18:00。
2.7 光合速率、蒸腾速率与各因子的相关性
由表1可知,3龄苗野生板栗的净光合速率与光合有效辐射相关性显著,与其他因子相关性不明显。6龄苗野生板栗的净光合速率与胞间CO2浓度呈显著负相关,与其他因子相关性不明显。3龄苗野生板栗的蒸腾速率与大气温度显著相关,与其他因子相关性不明显。6龄苗野生板栗的蒸腾速率与光合有效辐射显著相关,与大气温度极显著相关,与其他因子相关性不明显。
4 结论与讨论
光合作用、蒸腾作用是植物重要的生命活动,其中光合作用是植物生长的生理基础物质积累来源,蒸腾作用耗水则是树木水分散失的主要途径与吸收地下水的方式,水分利用效率则是评价植物水分消耗、利用能力的主要指标[19]。植物生理的叶片温度高影响了酶的活性,进而影响了植物本身的生化反应和光合速率,同时光照度也影响光合日变化的进程[25]。3龄苗为野生板栗的幼苗期,此时由于苗种生理发育尚未完全,光合作用受外界环境影响较为敏感,尤其是光合有效辐射,是3龄苗板栗净光合速率的主要限制因子。6龄苗为野生板栗的挂果初期,此时植物的净光合速率则与胞间CO2浓度密切相关,胞间CO2浓度作为植物叶片光合作用的物质来源,对挂果期板栗来说十分重要。3龄苗的蒸腾速率与大气温度密切相关,说明野生板栗幼苗期的蒸腾速率严格受到大气温度的控制。6龄苗的蒸腾速率首先受到大气温度的严格制约,其次受到光照度的影响,比3龄苗更符合植物进化演替规律,也更加趋向植物体的成熟发展。
试验结果中3龄苗的净光合速率要高于6龄苗,说明其处于生长期进行光合作用的能力远高于6龄苗的挂果期板栗;同时,3龄苗的蒸腾作用更易受外界影响,同时与其所处时期关系密切,导致其蒸腾速率高于6龄苗。6龄苗的板栗已经进入生产期,逐渐适应了环境变化,水分利用效率高于生长期的3龄苗。
综上所述,野生板栗是西南喀斯特地区的重要野生经济林种,适应范围较大,在南北方均可推广种植,并且能够很好地适应石漠化地区贫瘠浅薄的土壤,并且经过长期监测研究表明生态效益与社会经济效益显著。喀斯特环境由于特殊地质背景,在该地区培育驯化野生经济林时要充分考虑树种的光合生理特性,较高的净光合速率和较低的蒸腾速率充分利用喀斯特稀缺的水分资源,同时产生更多的物质累积,提高潜在的生态与经济效益。总体来说,要研究不同板栗与不同经济林间种提高光合作用效率,探索经济林乔-灌结合,增加地表覆盖度,减少土壤侵蚀。
参考文献:
[1]池永宽,熊康宁,王元素,等. 西南石漠化地区两种经济林木光合日动态特征[J]. 经济林研究,2015,33(1):45-49,55.
[2]Manes F,Seufert G,Vitale M. Ecophysiological studies of Mediterranean plant species at the Castelporziano estate[J]. Atmospheric Environment,1997,31(1):51-60.
[3]Cao B,Dang Q L,Zhang S. Relationship between photosynthesis and leaf nitrogen concentration in ambient and elevated [CO2] in white birch seedlings[J]. Tree Physiology,2007,27(6):891-899.
[4]Midgley G F,Aranibar J N,Mantlana K B,et al. Photosynthetic and gas exchange characteristics of dominant woody plants on a moisture gradient in an African savanna[J]. Global Change Biology,2004,10(3):309-317.
[5]Ueda Y S,Nishihara H,Tomita Y O. Photosynthetic response of Japanese rose species Rosa bracteata and Rosa rugosa to temperature and light[J]. Scientia Horticulturae,2000,84(3/4):365-371.
[6]John R,Dalling J W,Harms K E,et al. Soil nutrients influence spatial distributions of tropical tree species[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(3):864-869.
[7]Guo W H,Bo L,Huang Y M,et al. Effects of different water stresses on eco-physiological characteristics of Hippophae rhamnoides seedlings[J]. Acta Botanica Sinica,2003,45(10):1238-1244.
[8]熊康宁,池永宽.中国南方喀斯特生态系统面临的问题及对策[J]. 生态经济,2015,31(1):23-30.
[9]谭方友,吴德辉,黄 河.贵州省干果生产现状及发展对策[J]. 经济林研究,2009,27(4):126-128.
[10]郭献平,李兴亮,段续伟,等. 板栗野生和芽变雄花序赤霉素合成关键酶基因比较分析[J]. 中国农业大学学报,2012,17(4):91-95.
[11]艾呈祥,沈广宁,张 凯,等. 秦巴山区野板栗居群遗传多样性AFLP分析[J]. 植物遗传资源学报,2011,12(3):408-412.
[12]黄武刚,程丽莉,周志军,等. 板栗野生居群遗传多样性研究[J]. 果树学报,2010,27(2):227-232.
[13]柴迪迪,郭素娟,牛晓丹. 野生板栗根部菌根真菌的分离与回接效应研究[J]. 北方园艺,2010(21):18-21.
[14]黄武刚,程丽莉,周志军,等. 板栗野生居群与栽培品种间叶绿体微卫星遗传差异初探[J]. 林业科学,2009,45(10):62-68.
[15]马玉敏,陈学森,何天明,等. 中国板栗3个野生居群部分表型性状的遗传多样性[J]. 园艺学报,2008,35(12):1717-1726.
[16]吴得荣,王冬梅,刘 敏,等. 野生板栗群落优势树种间竞争研究[J]. 湖南农业科学,2011(7):115-118.
[17]杨 剑,涂炳坤,谢谱清,等. 湖北的主要板栗品种资源及其野生近缘种[J]. 广西植物,2001,21(2):187-190.
[18]邱国金. 野生板栗资源的开发与利用[J]. 江苏农业科学,1993(1):50-51.
[19]邱 权,潘 昕,李吉跃,等. 速生树种尾巨桉和竹柳幼苗的光合特性和根系特征比较[J]. 中南林业科技大学学报,2014,34(1):53-59.
[20]孟丙南,张俊佩,裴 东,等. 不同砧木对核桃光合特性的影响[J]. 经济林研究,2013,31(2):32-37.贾晓梅,崔彬彬,温陟良,等. 冬枣不同环剥宽度枣吊中内源激素含量分析[J]. 江苏农业科学,2016,44(4):240-241.
