稻瘟病菌MoRho2基因功能初步分析
2016-05-30杨帆宋林林谢序泽赵臻周立帅叶文雨鲁国东
杨帆 宋林林 谢序泽 赵臻 周立帅 叶文雨 鲁国东

摘 要 稻瘟病菌(Magnaporthe oryzae)是研究病原真菌生长发育及致病机理的重要模式生物。Rho家族蛋白是一类具有GTP酶活性的GTP结合蛋白,在多种细胞信号转导通路中起着分子开关的作用。通过生物信息学方法分析MoRho2的理化性质、蛋白结构域等信息,并推测其潜在功能。为进一步明确MoRho2蛋白的功能,利用基因敲除技术获得MoRho2基因的敲除突变体。表型分析结果表明,与野生型菌株相比,MoRho2敲除突变体的菌丝生长速度、分生孢子形态、附着胞形态、产孢量、对植物的毒力等方面均无明显的差异,但分生孢子萌发和附着胞形成略滞后于野生型菌株。突变体能够产生正常的侵入栓并侵入洋葱表皮。综上所述,MoRho2基因可能参与稻瘟病菌分生孢子的萌发和附着胞的形成,结果为进一步揭示基因MoRho2的生物学功能奠定基础。
关键词 稻瘟病菌;MoRho2;基因敲除;功能分析
中图分类号 S435.111 文献标识码 A
Abstract Rice blast fungus, Magnaporthe oryzae, is an important model for the study of the interaction between host plant and pathogenic fungus. Rho-family proteins act as molecular switches in signal transduction. In this study, we applied bioinformatics and gene knock-out techniques to study the function of MoRho2 in the fungus. Bioinformatics analysis showed that the gene had no signal peptide and transmembrane. A slight delay of conidium germination and appressorium development and a slight decrease of penetration pegs were observed in MoRho2 knock-out mutant, when compared to wild-type strain.However, the mutant was not affected in growth rate, conidiation and pathogenicity, probably on account of the gene redundancy of MoRho2 in the fungal genome. Taken together, it appeared more likely that MoRho2 regulated conidium germination and appressorium development of rice blast fungus. These results would help to well understand the biological function and the molecular regulation mechanism of MoRho2.
Key words Magnaporthe oryzae; MoRho2; Gene knock-out; Functional analysis
doi 10.3969/j.issn.1000-2561.2016.10.024
Rho族蛋白是Ras超家族蛋白最主要成员之一,具有内在的GTP酶活性,该家族成员有GTP结合的激活态和GDP结合的失活态2种状态,它们之间的转换受GEF、GAP、GDI这3种调控蛋白的调控,在调控细胞过程中发挥分子开关的作用[1]。真核生物的大量研究结果表明,Rho族蛋白调控细胞骨架相关的信号传导途径,起着调节细胞形态的建成、细胞极性、细胞运动、细胞粘附、细胞增殖、基因转录等过程[2-3]。
近年来,真菌中Rho族蛋白研究取得许多进展,Rho1,Rho2,Rho3和Rho4与真菌的细胞分化与极性生长有关。酿酒酵母(saccharomyces cerevisiae)中,Rho1参与调节细胞完整性、细胞极性生长、肌动蛋白重排等过程[4];Rho2结合到前纤维蛋白上,调控肌动蛋白细胞骨架的重排[5];裂殖酵母(Schizosaccharomyces pombe)Rhol对隔膜的形成、细胞周期中细胞壁生长以及肌动蛋白细胞骨架的重排密切相关[6];Rho2参与调控细胞壁a-葡聚糖的生物合成,依靠调控胞浆移动对细胞分离进行调控[7];Rho3与细胞分裂有关,对细胞极性以及细胞壁完整性是必需的[8]。
稻瘟病菌(Magnaporthe oryzae)是研究植物病原真菌与寄主互作的理想模式生物之一[9-10]。 稻瘟病菌基因组数据库共有7个Rho族基因,分别是Rho1-4、RhoX、Rac1、Cdc42[11]。前期的研究结果表明,稻瘟病菌中MoRho3敲除突变体与野生型相比,附着胞形成滞后且形成率降低、对植物的致病性降低、分生孢子的形态从倒梨形突变成长椭圆形、产孢量减少、分生孢子萌发延迟等[12];稻瘟病菌中Rac1影响分生孢子形成、发育以及对植物的致病性[13];稻瘟病菌中MoCdc42敲除突变体的产孢量减少,分生孢子形态变细变长,影响植物的致病性[14]。基于以上研究,推测稻瘟病菌中Rho2蛋白同源物(MoRho2)在稻瘟病菌的生长发育中可能起到较为关键的作用,并有可能与其它Rho族蛋白共同在稻瘟病菌生长发育及其致病过程中的信号传导途径中发挥关键作用。本文对Rho2基因的功能进行分析,将有助于阐明稻瘟病菌Rho2所调控的生物学功能,为开展防治稻瘟病菌与生产应用提供一定的理论基础。
1 材料与方法
1.1 材料
1.1.1 供试菌株与质粒 稻瘟病菌野生型菌株△Ku70,MoRho2基因敲除突变体,MoRho2异位整合突变体均保存于本实验室;供试质粒pCX62保存于本实验室。
1.1.2 植物材料 水稻品种CO39和大麦品种Gold Promise均保存于本实验室。
1.2 方法
1.2.1 蛋白序列获得及分析 以稻瘟病菌基因组数据库中(http://broad.harvard.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)公布的MoRho2(MGG_02457.6)蛋白序列在NCBI(http://www.ncbi.nlm.nih.gov/)中用Blastp功能搜索其它真菌基因组数据库,获得其它真菌中的同源蛋白序列。用Clustal X和MEGA6.0软件对MGG_02457.6同源蛋白进行多序列比对和系统进化树的绘制。利用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)工具对其进行跨膜结构分析[15]。使用在线工具SignalIP 4.0(http://www.cbs.dtu.dk/services/SignaIP/)软件进行信号肽的预测与分析[15]。
1.2.2 MoRho2基因敲除突变体的获得 以稻瘟病菌株ΔKu70的基因组DNA为模板,采用引物MoRho2AF和MoRho2AR扩增稻瘟病菌MoRho2开放阅读框上游0.9 kb的片段,经XhoI单酶切后,插入至pCX62载体上,形成重组载体。同理,采用引物MoRho2BF和MoRho2BR,扩增MoRho2开放阅读框下游1.0 kb的片段,并克隆到pCX62载体的HindIII和SacI酶切位点之间,最后获得基因敲除载体PCX62-MoRho2-AB。以稀释后的基因敲除载体PCX62-MoRho2-AB为模板,采用引物MoRho2AF和MoRho2BR扩增MoRho2A-hph-MoRho2B片段。取适量MoRho2A-hph-MoRho2B片段,在PEG介导下,转化至新鲜制备好的稻瘟病野生型ΔKu70的原生质体[16],通过同源重组的原理进行基因敲除。利用PCR的方法筛选基因敲除的阳性转化子,筛选阳性转化子的PCR引物为MoRho2OF和MoRho2OR、MoRho2UA和H853。所用引物及目的见表1。
1.2.3 MoRho2基因突变体表型分析 (1)生长速度测定。采用Wu等[17]方法,取滤纸片保存的稻瘟病野生型和MoRho2各突变体菌株在淀粉酵母固体培养基(酵母粉2 g/L,可溶性淀粉10 g/L,蔗糖3 g/L,琼脂粉20 g/L)上活化,26 ℃温度下,生长3~4 d后,在菌落外缘用直径0.5 cm的打孔器打孔,将圆形菌丝块倒放在新的淀粉酵母琼脂培养基平板上,于28 ℃下倒置培养;分别在生长的第3、5、7、10天测量菌落的直径,培养至第10天时,进行拍照。
(2)产孢量分析及细胞形态观察。采用Wu等[17]方法,用直径0.5 cm的打孔器在已长满菌丝的淀粉酵母固体培养基(酵母粉2 g/L,可溶性淀粉10 g/L,蔗糖3 g/L,琼脂粉20 g/L)上打孔,然后将圆形菌丝块转接于直径9 cm含米糠固体培养基(米糠40 g/L,琼脂粉20 g/L,pH6.0)中,28 ℃光照培养14 d。用2 mL无菌水冲洗收集稻瘟病菌分生孢子,2层擦镜纸过滤孢子收集液。用血球计数板测得分生孢子数,再换算成单位面积产孢量。3次重复实验,结果取平均值。
(3)孢子萌发及附着胞形成实验。采用Nishimura等方法[18],孢子悬浮液浓度为1×104~5×104个/mL。在Gelbond film(BMA公司产品)疏水表面和亲水表面进行孢子萌发及附着胞形成实验,液滴大小为15 μL,表面各滴3滴,每个处理设置3个重复,25 ℃保湿培养,分别于4、8、12、24 h在显微镜下观察孢子萌发率及附着胞形成率。
(4)洋葱表皮侵染试验。采用Nishimura[18]等方法,孢子悬浮液浓度为2×104个/mL。采用24孔细胞培养板,在每孔中加入2 mL蒸馏水,切取大小适宜的洋葱表皮第三层的内表皮,置于蒸馏水上,将孢子悬浮液滴放在洋葱表皮上,大小为20 μL每滴,设置10个重复,保湿培养,分别在24 h及48 h显微镜下观察侵染情况。
(5)致病性分析。采用Xu等[19]离体接种方法:剪取五叶期的水稻品种CO39嫩叶或10 d苗龄的大麦品种Gold Promise叶片置于保湿处理的培养皿中。将菌丝块接种于叶片表面,每个样品重复3 个叶片,置于26 ℃黑暗培养24 h,光照培养5 d 后观察病情,拍照。
以上每个实验均做3 次重复试验。
2 结果与分析
2.1 MoRho2(MGG_02457.6)的生物信息学分析
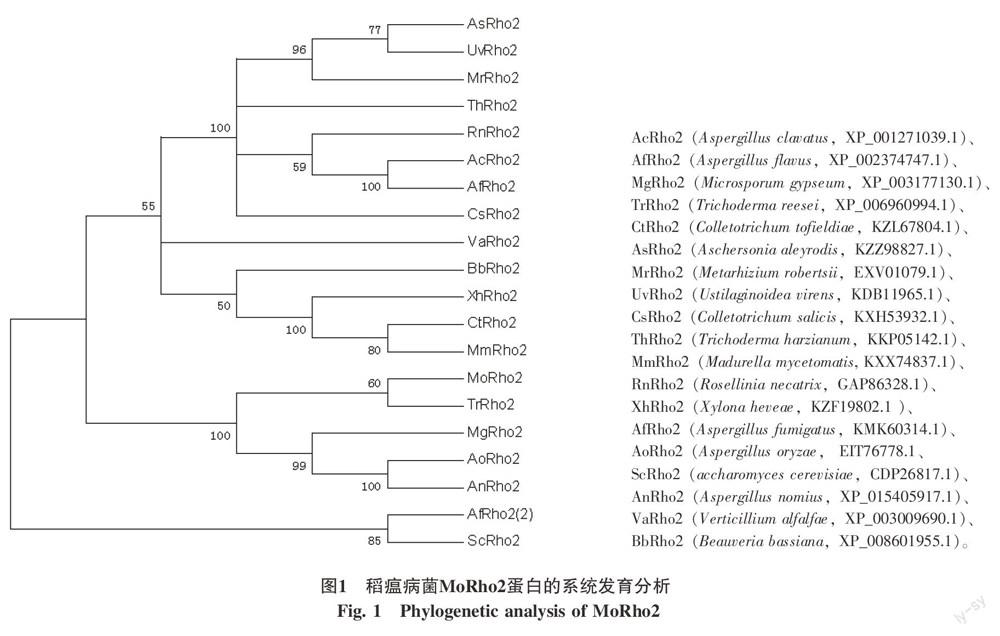
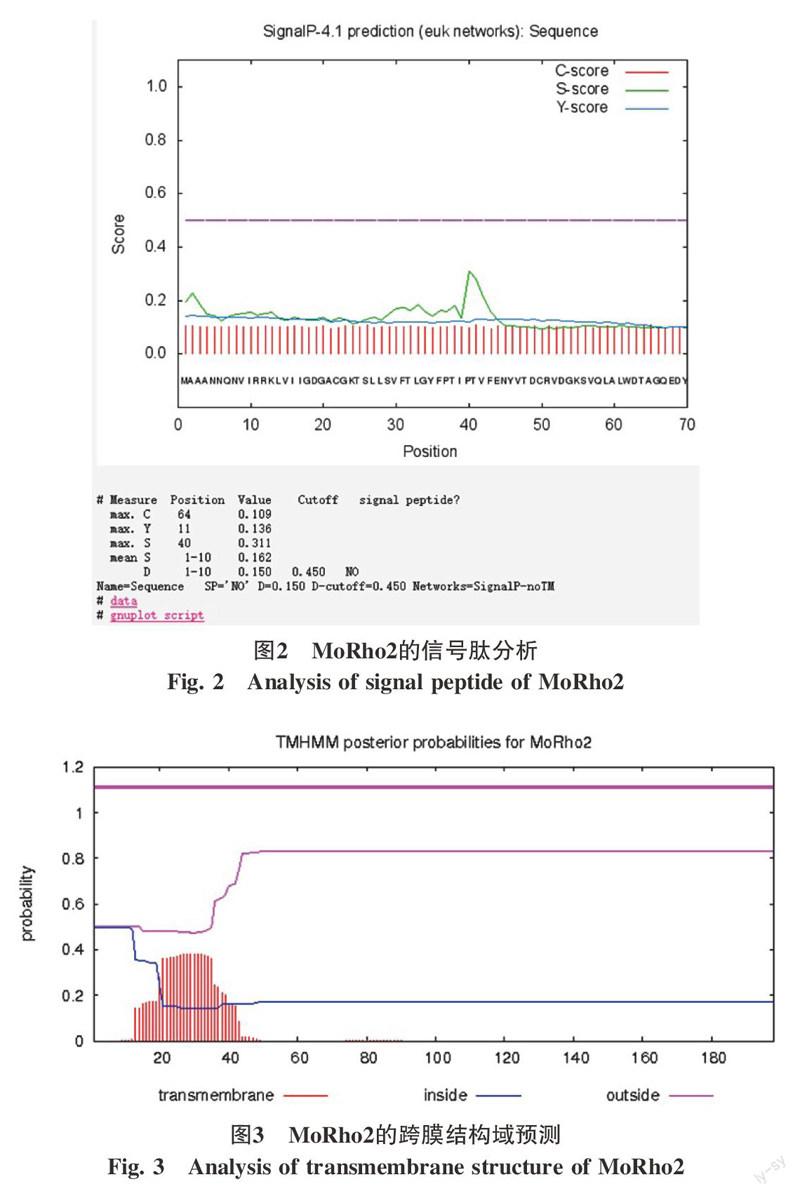
真菌与MGG_02457.6蛋白同源序列,用ClustalX软件和MEGA5.0软件采用临近法构建系统进化树的结果见图1。从图1可以看出,MoRho2与TrRho2在同一个分支上,说明这2个基因的亲缘关系较近(图1)。采用SignaIP 4.1在线软件对其进行信号肽预测,根据软件的默认参数,ORFs的Cmax 值<0.49,Smean值<0.5,表明MoRho2蛋白不存在信号肽(图2)。以TMHMM Server v.2.0对其进行跨膜域预测结果表明,该蛋白不含有跨膜区(图3)。
2.2 MoRho2基因敲除突变体获得
根据同源重组的原理(图4),构建了稻瘟病菌MoRho2基因敲除载体,转化至野生型菌株△Ku70的原生质体,转化子经400 μg/mL潮霉素和PCR分子验证,如图5所示转化子△MoRho2-1和△MoRho2-3的ORF均没有得到PCR产物,而得到了片段大小为1 117 bp的产物,转化子MoRho2-ect-4和MoRho2-ect-6获得了片段为745 bp的产物,说明获得了2个具潮霉素抗性的基因功能缺失转化子△MoRho2-1、△MoRho2-3和2个异位整合转化子MoRho2-ect-4、MoRho2-ect-6。滤纸片保存于-20 ℃,备用。
2.3 菌落生长速度及产孢量统计
CM琼脂培养基上的菌落生长速度表明,MoRho2基因敲除突变体与野生型菌株生长速度之间没有显著差异(图6;表2)。如稻瘟病菌野生型菌落和MoRho2基因敲除突变体菌株菌落的生长速度分别为(4.35+0.17)mm/d、(4.27+0.26)mm/d。
2.4 分生孢子形态、产孢量
进一步对ΔMoRho2基因突变体产孢能力进行分析,结果显示,突变体均能产生正常形态的分生孢子(图7,表3)和黑色素化的附着胞(图8,表4),产孢量相对野生型菌株也无明显变化(表2)。
2.5 分生孢子侵染洋葱表皮
将突变体的分生孢子接种洋葱表皮24 h后,发现野生型菌株ku70所产生的附着胞侵入栓的形成率是75.03%(表5),成功侵入到洋葱表皮细胞内,并进一步分化成侵染菌丝,在洋葱表皮细胞内侵染生长(图9),而MoRho2基因敲除突变体附着胞侵入栓的形成率是1.15%,另外MoRho2异味整合突变体附着胞侵入栓的形成率是45.05%;接种洋葱表皮48 h后,野生型菌株附着胞侵入栓的形成率是100%,而MoRho2基因敲除突变体附着胞侵入栓的形成率是48.19%,MoRho2异味整合突变体附着胞侵入栓的形成率是92.75%(表5)。可见,稻瘟病菌基因MoRho2影响分生孢子附着胞侵入栓产生的数量。
2.6 致病性分析
进一步分别用稻瘟病菌野生型和突变体菌株的菌丝块接种生长五叶期的水稻苗和10 d左右的大麦苗,结果表明,野生型和突变体均能在大麦和水稻叶片上产生典型病斑(图10)。可见,稻瘟病菌MoRho2基因与其致病性不相关。
3 讨论与结论
在丝状真菌中,Rho族蛋白是极性生长与细胞骨架重组过程中关键的调控因子。前人研究结果表明,在酿酒酵母(Saccharomyces cerevisiae)中,Rho2与肌动蛋白结合蛋白信号传导有关及调控细胞的形态建成[20-21];研究结果表明,稻瘟病基因组中有7个Rho族蛋白成员,且在稻瘟病菌中均有表达。稻瘟病菌Rho族蛋白MoRho3、MoRac1、MoCdc42在稻瘟病菌分生孢子形态建成、附着胞分化以及侵染性生长过程中起着非常重要作用[13-14,22]。因而推测Rho2蛋白可能与稻瘟病菌生长发育或致病过程有直接相关。
生物信息学结果表明,MoRho2蛋白为稳定的亲水蛋白,稳定系数为53.11,疏水性值为-0.092;分析特性表明,该蛋白不含有信号肽序列,不具有跨膜结构,不具有分泌途径,且无GPI锚定修饰,主要定位在细胞膜上。因此,该蛋白具有参与细胞的分化与极性生长的可能性。
然而,在本研究中,对突变体MoRho2表型分析结果表明,无论是在菌丝的生长速度、产孢能力以及致病性方面它们与野生型菌株均没有显著的差异。基于不同专家的研究成果,说明稻瘟病菌中有可能存在Rho基因功能冗余现象。因此,Rho族基因功能的鉴定对于探明真菌致病过程十分重要。本研究仅通过单基因敲除的方法进行功能研究,为进一步确定MoRho2基因参与信号传导过程,对该基因的分析可通过多个基因突变、蛋白互作等其他方法进一步研究。
参考文献
[1] Etienne-Manneville S, Hall A. Rho GTPases in cell biolog[J]. Nature, 2002, 420(6916): 629-635.
[2] Verhey K J, Gaertig J. The tubulin code[J]. Cell Cycle, 2007, 6(17): 2 152.
[3] Akhmanova A, Steinmetz M O. Tracking the ends: a dynamic protein network controls the fate of microtubule tips[J]. Nat Rev Mol Cell Biol, 2008, 9(4): 309.
[4] Drgonova J, Drgon T, Roh D H, et al. The GTP-binding protein Rho1p is required for cell cycle progression and polarization of the yeast cell[J]. J Cell Biol, 1999, 146(2): 373-387.
[5] Marcoux N, Cloutier S, Zakrzewska E, et al. Suppression of the Profilin-Deficient Phenotype by the RHO2 Signaling Pathway in Saccharomyces cerevisiae[J]. Genetics, 2000, 156: 579-592.
[6] Nakano K, Arai R, Mabuchi I. The small GTP-binding protein Rho1 is a multifunctional protein that regulates actin localization, cell polarity, and septum formation in the fission yeast Schizosaccharomyces pombe[J]. Genes Cells, 1997, 2(11): 679-694.
[7] Robinson N G, Guo L, Imai J, et al. Rho3 of Saccharomyces cerevisiae, which regulates the actincytoskeleton and exocytosis, is a GTPase which interacts with Myo2 and Exo70[J]. Molecular and Cellular Biology, 1999, 19(5): 3 580-3 587.
[8] Adamo J E, Rossi G, Brennwald P. The Rho GTPase Rho3 has a direct role in exocytosis that is distinct from its role in actin polarity[J]. Molecular Biology of the Cell, 1999, 10(12): 4 121-4 133.
[9] Dean R A, Talbot N J, Ebbole D J, et al. The genome sequence of the rice blast fungus Magnaporthe grisea[J]. Nature, 2005, 434: 980-986.
[10] Valent B, Chumley F G. Molecular genetic analysis of the rice blast fungus Magnaporthe grisea, Annu[J]. Rev Phytopathol., 1991, 29: 443-467.
[11] 郑 武, 陈继圣, 郑士琴, 等. 基因表达分析稻瘟菌7个Rho GTP酶的关系[J]. 中国农业科学, 2006, 39: 2 237-2 242.
[12] 郑 武, 稻瘟病菌Rho族GTP酶功能分析[D]. 福州: 福建农林大学, 2006.
[13] Chen J, Zheng W, Zheng S, et al. Rac1 is required for pathogenicity and Chm1-dependent conidiogenesis in rice fungal pathogen Magnaporthe grisea[J]. PLoS Pathog, 2008, 4(11): e1000202.
[14] Zheng W, Chen J S, Liu W D, et al. A Rho3 homolog is essential for appressorium development and pathogenicity of Magnaporthe grisea[J]. Eukaryot Cell, 2007, 6: 2 240-2 250.
[15] 王铁霖, 李 晶, 杨玉文, 等. 烟草野火病菌Pseudomonas syringae pv. tabaci yuexi-1信号肽预测及分析[J]. 中国烟草学报, 2016, 22(1): 92-100..
[16] Zheng W H, Zhou J, He Y l, et al. Retromer Is essential for autophagy dependent plant infection by the Rice Blast Fungus[J]. Plos Genetics, 2015, DOI: 10.1371/journal.pgen.1005704.
[17] Wu L M, Bian W Y, Hu F H, et al. Functional analysis of a putative phenylacetone monooxygenase gene(MoPAMO1)in Magnaporthe oryzae[J]. Agricultural Biotechnology, 2013, 5(3): 595-602.
[18] Nishimura M, Park G, Xu J R, The G-beta subunit MGB1 is involved in regulating multiple steps of infection-related morphogenesis in Magnaporthe grisea Mol[J]. Microbiol, 2003, 50: 231-243.
[19] Xu J R, Hamer J E. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe oryzae[J]. Genes Dev, 1996, 10(21): 2 696-2 706.
[20] Park H O, Bi E. Central roles of small GTPases in the development of cell polarity in yeast and beyond[J]. Microbiol Mol Biol Rev, 2007, 71: 48-96.
[21] Marcoux N, Cloutier S, Zakrzewska E, et al. Suppression of the Profilin-deficient phenotype by the RHO2 signaling pathway in Saccharomyces cerevisiae[J]. Genetics, 2000, 156: 579-592.
[22] Zheng W, Zhao Z, Chen J, et al. A Cdc42 ortholog is required for penetration and virulence of Magnaporthe grisea[J]. Fungal Genetics and Biology, 2009, 6(15): 450-460.
