拟南芥AP3基因植物表达载体的构建及在烟草中的遗传转化
2016-05-30刘彩霞代丽娟刘轶葛晓兰曲冠证
刘彩霞 代丽娟 刘轶 葛晓兰 曲冠证
摘要:拟南芥花器官B类特征基因属于MADS-box基因家族,其中APETALA3(AP3)基因在花瓣和雄蕊中特异性地表达。为探讨AP3基因在花瓣和雄蕊发育中的功能,本研究从拟南芥(Arabidop.sis thaliana)花序中克隆AtAP3基因构建植物表达载体,并转化烟草(Nicotiana tobacum)。通过PCR及qRT-PCR分析,表明AtAP3基因成功转入烟草基因组并表达转录。表型观测显示转基因烟草雄蕊与野生型相比明显变短,说明AP3基因特异性地参与雄蕊的发育并起着至关重要的作用。
关键词:拟南芥;APETALA3(AP3);转基因;烟草;雄蕊发育
中图分类号:S188+.1
文献标识号:A 文章编号:1001-4942(2016)02-0007-05
花是被子植物的重要生殖器官。花器官的发育模型最早源于Coen和Meverowitz对拟南芥和金鱼草的研究,并提出了花发育的“ABC”模型,随后该模型被后人逐步发展为“ABCD”或“ABCE”模型。典型的花发育具体可分为四轮,分别为萼片、花瓣、雄蕊和心皮,每一轮的发育都受相应基因的特异性调控。
隶属于MADS-box家族的植物花器官B类特征基因在雄蕊及花瓣发育中行使功能。研究表明,B类功能基因在花器官发育时与C类功能基因共同控制雄蕊发育,同时又与A类功能基因共同调控花瓣发育,当B类功能基因缺失,雄蕊会转化为心皮,花瓣则转化为萼片,形成只有心皮和萼片的不完整花器官。B类功能基因在植物进化过程中发生了两次基因重复事件,结果在这个亚家族中产生了四个不同的进化系,分别为DEF/AP3(paleoAP3)、GLO/PI、TM6和euAP3。其中拟南芥经历基因重复后,其B类功能基因产生了属于euAP3进化系成员的APETAIA3(AP3)基因和属于GLO/PI进化系成员的PISTELIATA(PI)基因。
目前对花器官发育的研究大多以“ABC”模型为理论基础来丰富其他物种的花器官发育机制,关于拟南芥AP3基因在烟草中异源表达的研究报道较少。本试验根据拟南芥B类功能基因的高度保守性及组织表达特异性,从拟南芥花序中克隆得到AtAP3基因,构建植物表达载体并进行烟草遗传转化。不仅对植物花器官B类特征基因AP3的功能进行了新的补充,还增加了对植物花发育“ABC”模型的新认识。
1 材料与方法
1.1 试验材料
野生型拟南芥(Arabidopsis thaliana Col-0)、野生型烟草(Nicotiana tobacum)种子均为本实验室保存。
1.2 菌株及主要试剂
大肠杆菌Transl-T1感受态购自Transgen(中国);农杆菌EHA105由本实验室保存;植物表达载体pROKII质粒,由山东师范大学张慧教授惠赠;质粒提取、胶回收试剂盒购自OMEGA公司(美国);PCR相关试剂、DNA marker、限制性内切酶、T4 DNA ligase、PrimeScriptrrM RT reagent Kit购自TaKaRa公司(中国大连);EasyPureTM PlantRNA Kit、pEASY-T1购自Transgen(中国);其他试剂为进口或国产分析纯。
1.3 试验方法
1.3.1 拟南芥花序总RNA提取及AtAP3基因的克隆 用EasyPure'rM Plant RNA Kit试剂盒提取拟南芥花器官的总RNA,采用PrimeScriptTM RT rea-gent Kit试剂盒进行cDNA合成。以cDNA为模版,利用引物AtAP3-F和AtAP3-R(表1)进行PCR扩增。反应程序:94℃预变性4min;94℃变性30s,55℃退火30s,72℃延伸1min,共35个循环;72℃再延伸7min,PCR结束后全部产物进行1%琼脂糖凝胶电泳分析。利用凝胶回收试剂盒回收目的片段,将回收后的片段根据操作手册介绍方法连人pEASY-T1载体中,转化大肠杆菌Transl-T1感受态,涂LB抗性平板(卡那抗性),对获得的单克隆进行菌落PCR验证,筛选后的阳性克隆命名为pEASY-T1-AtAP3,送至哈尔滨博仕生物技术有限公司测序。
1.3.2 植物表达载体构建利用限制性内切酶Xba I、Kpn I双酶切重组质粒pEASY-T1-A-tAP3,同时将pROKII载体质粒也用XbaI、Kpn I双酶切,并分别回收目的片段,利用T4 DNA ligase过夜连接转化大肠杆菌Transl-T1感受态,涂LB抗性平板(卡那抗性),对获得的单克隆进行菌落PCR验证,筛选后的阳性克隆命名为pROKII-AtAP3,提取质粒,送至哈尔滨博仕生物技术有限公司测序。选取测序正确的质粒转化农杆菌EHA105,提取农杆菌中的质粒利用引物pROKII-F和pROKII-R(表1)进行PCR检测。
1.3.3 转基因株系获得及分子检测 通过根瘤农杆菌介导法进行烟草的遗传转化,转化过程如下:预培养1~2d;农杆菌(OD=0.2)侵染3min;共培养1~2d;脱菌。一个月后获得抗性株系。用CTAB方法提取抗性植株的总DNA,利用引物pROKII-F和pROKII-R进行PCR,并用1%琼脂糖凝胶检测,分析是否成功获得转基因株系。
1.3.4 实时定量RT-PCR分析检测 用EasvPureTM Plant RNA Kit试剂盒提取转基因植株的总RNA,经反转录试剂盒进行cDNA合成。将合成的第一链cDNA加去离子水稀释10倍后作为模板,进行定量检测,引物为AtAP3-RT-F和AtAP3-RT-R(表1)。反应体系为20μL:其中2×SYBR Green实时荧光染料混合液10μL,ROXDye Ⅱ0.4μL,2μL cDNA模板,正、反向引物各0.8μL。以NtActin基因作为内参,扩增反应条件为:95℃30s;95℃5s,60℃35s,40个循环;95℃15s,60℃lmin,95℃15s。每个样品进行3次重复,并通过2-△△Ct法进行计算。
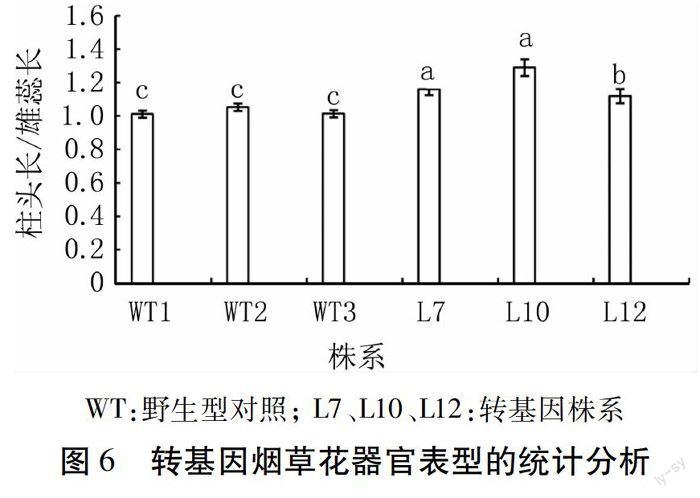
1.3.5 转基因植株的表型观测 挑选目的基因表达量较高的3个转基因株系及对照株系分别移栽10棵至温室,对其进行定期观察,待进入盛花期后,在顶部花序中,每株随机选取5朵刚刚开放的花,分别测量其花瓣、柱头及雄蕊的长度,对测量结果利用SPSS 19.0软件进行SNK差异性统计分析。
2 结果与分析
2.1 拟南芥花序AtAP3基因的克隆
利用RNA提取试剂盒从拟南芥花序中提取总RNA,经浓度及质量检测可用于下一步试验。将提取的RNA经反转录试剂盒反转录为cDNA作为模板,利用引物AtAP3-F和AtAP3-R进行PCR扩增,1%琼脂糖电泳检测表明,扩增条带清晰且和预期大小一致(图IA)。将目的条带切胶回收后与克隆载体pEASY-T1连接,转化大肠杆菌,经菌落PCR验证(图1B),将获得的阳性克隆命名为pEASY-T1-AtAP3。
2.2 植物表达载体的构建
对获得的pEASY-T1-AtAP3质粒和pROKII载体质粒,同时利用限制性内切酶XbaI、Kpn I进行双酶切,反应结束用1%琼脂糖凝胶电泳检测(图2A),分别回收目的片段。利用T4 DNA ligase进行连接反应后,转化大肠杆菌Transl-T1感受态,挑取单菌落进行PCR检测,获得750 bp左右目的条带(图2B),测序结果显示连接正确,基因未发生突变。提取质粒转化农杆菌EHA105,挑取单克隆并进行PCR检测(图2C),结果显示所选单菌落均为阳性克隆,表明植物表达载体构建成功。
2.3 转基因植株的获得及PCR检测
用含有pROKII-AtAP3的农杆菌EHA105侵染烟草叶片,经4~6周的选择培养,从叶片周围生成大量抗性芽。待抗性芽长出叶片后,将叶片切下放在含有Kan(50mg·L-l)抗生素的分生培养基上继代培养。将经两次分化获得的芽在生根培养基中进行生根定植,最终共获得18株转基因烟草植株。提取抗性转基因株系的基因组DNA,利用表1中的pROKII通用引物对其进行PCR扩增检测(图3),结果显示抗性植株中均含有目的基因,说明外源基因已经整合人烟草基因组中。
2.4 转基因植株的荧光定量PCR检测
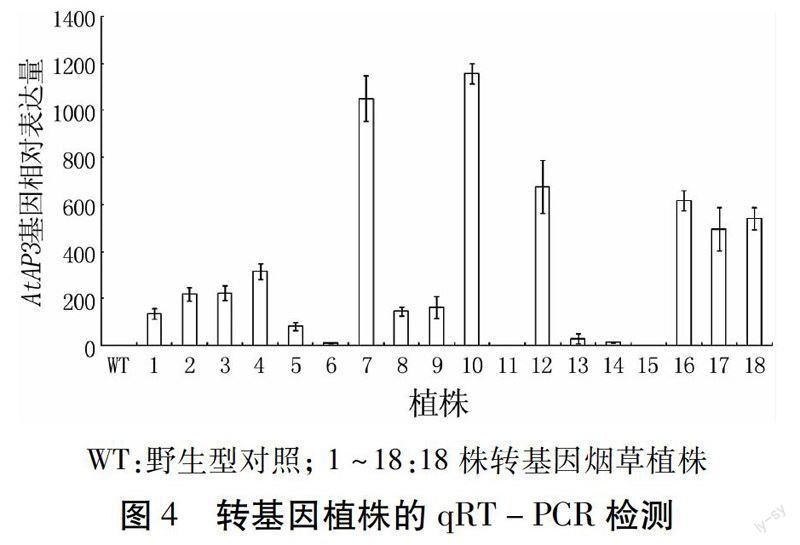
为了比较AtAP3基因在烟草植株中的表达情况,提取烟草叶片总RNA进行荧光定量PCR检测,结果如图4。所有的转基因株系中均有AtAP3的表达,其中7、10、12、16、17、18号株系表达量最高;1、2、3、4、5、8、9号株系表达量低于以上几个株系;表达量最低的株系是6、1 1、13、14、15。
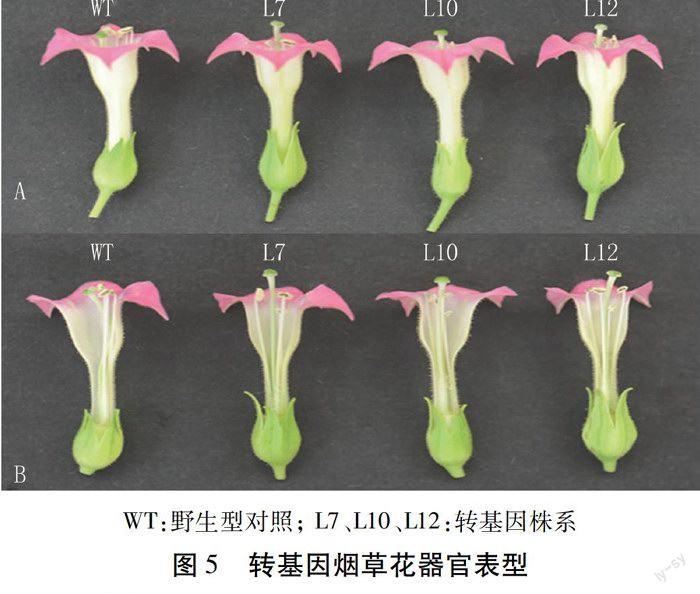
2.5 转基因株系表型观察
经过定量PCR检测,筛选目的基因表达量高的3个转基因株系和野生型烟草,移栽到温室中。表型观测结果显示,在营养生长期,转基因植株与野生型相比无明显性状差异;待进入花器官发育成熟期后,转基因株系雄蕊变短(图5)。分别选取3个转基因株系和对照株系(每个株系顶端随机取5朵花),测量柱头和雄蕊的长度,用每一朵花的柱头长度与其雄蕊长度做比值,再取比值的平均值,利用SPSS 19.0软件,采用SNK方差齐性检验的方法对其差异显著性进行分析,结果如图6。转基因株系与野生型株系比,雄蕊变短,且差异显著;3个转基因株系之间L10>L7>L12,与实时定量结果一致,表明AtAP3基因表达量越高雄蕊缩短越明显。上述结果表明,过表达AtAP3基因影响转基因烟草植株雄蕊的发育,进而导致转基因株系结种率下降。
3 讨论
自“ABC”模型提出之后,许多人都从这三类特征基因着手,研究不同物种花器官发育机制,得到了许多类似的基因。基于植物之间的亲缘远近、物种差异、进化程度强弱等特点,同一种功能基因会出现序列差异,但该类基因均保留有相同的保守区域,来保证行使相应的功能。其中属于MADS-box家族的B类特征基因在花瓣和雄蕊的发育中行使功能,其在拟南芥中包括PI和AP3两个成员。被子植物的B类功能基因在具有高度保守性的同时随物种的进化等原因产生序列多态性,其中包括一个PI基因系和三个AP3-like基因系,且AP3/PI基因较其他功能基因的进化速度快40%左右,这使得B类功能基因产生多元结构,主要表现为雄蕊及花瓣的形态多样性。
本试验为探究AtAP3基因异源表达的功能,从拟南芥花序中克隆得到AtAP3基因,并将其转人烟草,与野生型相比,转基因烟草的花器官发生明显变化,主要表现为雄蕊的长度短于对照植株,花冠长度也有变短现象,但变化不明显。前人研究表明,花瓣是由A类功能基因与B类功能基因中的PI基因共同作用形成的,而雄蕊则是由B类功能基因中的AP3基因与C类功能基因共同作用形成的,本研究中雄蕊发育发生变化导致转基因植株的育性下降甚至败育,这与AP3基因调控雄蕊发育的理论相一致,表明AtAP3在异源调控植物的花器官发育方面有一定作用,AtAP3基因与烟草中的B类基因和C类基因共同作用从而抑制雄蕊发育。但外源AtAP3基因在转基因烟草中是通过怎样的方式调控雄蕊形态变化仍需进一步深入探究,以期对该现象作出机理性的阐述,挖掘出其潜在的应用价值。
