生境及气象因子对南瓜实蝇雄成虫数量动态的影响
2016-05-30李磊牛黎明张方平韩冬银陈俊谕符悦冠
李磊 牛黎明 张方平 韩冬银 陈俊谕 符悦冠
摘 要 监测不同生境下南瓜实蝇雄成虫的数量动态,分析影响南瓜实蝇数量变动的关键气象因子。结果表明:生境和气象因子显著影响南瓜实蝇雄虫的数量动态。混合菜地、园林区以及混合果园诱集的南瓜实蝇雄虫数量多于住宅区、农贸市场、苦瓜种植地同期诱集的雄虫数;通过逐步回归分析和通径分析得出影响南瓜实蝇雄虫数量动态的气象因子有月平均气温、月平均气压、月平均相对湿度、月最小相对湿度和月日照时数,其中月平均温度是影响其数量动态的最关键气象因子。基于本研究的结果,建议对南瓜实蝇的防控应不仅仅局限在具有其寄主植物的生境,而应根据监测的数据对其他生境的南瓜实蝇进行一并治理。
关键词 南瓜实蝇;生境;气象因子;数量动态
中图分类号 S433 文献标识码 A
Effect of Habitats and Climatic Factors on the Quantitative
Dynamics of Male Bactrocera tau(Walker)
LI Lei, NIU Liming, ZHANG Fangping, HAN Dongyin, CHEN Junyu, FU Yueguan*
Environment and Plant Protection Institute, CATAS / Key Laboratory of Integrated Pest Management
for Tropical Crops of Ministry of Agriculture, Haikou, Hainan 571101, China
Abstract The quantitative dynamics of male B. tau were monitored in different habitats. Then the key climatic factors affecting the quantitative fluctuation of B. tau were analyzed. The results showed that habitats and climatic factors significantly influenced the quantitative dynamics of male B. tau. The number of B. tau in the mixed vegetable plot, landscape area and mixed fruit orchard were more than those in the residential area, farmers market and field planted bitter gourd. The stepwise regression analysis and path analysis indicated that the quantitative fluctuation of male B. tau was controlled by mean monthly temperature, monthly atmospheric pressure, monthly relative humidity, monthly minimum relative humidity and monthly sunlight hours, hereinto, mean monthly temperature was the most important factors. Based on these results, the control of B. tau was not limited in its hostplant habitats. Bactrocera tau should be managed in other habitats arrcording to its monitoring data.
Key words Bactrocera tau; Habitat; Climatic factor; Quantitative dynamics
doi 10.3969/j.issn.1000-2561.2016.09.022
南瓜实蝇(Bactrocera tau)是一种世界性的检疫性害虫,最早于1849年在中国报道,现广泛分布在东南亚、南太平洋地区[1-4]。在中国,该虫主要分布在海南、广东、广西、福建、台湾、云南、贵州、四川、重庆、浙江、江西、湖北、湖南等地,其中华南和西南地区的省市受害最为严重[1-2,5]。南瓜实蝇寄主植物广泛,可为害16个科80多种蔬菜和水果,尤其嗜好为害葫芦科植物果实[2,6]。该虫偏好将卵产于果实表皮下层组织中,幼虫孵化后果实内部蛀食,常常导致果实提前黄化、畸形或脱落,严重影响果实的产量和品质[2,7],进而造成严重的经济损失[8-9]。
摸清南瓜实蝇的发生动态和影响其发生动态的气象因子有助于规划科学的防治对策。南瓜实蝇一年可发生3~8代,其发生高峰因地而异,如浙江台州黄岩地区发生高峰为9~12月、重庆发生高峰为7~9月等,但发生高峰大多集中在3~12月[2,5,10-12]。儋州是南瓜实蝇在海南发生的主要区域之一,但该虫在儋州的发生动态不明确。通过拟信息素或食物饵剂定时定点监测南瓜实蝇的种群动态发现该虫的数量动态受日降雨量、降雨次数、日平均温度、种植的寄主种类等因素影响[5,13-14],但上述研究仅局限于某几个月份和某个生境,因此无法准确找出在多种生境条件下影响南瓜实蝇全年种群动态的关键气象因子。基于此,本研究于2014年3月~2016年2月在儋州监测了不同生境下南瓜实蝇雄成虫的数量动态,分析了影响南瓜实蝇数量变动的关键气象因子,以期为科学防控南瓜实蝇提供参考。
1 材料与方法
1.1 材料
1.1.1 供试材料和气象资料 供试南瓜实蝇雄虫引诱剂为诱蝇酮,又称Cuelure,由广州瑞丰生物科技有限公司提供,纯度≥98%;供试瓶式诱捕器(B型)由广州瑞丰生物科技有限公司提供。
监测地点的气象资料由海南省气象局提供。
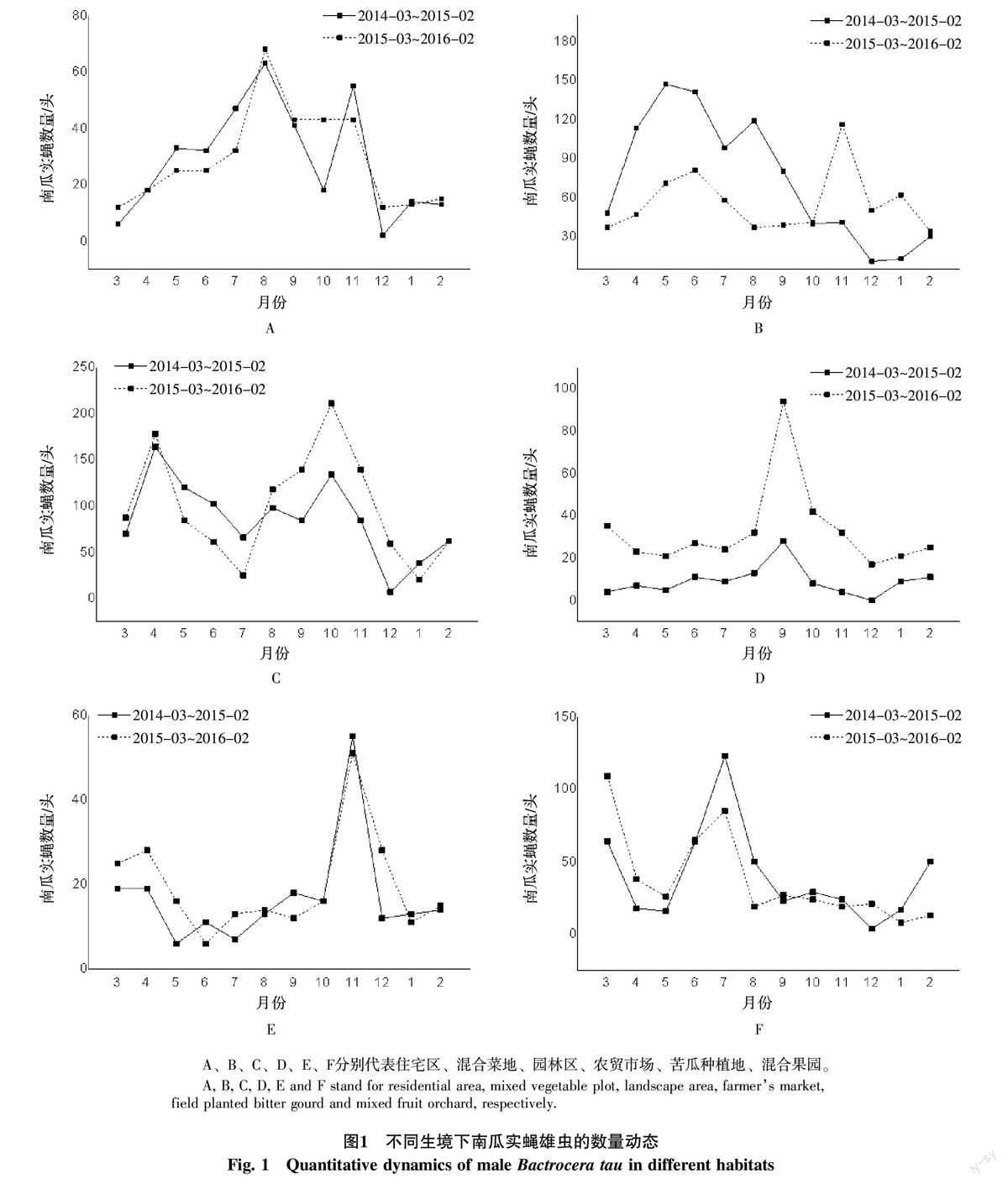
1.1.2 监测时间和监测生境概况 试验于2014年3月~2016年2月在海南省儋州市宝岛新村进行,整个监测地点以中国热带农业科学院儋州院区(简称院区)总部大楼为中心向四周辐射,分住宅外围区(简称住宅区)、混合菜地(多种蔬菜种植地,简称混合菜地)、园林作物种植区(简称园林区)、农贸市场外围区(简称农贸市场)、苦瓜种植地、混合果园(多种果树种植园)等6个不同生境监测点同时进行,监测总面积120 hm2以上。住宅区位于院区的东部住宅区,每栋楼间种有稀疏的大王棕、散尾葵、凤凰木、印度紫檀等行道树;混合菜地毗邻住宅区的南面和北面,长年种植各类蔬菜,监测期间种植的蔬菜有豇豆、萝卜、白菜、黄瓜、丝瓜、苦瓜、葫芦瓜、南瓜、四季豆等;园林区位于院区内的海南热带植物园周边,园内和周边主要种植1 000多种热带植物,以乔木为主;农贸市场位于院区西面,主要经营蔬菜、水果、肉禽及其他农产品等,周边零星种植大王棕、凤凰木等行道树;苦瓜种植地位于院区的丘凡试验基地,监测期间主要成片种植苦瓜;混合果园距园林区向东1 km左右的中国热带农业科学院热带作物品种资源研究所的果树基地,里面主要种植大片的芒果、黄皮、人心果以及零星的杨桃和莲雾。
1.2 方法
将南瓜实蝇雄虫引诱剂滴于诱捕器中的高密度纤维板上,诱捕器悬挂于各个监测点的阴凉处,距离地面约1.5 m,每个监测点4个诱捕器,每个诱捕器间隔约50 m。每隔10 d统计诱捕器内的南瓜实蝇数量,每隔20 d添加1次诱剂,每半年更换1次诱捕器。
1.3 数据统计与分析
不同生境每月南瓜实蝇雄虫数量由各生境所有诱捕器诱集的南瓜实蝇雄虫总数得来,儋州南瓜实蝇雄虫数量动态由6种生境下诱集的南瓜实蝇雄虫总数得来。通过SAS 9.0进行数据统计分析,以每月南瓜实蝇雄虫诱集的数量为因变量,以气象因子为自变量进行相关性分析、逐步回归分析以及通径分析,并对观测的南瓜实蝇发生数量与理论的发生数量进行适合性检验。采用Origin 8.5进行绘图。
2 结果与分析
2.1 生境对瓜实蝇雄虫数量动态的影响
由图1可知,不同生境下南瓜实蝇的发生高峰和低谷出现的时间不同,但不同年份间相同生境相同月份的南瓜实蝇发生动态趋势相似。在住宅区,南瓜实蝇的发生高峰时间为8月和11月,出现低谷时间为12月、1月、2月、3月;混合菜地2014年3月~2015年2月南瓜实蝇的发生高峰时间为5月和6月,出现低谷时间为12月和翌年1月,而2015年3月~2016年2月的发生高峰时间为5月、6月和11月,出现低谷时间为12月和翌年的1月、2月、3月;园林区南瓜实蝇的发生高峰为4月和10月,出现低谷的时间为7月、12月和翌年的1月;农贸市场区南瓜实蝇发生高峰为9月,其余月份发生量均相对较低;苦瓜种植地南瓜实蝇发生高峰为11月份,除3、4月份外其余月份的发生量均相对较低;混合果园2014年3月~2015年2月南瓜实蝇的发生高峰为3月、7月和翌年的1月,其余发生量都相对较低,而2015年3月~2016年2月的发生高峰为3月和7月,其余发生量都相对较低。从图1也可发现混合菜地、园林区以及混合果园诱集的南瓜实蝇数量显著多于在住宅区、农贸市场、苦瓜种植地诱集的数量。
2.2 气象因子对南瓜实蝇雄虫数量动态的影响
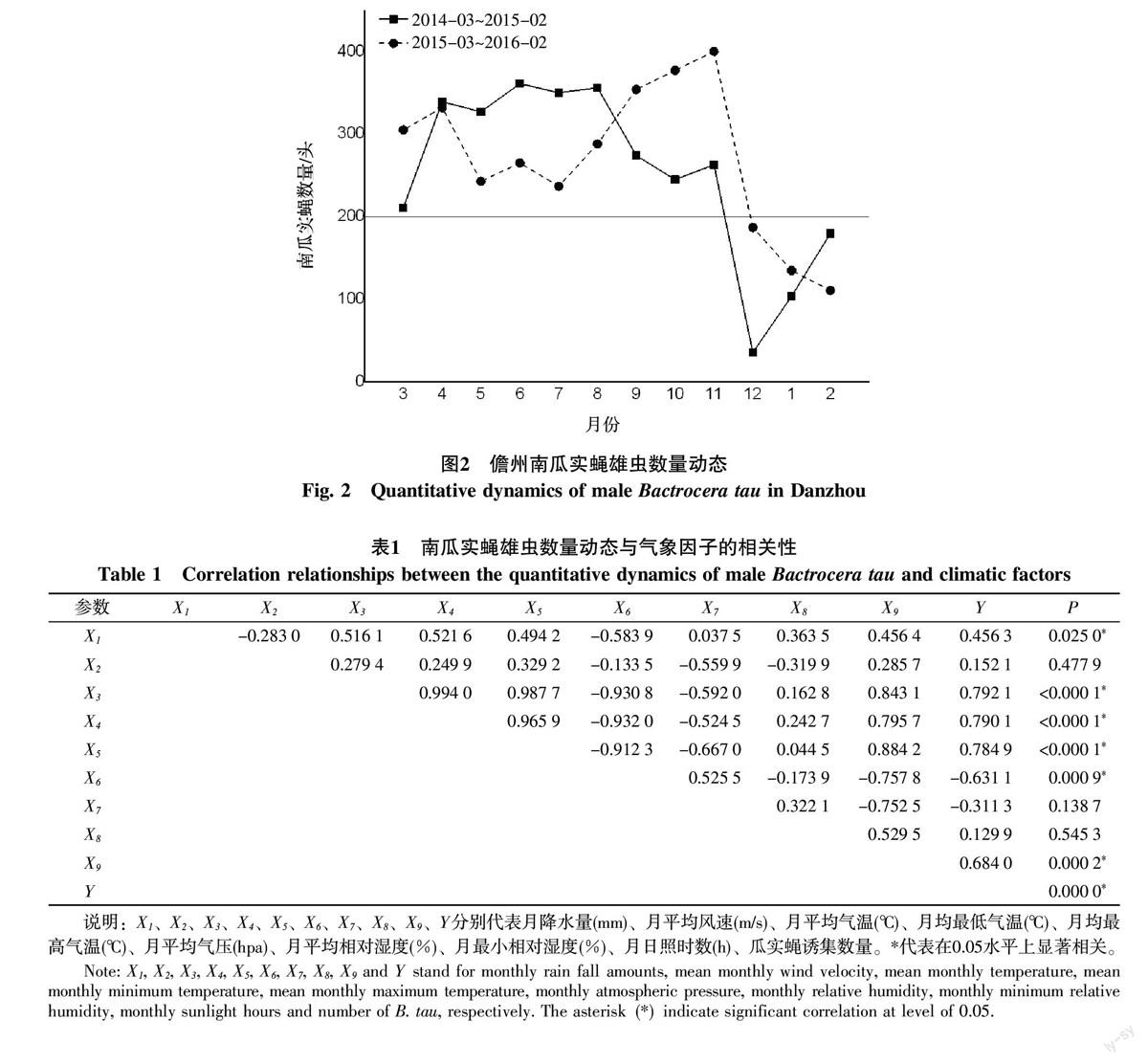
由图2可知,南瓜实蝇雄虫的发生高峰期主要从3月份开始波动发生至11月份,每月诱集的雄虫数量均大于200头。而从12月份起至翌年2月,南瓜实蝇的每月诱集数量均低于200头。
将南瓜实蝇雄虫诱集总数(Y)与月降水量(X1)、月平均风速(X2)、月平均气温(X3)、月均最低气温(X4)、月均最高气温(X5)、月平均气压(X6)、月平均相对湿度(X7)、月最小相对湿度(X8)和月日照时数(X9)9种气象因子进行相关性分析发现(表1):南瓜实蝇雄虫诱集的数量与月降水量(X1)、月平均气温(X3)、月均最低气温(X4)、月均最高气温(X5)、月日照时数(X9)成正相关,与月平均气压(X6)成负相关,且相关性均达到显著水平,说明这些气象因子显著影响了南瓜实蝇雄虫的数量动态。此外,由表1也可知南瓜实蝇雄虫诱集数与月平均风速(X2)、月最小相对湿度(X8)成正相关,与月平均相对湿度(X7)成负相关,但相关性均未达到显著水平,说明这3种气象因子对南瓜实蝇雄虫的数量动态影响较小。
以南瓜实蝇诱集数量为因变量,以气象因子Xi为自变量进行逐步回归分析,得到方程为:Y=-16 482+42.287 3X3+14.573 4X6+17.010 2X7-5.448 7X8+0.529 1X9(R=0.844 3,F=19.51,P<0.000 1),这表明月平均气温(X3)、月平均相对湿度(X7)、月最小相对湿度(X8)和月日照时数(X9)综合影响了南瓜实蝇雄虫的数量动态。
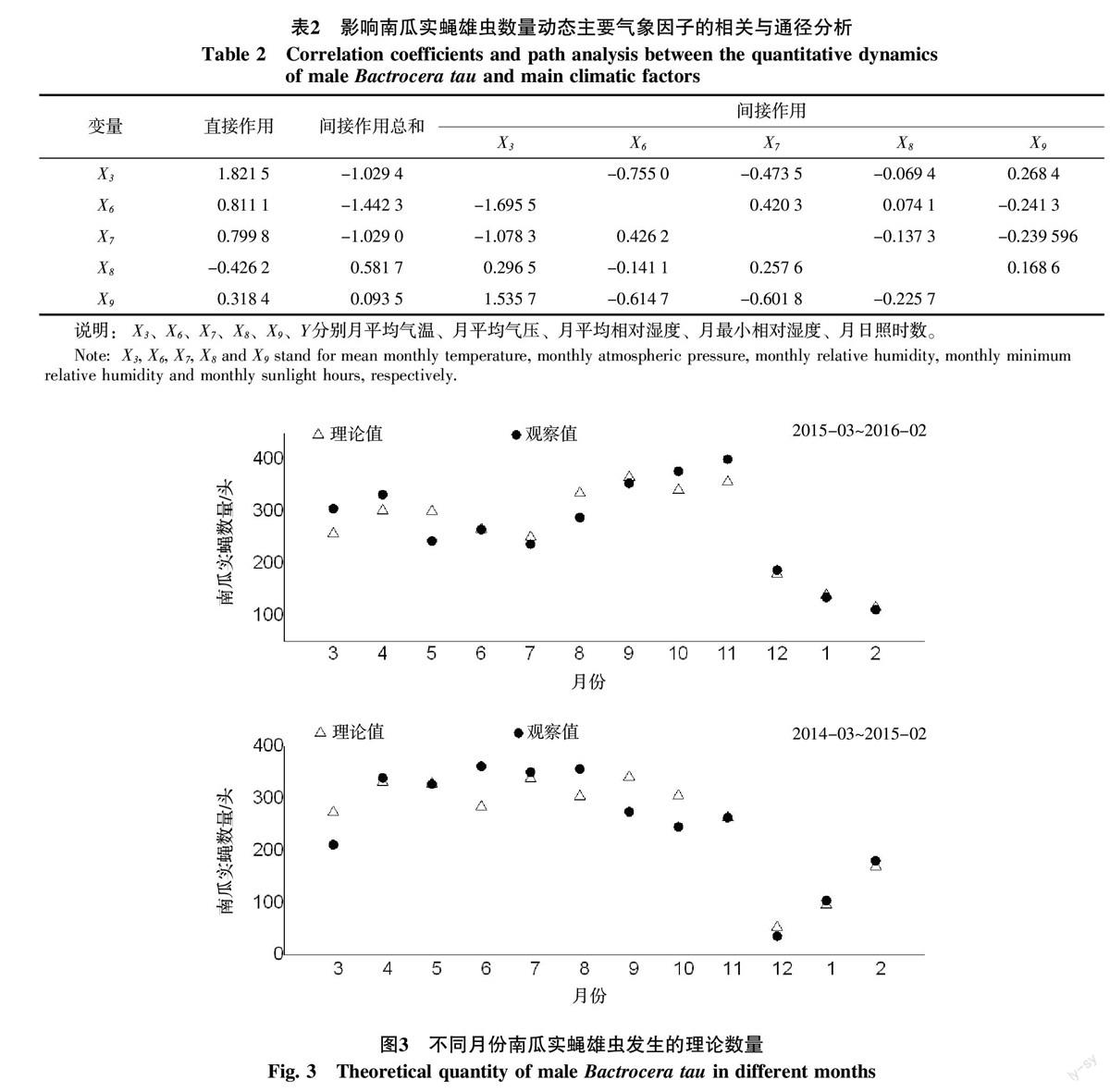
根据月平均气温(X3)、月平均气压(X6)、月平均相对湿度(X7)、月最小相对湿度(X8)和月日照时数(X9)与南瓜实蝇雄虫诱集数(Y)的相关系数剖分为直接作用和通过其他气象因子(Xi)的间接影响2个部分进行统计分析得出(表2):月平均气温(X3)对南瓜实蝇雄虫数量动态影响最大,其余依次为月平均气压(X6)、月平均相对湿度(X7)、月日照时数(X9)、最小相对湿度(X8),其中月平均气温(X3)、月平均气压(X6)、月平均相对湿度(X7)、月日照时数(X9)对南瓜实蝇雄虫数量动态影响的直接作用大于通过其他气象因子影响的间接作用,最小相对湿度(X8)对南瓜实蝇数量动态影响的间接作用大于直接作用。
综合分析表明,影响南瓜实蝇雄虫数量动态的气象因子有月平均气温(X3)、月平均气压(X6)、月平均相对湿度(X7)、月最小相对湿度(X8)和月日照时数(X9),这些气象因子除单独地作用于南瓜实蝇雄虫数量动态外,彼此间相互影响、相互关联对南瓜实蝇的种群变动形成了综合效应。
将影响南瓜实蝇雄虫数量动态的关键气象因子依次代入上述拟合的方程得到不同月南瓜实蝇雄虫发生的理论数量(图3),经T检验发现南瓜实蝇发生的理论数量与实际数量间无显著差异(2014-03~2015-22:t=0.075 0,P=0.941 0;2015-03~2016-02:t=-0.063 0,P=0.950 0),这也表明上述拟合的回归方程能准确反应南瓜实蝇雄虫数量动态与气象因子的关系。
3 讨论与结论
由于本研究选用了南瓜实蝇雄虫引诱剂诱蝇酮监测其数量动态,故所诱集的南瓜实蝇基本为雄虫,但调查中发现南瓜实蝇在自然界中的雌雄性比接近1 ∶ 1,因此以其雄虫的诱集数量也可反应出南瓜实蝇在自然界中的数量动态,这与利用诱蝇酮监测瓜实蝇(B. Cucurbitae)和甲基丁香酚监测桔小实蝇(B. Dorsalis)的研究类似[15-17]。通过监测发现儋州2014年3月~2015年2月和2015年3月~2016年2月南瓜实蝇发生的动态趋势从9月至翌年2月起有所差异,造成这种差异的原因可能由于这段时间气候变化差异较大和生境中的植被布局不同综合导致的。其中,2014年3月~2015年2月间南瓜实蝇的诱集数量12月份下降到最低,翌年1月份种群开始上升,而2015年3月~2016年2月间南瓜实蝇的诱集数量从12月份开始直线下降到翌年2月份,这两种趋势分别与两个时间段平均温度的趋势相同,这说明平均温度在这段时间内是影响南瓜实蝇种群发生的一个重要气象因子,这也与本研究中平均温度是影响南瓜实蝇数量动态最主要气象因子的结果一致。温度可以直接影响南瓜实蝇的发育、繁殖、种间竞争等[18-19],湿度可以通过空气的相对湿度、土壤的相对含水量等来影响南瓜实蝇种群的存活[20],而本研究除以上2个因素外发现气压和日照时数也是影响南瓜实蝇种群发生的重要气象因子,但具体如何影响还有待进一步验证。
本研究发现混合菜地、园林区以及混合果园诱集的南瓜实蝇数量显著多于在住宅区、农贸市场、苦瓜种植地诱集的数量,这一结果与瓜实蝇在6种生境下诱集的结果相似[21]。通过调查和研究发现南瓜实蝇和瓜实蝇的自然分布、寄主范围、种群增长能力等非常相似[19],这有可能是导致2种实蝇偏好相同的生境的原因。此外,本研究也说明南瓜实蝇的防治应该不仅仅局限在具有其寄主植物的生境。
综上,生境和气象因子显著影响了南瓜实蝇雄虫的数量动态。混合菜地、园林区以及混合果园诱集的南瓜实蝇雄虫数量显著多于住宅区、农贸市场、苦瓜种植地诱集的雄虫数;影响南瓜实蝇雄虫数量动态的气象因子有月平均气温、月平均气压、月平均相对湿度、月最小相对湿度和月日照时数,其中月平均温度是影响其数量动态的最关键气象因子。基于本研究的结果,南瓜实蝇的防控应不仅仅局限在具有其寄主植物的生境,而应根据监测的数据对其他生境的南瓜实蝇进行一并治理。
参考文献
[1] 王晓艳. 南瓜实蝇生态学及适生区预测研究[D]. 广州: 华南农业大学, 2009.
[2] 安坤鹏, 吴保锋, 申 科, 等. 南瓜实蝇特性及防治技术的研究进展[J]. 长江蔬菜, 2011(20): 7-13.
[3] Ohno S, Tamura Y, Haraguchi D, et al. First detection of the pest fruit fly, Bactrocera tau(Diptera: Tephritidae), in the field in Japan: Evidence of multiple invasions of Ishigaki Island and failure of colonization[J]. Applied Entomology and Zoology, 2008, 43(4): 541-546.
[4] White I M, Elson-Harris M M. Fruit flies of economic significance: their identification and bionomics[M]. Wallingford, United Kingdom: CAB International, 1992.
[5] 王泽乐, 刘映红, 江兴培, 等. 诱剂监测南亚果实蝇在重庆的分布与发生动态[J]. 西南农业大学学报(自然科学版), 2006, 28(2): 309-313.
[6] 付翠玉, 陈朝阳. 南瓜实蝇生活习性及其防治试验[J]. 植物保护, 1992, 49(1): 23-24.
[7] Singh S K, Kumar D, Ramamurthy V V. Biology of Bactrocera(Zeugodacus)tau(Walker)(Diptera: Tephritidae)[J]. Entomological Research, 2010, 40(5): 259-263.
[8] 方 焱, 李志红, 秦 萌, 等. 南亚果实蝇对我国南瓜产业的潜在经济损失评估[J]. 植物检疫, 2015, 29(3): 28-33.
[9] 马兴莉, 李志红, 胡学难, 等. 橘小实蝇、 瓜实蝇和南亚果实蝇对广东省造成的经济损失评估[J]. 植物检疫, 2013, 27(3): 50-56.
[10] 李小珍, 刘映红, 王泽乐. 检疫性害虫南亚果实蝇生物学及控制技术[J]. 植物保护, 2006, 32(6): 141-145.
[11] 张小亚, 陈国庆. 黄岩地区南亚果实蝇和瓜实蝇的发生代数预测及种群动态[J]. 浙江农业科学, 2012(11): 1 540-1 542.
[12] 周 湾, 蒋锡山, 邱志刚, 等. 南瓜实蝇在杭州地区的种群消长动态[J]. 浙江农业科学, 2010(2): 355-356.
[13] Hasyim A, de Kogel W. Population fluctuation of adult males of the fruit fly, Bactrocera tau Walker(Diptera: Tephritidae)in passion fruit orchards in relation to abiotic factors and sanitation[J]. Indonesian Journal of Agricultural Science, 2008, 9(1): 29-33.
[14] 刘丽红, 刘映红, 周 波, 等. 南亚实蝇在不同寄主上数量动态及危害研究[J]. 西南农业大学学报(自然科学版), 2005, 27(2): 176-179.
[15] 陈 鹏, 叶 辉, 刘建宏. 云南瑞丽桔小实蝇成虫种群数量变动及其影响因子分析[J]. 生态学报, 2006, 26(9): 2 801-2 809.
[16] 郑思宁. 不同生境中橘小实蝇种群动态及密度的差异[J]. 生态学报, 2013, 33(24): 7 699-7 706.
[17] Dhillon M K, Singh R, Naresh J S, et al. The melon fruit fly, Bactrocera cucurbitae: A review of its biology and management[J]. Journal of Insect Science, 2005, 5(40): 1-16.
[18] Zhou C, Wu K, Chen H, et al. Effect of temperature on the population growth of Bactrocera tau(Walker)(Dipt. Tephritidae)[J]. Journal of Applied Entomology, 1994, 117(1-5): 332-337.
[19] Shen K, Hu J, Wu B, et al. Competitive interactions between immature stages of Bactrocera cucurbitae(Coquillett)and Bactrocera tau(Walker)(Diptera: Tephritidae)under laboratory conditions[J]. Neotropical Entomology, 2014, 43(4): 335-343.
[20] Li X Z, Liu Y H, Wang J J, et al. Growth, development and water-loss dynamics in Bactrocera tau(Walker)(Diptera: Tephritidae)pupae exposed to determined humidity[J]. Pan-Pacific Entomologist, 2009, 85(3): 150-158.
[21] 李 磊, 马华博, 牛黎明, 等. 生境及气象因子对瓜实蝇雄成虫数量动态的影响[J]. 环境昆虫学报, 2016, 38(4): 766-770.
