小麦品种宁7840突变体农艺性状和赤霉病抗性解析
2016-05-27董晶晶李长成
钱 丹,骆 孟,董晶晶,李长成,李 磊,李 韬
(扬州大学江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心/教育部植物功能基因组学重点实验室/小麦研究中心,江苏扬州 225009)
小麦品种宁7840突变体农艺性状和赤霉病抗性解析
钱 丹,骆 孟,董晶晶,李长成,李 磊,李 韬
(扬州大学江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心/教育部植物功能基因组学重点实验室/小麦研究中心,江苏扬州 225009)
摘要:为了解突变体与野生型小麦的农艺性状、产量和抗性差异,以高抗赤霉病小麦品种宁7840及其23个甲磺酸乙酯(EMS) 诱导的纯合突变体(Mu4)作为材料,研究其分蘖数、株高、叶绿素含量、千粒重、小穗数、穗粒数及赤霉病抗性等12个指标的差异。结果表明,除叶长、分蘖数和小穗数在突变体与野生型之间无显著差异外,其余9个性状在野生型与部分突变体之间有显著差异。N8和N16两个突变体表现高感赤霉病;N16突变体的叶宽、SPAD值、千粒重和粒宽显著优于野生型,其株高显著低于野生型。赤霉病病小穗率与叶长显著负相关,叶长可作为筛选小麦赤霉病抗性品系的间接指标;叶宽和粒宽可作为筛选小麦产量性状的重要指标。基于12个性状的聚类分析,将24个小麦材料聚为3类,其中第I类在产量性状方面优于其他两类。
关键词:小麦;野生型;突变体;赤霉病抗性;农艺和产量性状
小麦遗传资源的多样性是小麦育种的重要基础[1]。随着小麦育种进程的加快,小麦种质资源主要集中在少数亲本上[2],导致大部分新育成品种的主要性状的变异幅度较狭窄。要继续提高小麦的品质特性,必须提高育种基础材料的遗传多样性[3]。突变体在植物遗传育种研究中,对快速挖掘重要基因并解析其功能具有重要的意义,不仅能够极大地丰富种质资源,对提高产量、增强抗性、改善品质和提高生产效率等均具有重要意义[4]。过去30多年来,育种家将杂交育种、物理和化学诱变相结合,并对育成品种的后代进行多代选择和鉴定,培育出了综合农艺性状良好、产量高、抗性与适应性较好的小麦品种[5]。
小麦的农艺性状能直观地反映小麦的状态[1],良好的株型是获得高产的重要因素。株高是影响小麦产量的主要因子之一[6],20 世纪 50~60 年代,矮秆基因被广泛研究和利用,并引发了第一次“绿色革命”,使小麦育种产生了阶段性变革,大幅提升了世界小麦产量,为解决世界粮食问题做出了巨大贡献[7]。叶片的长度、宽度及其空间分布对小麦产量也有重要影响。小麦有机物主要来源于光合作用[5],其积累量和叶片中叶绿素的含量关系密切,叶绿素含量可以通过SPAD值反应。小麦产量主要是由有效穗数、穗粒数和千粒重三个因素决定,有效穗数与分蘖的多少有关[8];有效穗数和穗粒数之间又相互制约,为了使小麦达到最大的产量,协调这三个性状关系是农业工作者的首要任务[9]。
小麦赤霉病是由禾谷镰刀菌 (Fusariumgraminearum) 引起的一种毁灭性真菌病害,主要在温暖潮湿和半潮湿麦区广泛发生。中国是世界上小麦赤霉病危害最大的国家,每年受害面积平均约750万hm2,占全国小麦总面积的 1/4[10],其中长江中下游麦区是小麦赤霉病重发区[11]。赤霉病大流行年份减产甚至达 50%[12],不仅严重影响小麦产量,使其品质恶化,种用价值降低[13],被感染的籽粒中含有的脱氧雪腐镰刀菌烯醇(DON)毒素,还可严重影响人类的健康[14]。选用抗病品种是克服赤霉病危害的最有效和环保的措施,赤霉病抗源的鉴定、筛选和创新是培育抗赤霉病品种的关键[15]。小麦品种对赤霉病的抗性主要表现为抗病害扩展,即降低病害自侵染点向邻近小穗扩展速度,保护穗轴不受伤害,减少病小穗数[16],目前大多数的抗性遗传分析和定位研究都建立在这类抗性基础上[17]。
本研究对宁7840及其突变体的主要农艺、产量性状以及赤霉病抗性进行鉴定,分析突变体与野生型在目标性状上的差异,以期筛选出小麦重要的种质资源,为后续的基因挖掘和育种提供参考依据。
1材料与方法
1.1试验材料
供试材料野生型宁7840(安徽11/Aurora//苏麦3号)是江苏省农科院培育的苏麦3号的衍生系,具有高抗赤霉病、条锈病、叶锈病和杆锈病的特点。23个突变体(Mu4)是宁7840经甲磺酸乙酯(EMS)诱导、连续自交和单株选择而来。宁7840的野生型和23个突变体分别命名为N1(WT)和N2~N24。
1.2试验方法
1.2.1材料的种植
试验于2013年10月至2014年5月在扬州大学教学试验田种植,前茬作物为水稻,肥力中等。每个品系等距种两行,行头种植野生型作为对照,人工点播以保证苗间距。
1.2.2赤霉病抗性鉴定
赤霉病接种及其鉴定方法参照Li等[17]的方法。在小麦扬花期每品系选取约15个穗子进行赤霉病接种。所用菌株为毒性较强的Fg 65菌株。将约20 μL菌液注射到倒数第5小穗小花中,贴上防水胶带,注明接种日期。接种21 d后进行鉴定,计算病小穗率,病小穗率=病小穗数/总小穗数。
小麦成熟后,将15株接种赤霉病菌的小麦分别装在纸袋中,标明行号;单独脱粒,明显发白或皱缩的籽粒计为病粒,籽粒感染率(病粒率)=病粒数/总粒数
1.2.3农艺性状鉴定
花后15 d,野生型和突变体每个品系随机挑选5株植株,分别测量株高、旗叶的长、宽和叶绿素含量;收获时进行考种,统计或检测有效分蘖数、小穗数、穗粒数、千粒重、籽粒长和宽等。叶绿素含量是用SPAD仪测定旗叶上、中、下三部分的SPAD值,取平均值。
1.2.4数据处理
采用Excel 2010进行进行数据初处理,采用SPSS 21进行多重比较;Pearson相关分析采用Sigma Plot 10.0软件进行;聚类分析(采用最小平方和为距离的方法)采用MATLAB R2012b进行。
2结果与分析
2.1宁7840及其突变体的赤霉病抗性
赤霉病的扩展抗性一般用病小穗率表示。宁7840野生型的病小穗率变幅为0.09~0.12,平均为0.10;23个突变体的病小穗率变幅为0.04~0.66,平均为0.24。各品系的病小穗率间有不同程度差异。其中N8和N16两个突变体的病小穗率均显著高于野生型。
宁7840野生型的病粒率变幅为0.19~0.25,平均为0.22;突变体的病粒率变幅为0.22~1.0,平均为0.78。各品系的赤霉病籽粒抗性间存在不同程度差异,其中N3、N8、N16、N20、N21、N23和N24等7个品系的病粒率均显著高于野生型。
2.2宁7840及其突变体的叶长、叶宽和叶绿素含量
宁7840野生型叶长变幅为19.00~30.00 cm,平均为24.70 cm;突变体叶长平均为26.28 cm,变幅为14.67~37.80 cm。各品系的叶长有不同程度差异;所有突变体与野生型间无显著差异(表2)。
宁7840野生型叶宽变幅为1.20~1.80 cm,平均为1.54 cm;突变体叶宽变幅为1.16~2.30 cm,平均为1.72 cm。各品系的叶宽有不同程度差异,其中突变系N17、N18、N21、N16的叶宽显著大于野生型,分别比野生型宽0.48、0.48、0.5、0.76 cm。
宁7840野生型扬花期期叶的SPAD值变幅为12.80~26.20,平均为21.62。突变体SPAD值变幅为5.64~48.70,平均为25.91。其中N16的SPAD值显著高于野生型,比野生型高27.08。
2.3宁7840及其突变体的株高、有效分蘖数和小穗数
宁7840野生型株高变幅为97.00~105.00 cm,平均为99.00 cm;突变体株高变幅为62.67~103.1 cm,平均为87.34 cm。其中N7、N3、N11、N8、N18、N10、N12、N16、N9和N4等10个突变体的株高均显著低于野生型,比野生型分别低38.16、33.66、27.66、27.16、26、23.66、22.83、20.0、13.83、13.16 cm。
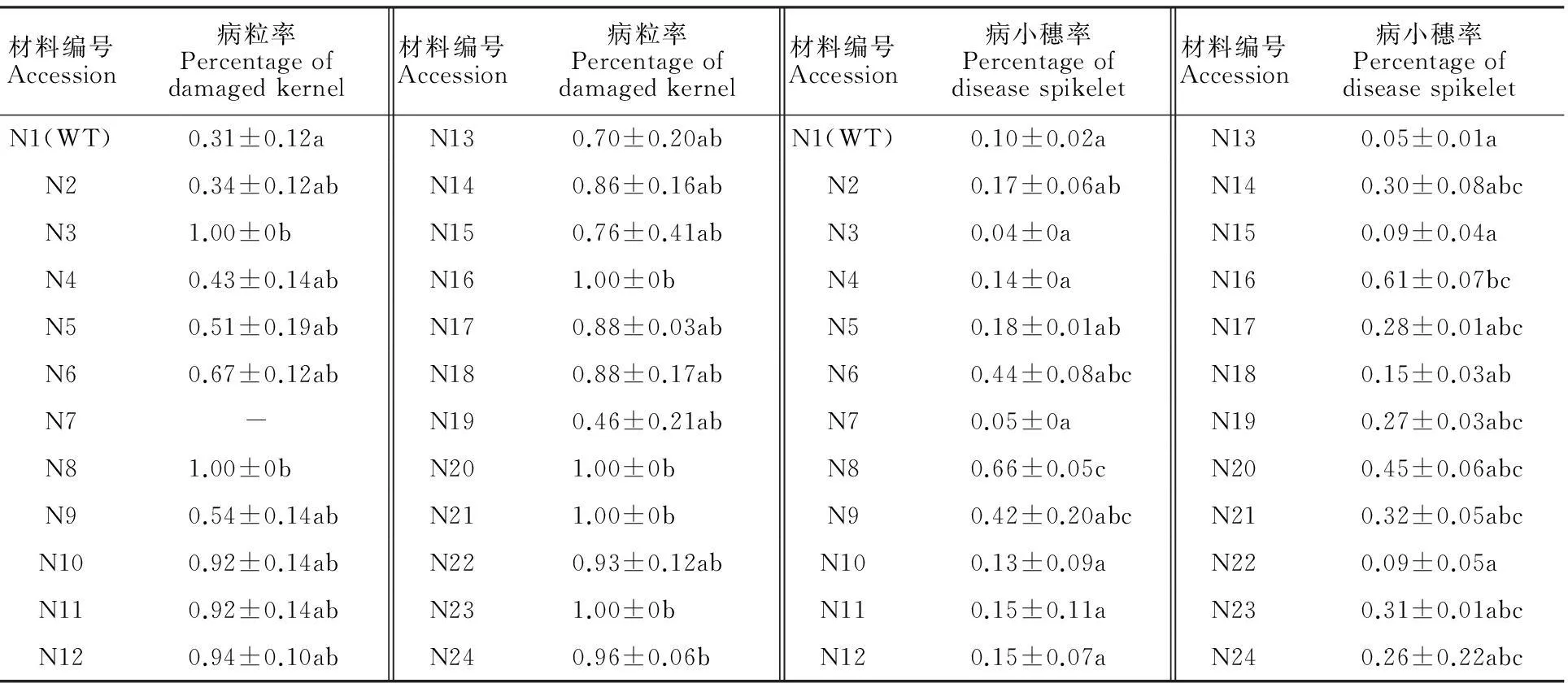
表1 宁7840野生型及其突变体对赤霉病的抗性
数据后不同字母表示材料间差异显著(P<0.05)。下同
Data followed by different letters mean significant differences among different lines(P<0.05). The same as following tables
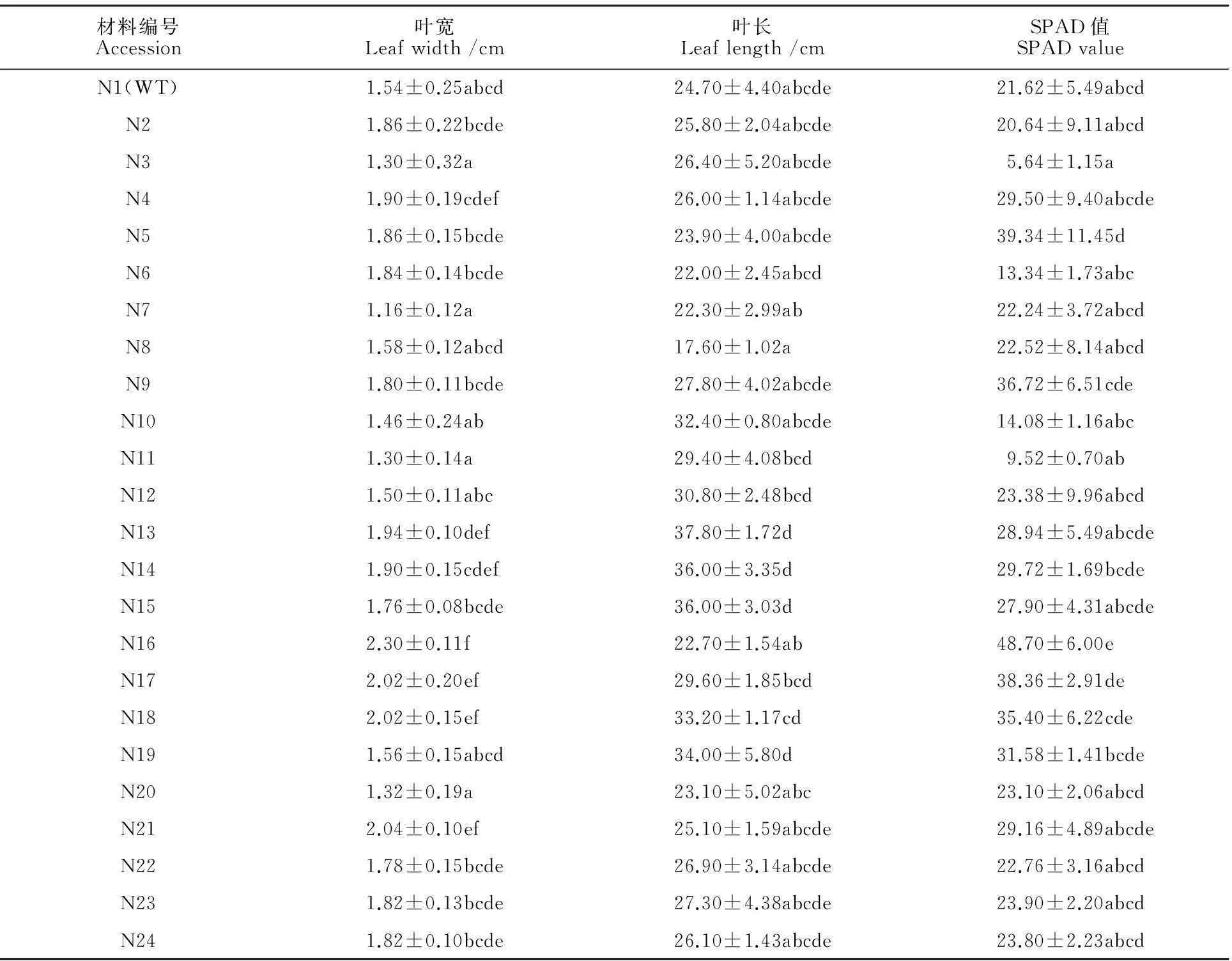
表2 宁7840野生型及其突变体叶宽、叶长和叶绿素含量
宁7840野生型的分蘖数变幅为3.00~7.00个,平均为4.80个;突变体的分蘖数变幅为2.60~6.80个,平均为4.59个。所有突变体与野生型间无显著差异。
野生型的小穗数变幅为20.00~22.00,平均为20.85个;突变体小穗数变幅为14.80~20.50个,平均为18.0个。N13的小穗数显著低于野生型,其他突变体与野生型的小穗数间无显著差异(表3)。
2.4宁7840及其突变体的穗粒数、千粒重、粒长和粒宽
宁7840野生型的穗粒数变幅为42.00~55.00粒,平均为48.50粒;突变体的穗粒数变幅为19.50~52.50粒,平均为 36.00粒。其中N10、N15、N11、N12、N8等5个突变体与野生型的穗粒数存在显著差异(表4),比野生型分别低29.13、27.15、23.46、22.82和21.94粒,其他突变体与野生型无显著差异。
宁7840野生型千粒重变幅为31.00~33.60 g,平均为32.30 g,其突变体千粒重变幅为12.80~49.70 g,平均为32.80 g。其中N3、N7、N2、N22、N4、N16等5个突变体与野生型间存在显著差异(表4)。N3、N7突变体千粒重显著低于野生型,比野生型低19.40、7.40 g,其余4个突变体千粒重均高于野生型,比野生型分别高7.10、7.30、8.40、17.40 g。
宁7840野生型的粒长变幅为0.63~0.64 cm,平均为0.64 cm,其突变体粒长变幅为0.54~0.72 cm,平均为0.66 cm。其中N3突变体的粒长显著小于野生型,二者相差0.10cm(表4)。宁7840野生型的粒宽为0.31 cm,其突变体的粒宽变幅为0.23~0.40 cm,平均为0.32 cm。其中N3粒宽显著小于野生型,二者相差0.80 cm,而N16粒宽则显著高于野生型,与野生型差0.85 cm。
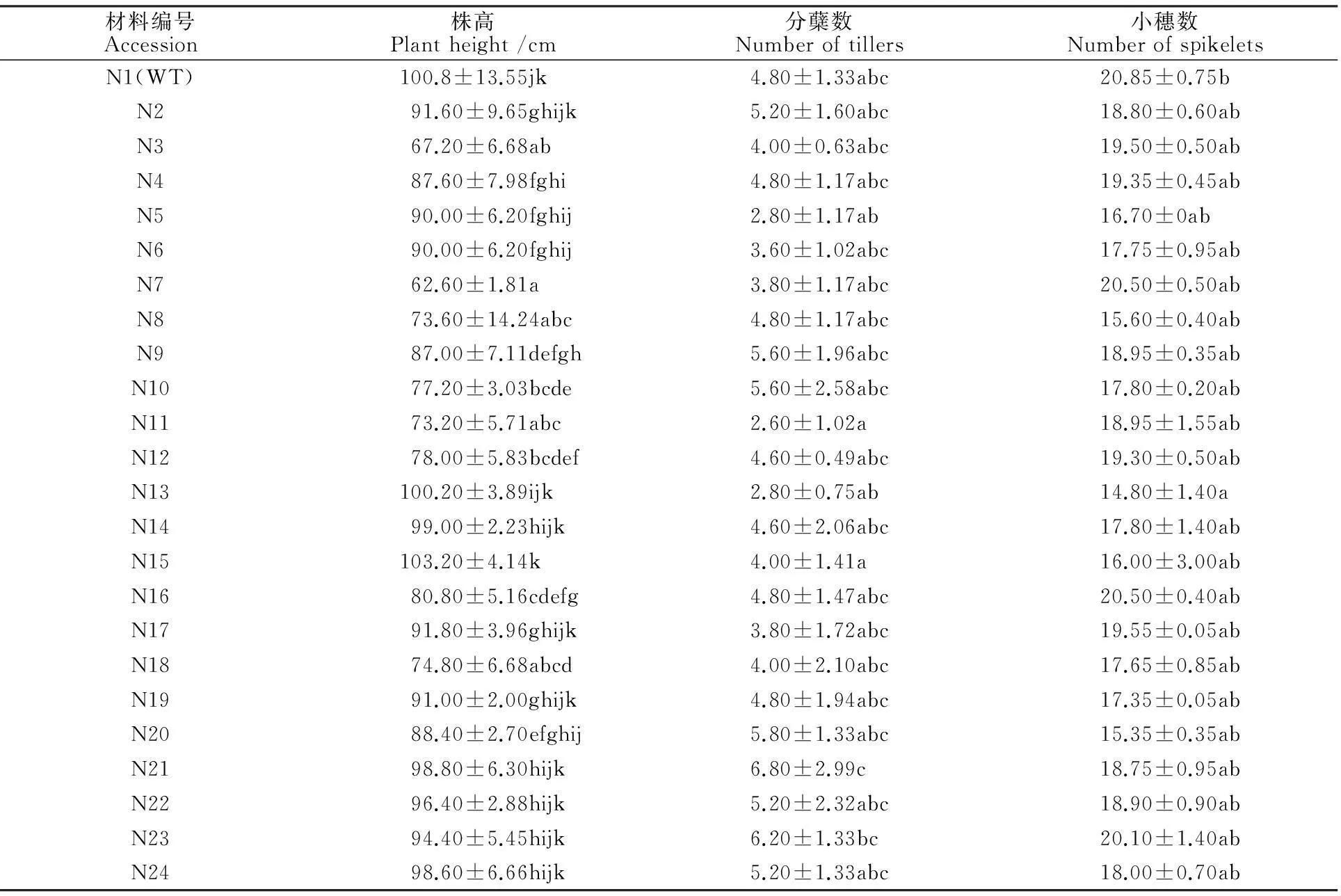
表3 宁7840野生型及其突变体的株高、分蘖数和每穗小穗数
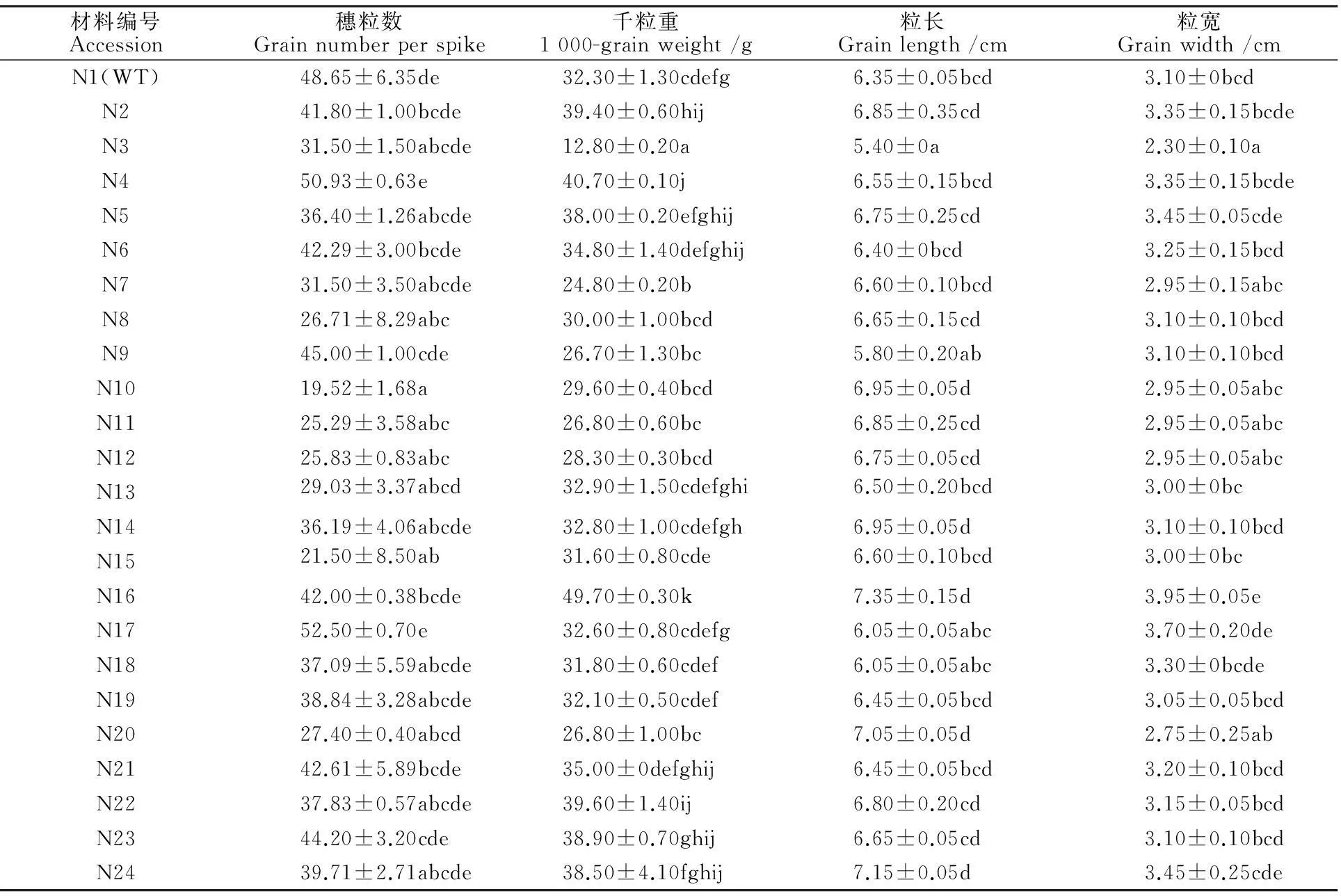
表4 宁7840野生型及其突变体的穗粒数、千粒重、粒长和粒宽
2.5小麦各性状的相关性分析
相关性分析表明(表5),小麦叶长与病小穗率显著负相关(P<0.05);叶宽与SPAD值、穗粒数、千粒重、粒宽呈极显著正相关(P<0.01),与株高显著正相关;SPAD值与穗粒数显著正相关,而与千粒重和粒宽极显著正相关;穗粒数与小穗数和粒宽呈极显著正相关,与千粒重显著正相关;株高仅与千粒重显著正相关;千粒重与粒长和粒宽极显著正相关。
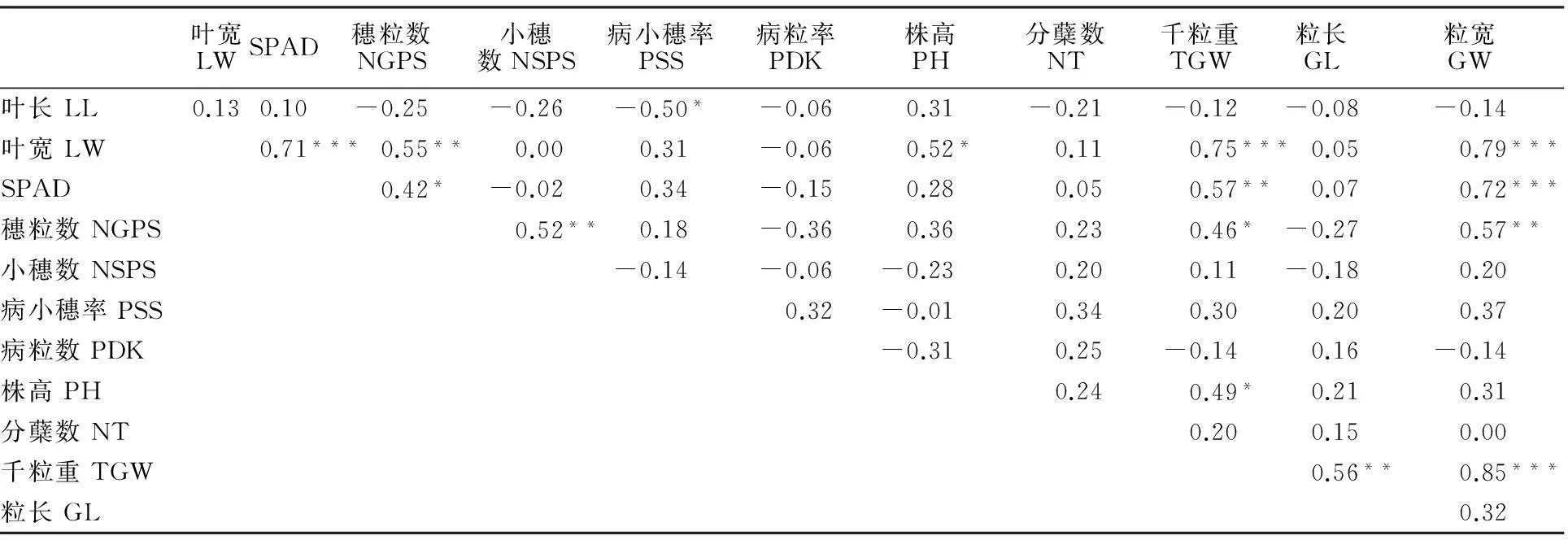
表5 宁7840及其突变体各个性状间的相关性
***、**和*分别表示在0.001、0.01和0.05水平上显著。*** indicates significant difference at 0.1% level ,** at 1% level and * at 5% level.LW:Leaf width; NGPS:Number of grains per spike; NSPS: Number of spikelets per spike; PSS: Proportion of symptomatic spikelets; PDK: Proportion of damaged kernel;PH: Plant height; NT: Number of tillers; TGW: 1000- grain weight; GL: Grain length; GW: Grain weight
2.6聚类分析
对宁7840及其突变体的12个农艺性状进行主成分分析,再提取前三个主成分(累积贡献率66.5%),采用最小组内平方和计算距离的动态聚类方法,将野生型及其突变体聚为3类(图1),第Ⅰ类包括11个品系,分别是N1(野生型)、N2、N4、N6、N9、N16、N17、N21、N22、N23、N24;第Ⅱ类包括6个品系,分别是N5、N13、N14、N15、N18、N19;第Ⅲ类包括7个品系,分别是N3、N7、N8、N10、N11、N12、N20。对12个不同性状在3类间运用Tukey法进行多重比较,除了病小穗率、病粒率、分蘖数和粒长在类间无显著差异,其余8个性状至少在两类间有显著差异(P<0.05)(表6)。在叶宽、粒宽、千粒重、分蘖数、小穗数和穗粒数等性状方面,第Ⅰ类品系的表现优于其他两类。
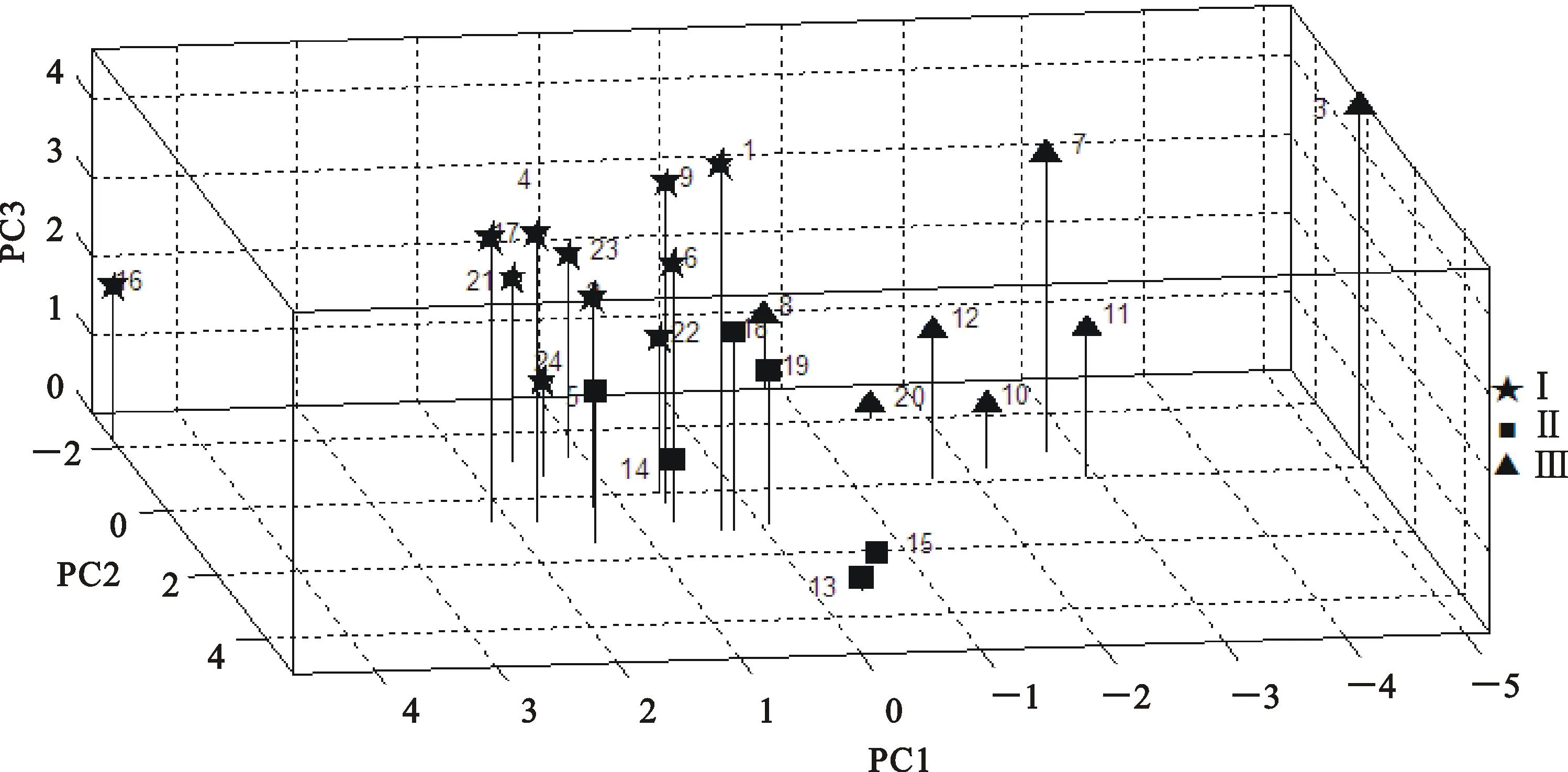
图1 宁7840野生型和突变体的聚类图
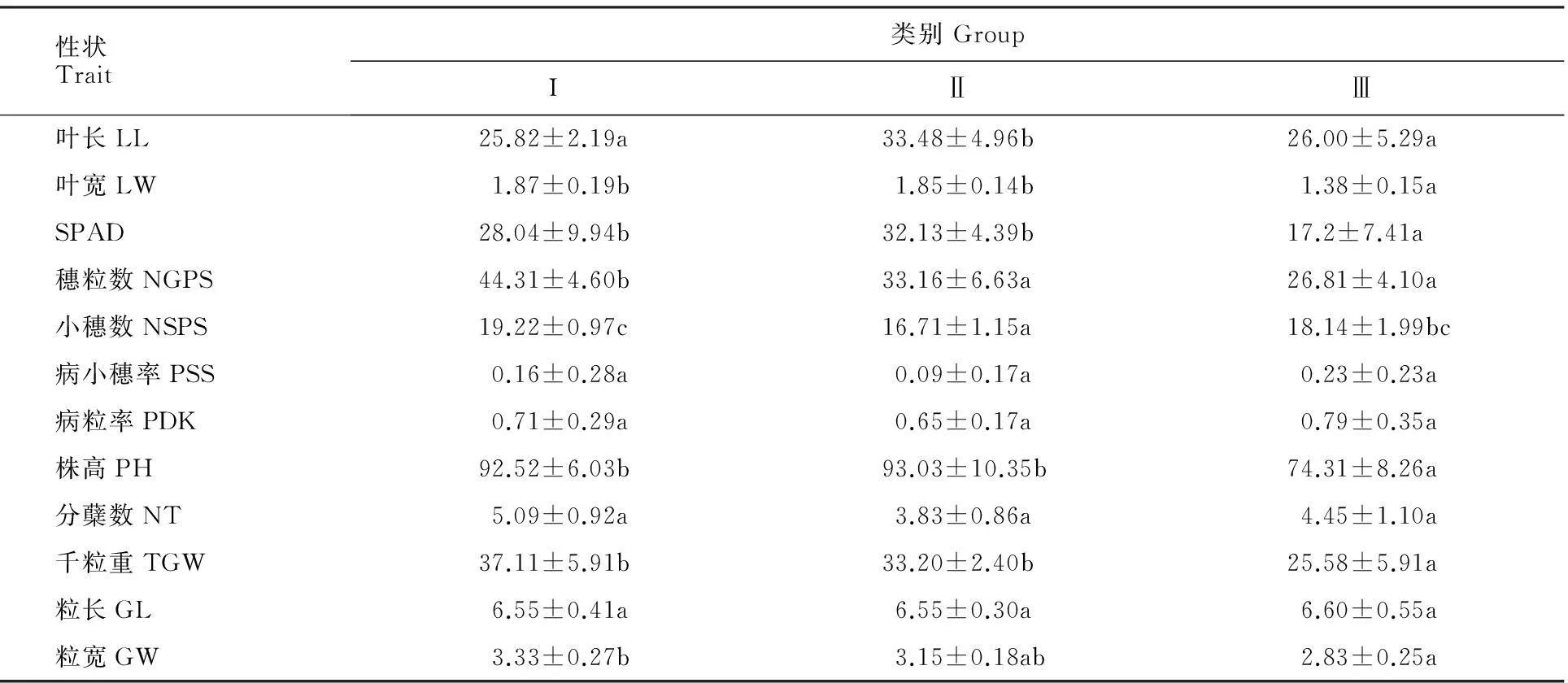
性状Trait类别GroupIⅡⅢ叶长LL25.82±2.19a33.48±4.96b26.00±5.29a叶宽LW1.87±0.19b1.85±0.14b1.38±0.15aSPAD28.04±9.94b32.13±4.39b17.2±7.41a穗粒数NGPS44.31±4.60b33.16±6.63a26.81±4.10a小穗数NSPS19.22±0.97c16.71±1.15a18.14±1.99bc病小穗率PSS0.16±0.28a0.09±0.17a0.23±0.23a病粒率PDK0.71±0.29a0.65±0.17a0.79±0.35a株高PH92.52±6.03b93.03±10.35b74.31±8.26a分蘖数NT5.09±0.92a3.83±0.86a4.45±1.10a千粒重TGW37.11±5.91b33.20±2.40b25.58±5.91a粒长GL6.55±0.41a6.55±0.30a6.60±0.55a粒宽GW3.33±0.27b3.15±0.18ab2.83±0.25a
同行数据后不同字母表示类别之间差异在0.05水平显著
Data followed by different letters in the same line mean significant difference at 0.05 level
3讨 论
3.1突变体与野生型性状的差异
本研究检测并分析了宁7840及其突变体的多个性状后发现,宁7840品系中,N16突变体在7个农艺性状上与野生型有显著差异,与野生型相比,株高明显降低,叶片显著变宽,SPAD值、千粒重和粒宽也显著增加,抗倒伏能力以及总体的农艺性状均有所提高,但对赤霉病由高抗变成了高感。因此该突变体是研究农艺性状、产量和赤霉病抗性的理想遗传材料。从育种角度讲,如果该突变体能够保持宁7840的高抗赤霉病的特性,就可能育出矮杆抗倒,株型优良,高产抗病的优异小麦品种,但是农艺、产量和抗性三者是否具有一因多效则需要在大的分离群体中进行验证。
N3突变体和N8突变体在株高、粒长和粒宽以及抗性方面均变差,虽然从育种角度没有明显意义,但是遗传上可以对其研究,挖掘农艺性状相关的基因位点。
聚类分析发现N22、N23、N24与野生型聚在同一类,但其对赤霉病的籽粒抗性变差,因此这三个材料是研究赤霉病籽粒抗性的理想遗传材料。
3.2农艺性状间的相互关系
许多研究表明,小麦病小穗率与株高呈负相关,即株高越高,抗病性越好。本研究发现,病小穗率与株高的相关性远未达显著水平,而病小穗率与叶长呈显著负相关(P<0.05),表明叶长越长,小麦对赤霉病的扩展抗性越好,可为育种家培育赤霉病抗病品种提供一定的参考信息。叶宽与穗粒数、千粒重、粒宽极显著相关,因此可作为选择产量性状的一个重要指标。粒宽与千粒重、穗粒数极显著正相关,表明粒宽也是选择产量性状的一个重要指标。控制显著相关性状的基因位点是一因多效或紧密连锁,有待于在相关突变体中进一步分析。
本试验中的突变体,可为小麦重要性状的遗传以及不同性状间的遗传关系研究提供重要材料,部分农艺性状显著改良的突变体可为育种提供重要的资源。本试验对各个突变体采用了基于最小组内平方和距离的动态聚类分析方法,可综合评价野生型与突变体,以及突变体本身间的差异,可以综合判断材料间的相似情况,同时也是本研究的另一重要创新点。
参考文献:
[1]王林海,王晓伟,詹克慧,等.黄淮麦区部分小麦种质资源农艺性状的聚类分析[J].江苏农学通报,2008,24(4):186-191.
Wang L H,Wang X W,Zhan K H,etal.Cluster analysis of some wheat germplasms in Huang-huai area based on agronomic traits [J].ChineseAgriculturalScienceBulletin,2008,24(4):186-191.
[2]王翠玲,张灿军,王书子.河南省优质小麦系谱追溯及遗传改良分析[J].中国农学通报,2002,18(2):80-82.
Wang C L,Zhang C J,Wang S Z.Pedigree analysis and genetic improvement of wheat with elite qualityin Henan province [J].ChineseAgriculturalScienceBulletin,2002,18(2):80-82.
[3]Rasmusson D C,Phillips R L.Plant breeding progress and genetic diversity from denovo variation and elevated epistasis [J].CropScience,1997,37:330-336.
[4]杨 辉,李金秀,张明辉,等.小麦突变体的创造及育种利用研究[J].农业科技通讯,2006(3):18-19.
Yang H,Li J X,Zhang M H,etal.The creation of the wheat mutants and the breeding [J].BulletionofAgricuturalScienceandTechnology,2006(3):18-19.
[5]海 燕,何 宁,康明辉,等.小麦主要农艺性状的遗传分析[J].中国农学通报,2008,24(6):168-171.
Hai Y,He N,Kang M H ,etal.Genetic analysis of agronomic traits in wheat [J].ChineseAgriculturalScienceBulletin,2008,24(6):168-171.
[6]何中虎,张树榛.小麦矮秆育种中性状间关系的多元分析[J].华北农学报,1992,7(1):1-7.
He Z H,Zhang S Z.Relations between wheat stem traits in breeding of multivariate analysis [J].ActaAgriculturaeBoreali-Sinica,1992,7(1):1-7.
[7]陈 亮.矮秆基因 Rht12对小麦重要农艺性状的遗传效应及新矮秆突变体的筛选[D].杨凌:西北农林科技大学,2014.
Chen L.Genetic effects of dwarfing gene Rht12 on the improtant agronomic traits of common bread wheat and screening of new dwarf mutants [D].Yangling:Northwest A&F University,2014.
[8]张 晶,张定一,王姣爱,等.小麦单株有效分蘖数与农艺性状的相关性研究[J].山西农业科学,2009,37(6):17-19.
Zhang J,Zhang D Y,Wang J A,etal.The dependence study of the effective tillers per plant andagronomic characters in wheat [J].JournalofShanxiAgriculturalSciences,2009,37(6):17-19.
[9]毕晓静,史秀秀,马守才,等.小麦农艺性状的主基因+多基因遗传分析[J].麦类作物学报,2013,33(4):630-634.
Bi X J,Shi X X,Ma S C,etal.Genetic analysis of agronomic traits related to yield based on major gene plus polygene model in wheat [J].JournalofTriticeaeCrops,2013,33(4):630-634.
[10]方兴洲.小麦赤霉病研究进展[J].现代农业科技,2014(23):134-135.
Fang X Z.Research progress on wheat head scab [J].ModernAgriculturalSciencesandTechnology,2014(23):134-135.
[11]姚金保,任丽娟,张平平,等.小麦赤霉病的抗性遗传分析[J].麦类作物学报,2011,31(2):370-375.
Yao J B,Ren L J,Zhang P P,etal.Genetic analysis of resistance to fusarium head blight in wheat [J].JournalofTriticeaeCrops,2011,31(2):370-375.
[12]林壮森,张 焜,赵肃清,等.食品中生物毒素的 ELISA分析方法研究进展[J].食品科学,2009,30(2):281-283.
Lin Z S,ZhangK,Zhao S Q,etal.Research progress on ELISA method for analysis of biotoxins in foodstuffs [J].FoodScience,2009,30(2):281-283.
[13]陆维忠,程顺和,王裕中.小麦赤霉病研究[M].北京:北京科学出版社,2001:2.
Lu W Z,Cheng S H,Wang Y Z.Studies on Fusarium Head Blight of Wheat [M].Beijing:Beijing Science Press,2001:2.
[14]李小勋,顾乃杰,张玉松.小麦抗赤霉病育种研究进展[J].作物研究,2010,25(3):264-268.
Lei X X,Gu N J,Zhang Y S,etal.Advances in wheat breeding on scab resistance [J].CropResearch,2010,25(3):264-268.
[15]程顺和,张 勇,别同德,等.中国小麦赤霉病的危害及抗性遗传改良[J].江苏农业学报,2012,28(5):938-942.
Chen S H,Zhang Y,Bie T D,etal.Damage of wheat Fusarium head blight (FHB) epidemcs and genetic improvement of wheat for scab resistance in China [J].JiangsuAgriculturalSciences,2012,28(5):938-942.
[16]刘惕若,薛国兴,张匀华.小麦品种对赤霉病的抗性[J].黑龙江八一农垦大学学报,1989(1):9-18.
Liu X R,Xue G X,Zhang Y H.Resistance to Fusarium head blight of wheat varieties [J].JournalofHeilongjiangAugustFirstLandandReclamationUniversity,1989(1):9-18.
[17]Li T,Bai G,Wu S and Gu S.Quantitative trait loci for resistance to fusarium head blight in a Chinese wheat landrace Haiyanzhong [J].TheoreticalandAppliedGenetics,2011,122:1497-1502.
Evaluation of Agronomic Traits and Resistance to Fusarium Head Blight of Ning 7840 and Its Mutants
QIAN Dan,LUO Meng,DONG Jingjing,LI Changcheng,LI Lei,LI Tao
(Jiangsu Provincial Key Laboratory of Crop Genetics and Physiology/Co-Innovation Center for Modern Production Technology of Grain Crops/Key Laboratory of Plant Functional Genomics of Ministry of Education/wheat Rearch Center;Yangzhou University,Yangzhou,Jiangsu 225009,China)
Abstract:To understand the alterations in agronomic traits,yield traits and Fusarium head blight (FHB) resistance between the wild type Ning 7840 and its 23 ethyl methanesulfonate (EMS)-induced mutants (Mu4),twelve traits,such as number of tillers per plant,plant height,chlorophyll content (SPAD value),1 000-grain weight,number of spikelet,number of grain per spike,and FHB resistance,etc. were investigated. Significant differences between the wild type and the mutants were found in nine out of twelve traits except for flag leaf length,number of spikelet,and number of tiller per plant. Two mutants,N8 and N16,became susceptible to FHB. However,the mutant N16 was significantly different from and superior to the wild type in flag leaf width,SPAD value,1 000-grain weight,and grain width. The plant height of N16 was significantly shorter than that of the wild type. The significant negative correlation between FHB severity and flag leaf length suggested that the latter can be an indicator trait for prediction of FHB severity of a wheat variety.Increase of flag leaf width and grain width could be essential for achieving high yield in wheat breeding. These 24 accessions were classified into three distinct groups based on all the twelve traits,and in general the group I was superior to the other two groups in yield-related traits.
Key words:Wheat; Wild type; Mutant; Resistance to Fusarium head blight; Agronomic and yield traits
中图分类号:S512.1;S435.121
文献标识码:A
文章编号:1009-1041(2016)02-0243-08
通讯作者:李 韬(E-mail:taoli@yzu.edu.cn)
基金项目:国家科技重大专项子课题(2012ZX08009003-004);国家自然科学基金项目(31171537,31270704);江苏省高校自然科学基金重大项目(12KJA210002);江苏省高校优势学科建设工程资助项目(PAPD)
收稿日期:2015-07-23修回日期:2015-10-13
网络出版时间:2016-01-26
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160126.1946.032.html
第一作者E-mail:572652817@qq.com; 1946220133@qq.com(骆 孟与第一作者同等贡献)
