自噬对体外醛固酮诱导的大鼠系膜细胞凋亡的影响
2016-05-24曲高婷张爱青施会敏甘卫华
曲高婷,张爱青,施会敏,甘卫华
(南京医科大学第二附属医院 儿科,江苏 南京 210003)
·基础医学·
自噬对体外醛固酮诱导的大鼠系膜细胞凋亡的影响
曲高婷,张爱青,施会敏,甘卫华
(南京医科大学第二附属医院儿科,江苏南京210003)
【摘要】目的:通过体外研究醛固酮(ALD)和蟾蜍灵(Bufalin)作用于大鼠系膜细胞后自噬与凋亡的变化,探讨自噬在醛固酮诱导系膜细胞凋亡中的作用,进而说明醛固酮参与肾损伤的可能机制。 方法:体外培养大鼠系膜细胞,分别应用醛固酮、蟾蜍灵、氯喹(CQ)作用于系膜细胞,实验分为a:对照组、b:ALD组(10-6mol/L 醛固酮),c:Bufalin组(10-8mol/L 蟾蜍灵),d:ALD+Bufalin组(10-6mol/L 醛固酮+10-8mol/L 蟾蜍灵),e:ALD+CQ组(10-6mol/L 醛固酮+10 μmol /L氯喹),f:ALD+Bufalin+CQ组(10-6mol/L 醛固酮+10-8mol/L 蟾蜍灵+10 μmol /L氯喹);应用透射电镜观察各组大鼠系膜细胞的自噬水平;应用Western Blot检测各组自噬标志蛋白P62、LC3表达的变化;应用An- nexin V /PI 双染流式细胞术检测各组大鼠系膜细胞凋亡的变化。 结果:透射电镜下:a组和c组未见典型自噬体,b组可见典型自噬体,d组自噬体数目小于b组,e组、f组自噬体数目均明显高于各组;Western Blot检测P62、LC3蛋白表达结果显示:c组表达量与a组基本相同,b组较a、c组升高(P<0.05);d组较b组下降(P<0.05);e组、f组表达量基本相当,且显著高于各组(P<0.05);流式细胞仪检测凋亡结果显示:c组细胞凋亡率(46.23±5.83)%与a组(46.10±3.49)%基本相同,b组细胞凋亡率(76.47±3.03)%较a组明显升高(P<0.05),d组(61.73±2.76)%、e组(61.03±3.62)%、f组细胞凋亡率(60.50±1.95)%基本相同,较a、c组明显升高(P<0.05),但较b组下降(P<0.05)。 结论:醛固酮可同时诱导系膜细胞自噬及凋亡增加,蟾蜍灵及氯喹可以抑制醛固酮对自噬及细胞凋亡的诱发作用,提示自噬可促进醛固酮诱发的系膜细胞凋亡,醛固酮可能通过增加自噬从而促进系膜细胞凋亡。
【关键词】醛固酮;蟾蜍灵;系膜细胞;自噬;凋亡
【DOI】10.3969/j.issn.1002-0217.2016.02.003
醛固酮(aldosterone,ALD)是引起肾损伤的重要物质,它可通过作用于肾脏固有细胞发挥作用。系膜细胞是肾小球内最活跃的固有细胞,其数量、形态及位置的相对稳定直接影响着肾脏正常结构和功能的维持,其受损可促进肾脏疾病的发生发展。研究表明醛固酮可直接诱导系膜细胞凋亡,可能是醛固酮引起的肾损伤的机制之一[1],但醛固酮如何调节细胞凋亡的机制尚不清楚。近年来,多项研究表明细胞自噬与多种肾脏疾病的发生发展密切相关[2],且自噬与细胞凋亡间有着密不可分且错综复杂的关系,自噬既可诱导凋亡,又可拮抗凋亡的发生[3]。因此,本研究通过观察醛固酮,蟾蜍灵及自噬抑制剂氯喹分组作用于大鼠系膜细胞后,细胞自噬与凋亡的变化,探讨系膜细胞自噬与凋亡间的关系,并为临床应用醛固酮受体拮抗剂和蟾蜍灵治疗肾脏疾病提供相应实验基础和理论依据。
1材料与方法
1.1主要实验材料和仪器正常大鼠肾小球系膜细胞株[中国典型培养物保藏中心(CCTCC)];胎牛血清、0.25% 含EDTA的胰蛋白酶、DMEM培养液(Gibco,美国);醛固酮、蟾蜍灵、氯喹(Sigma,美国);β-actin抗体(Bioworld,美国);P62抗体、LC3抗体(Sigma,美国);HRP-标记的羊抗兔二抗、HRP-标记的羊抗小鼠二抗(CST,美国);蛋白Marker(Thermo,加拿大);透射电子显微镜(JEOL,日本);流式细胞仪(BD,法国)。
1.2实验方法
1.2.1细胞培养大鼠肾小球系膜细胞生长于含10%胎牛血清的DMEM 培养液(含青霉素 100 U /mL,链霉素100 μg /mL ) 中,置于37 °C、 5% CO2培养箱内培养,每 2 天换液,当细胞长至70%~80% 融合时用0.25%的胰酶消化传代,取第 4~6 代细胞用于实验。
1.2.2实验分组将处于对数生长期的细胞用胰酶消化后,以 2×105个/孔密度接种于6孔板培养,当细胞长至 70%~80%融合时换用无血清培养液继续培养24 h,使细胞生长同步化。实验分组为a组(对照组);b组(醛固酮组:醛固酮10-6mol/L,根据既往研究确定[1]);c组(Bufalin组:蟾蜍灵10-8mol/L,根据既往研究确定[4]);d组(ALD+Bufalin组:醛固酮10-6mol /L+蟾蜍灵10-8mol/L);e组(ALD+CQ组:醛固酮10-6mol/L+氯喹10 μmol /L);f组(ALD+Bufalin+CQ组:醛固酮10-6mol/L+蟾蜍灵10-6mol /L+氯喹10 μmol /L)。
1.2.3透射电子显微镜术细胞生长同步化后,根据不同分组处理细胞。24 h 后PBS 洗涤细胞3 次,胰酶消化,1500 r/min 离心10 min,使细胞成团,小心吸去上清,加入2.5%戊二醛固定2 h,送电镜室包埋、切片、染色,透射电子显微镜下观察自噬体的存在。
1.2.4Western Blot 检测自噬相关蛋白P62、LC3表达细胞生长同步化后,根据不同分组处理细胞。24 h 后PBS洗涤细胞3 次,按100∶1 加入蛋白裂解液和蛋白酶抑制剂(100 μL+1 μL),并于冰上裂解30 min,4℃,15 000 r/min 离心10 min,取上清,加入上样缓冲液,100℃水浴变性10 min。根据二喹啉甲酸(BCA)蛋白浓度测定试剂盒说明书测定蛋白浓度。每组按30 μg 蛋白上样量进行SDS-PAGE 电泳,将电泳分离的样本转移到固相载体PVDF膜上,室温下5%脱脂奶粉封闭2 h,加入LC3、P62 及β-actin 一抗(工作液浓度1∶1000)于4℃孵育过夜,1×TBST漂洗一抗,加入相应的HRP 标记的二抗(工作液浓度1∶5000)室温孵育2 h,1×TBST 漂洗后ECL 显色,凝胶成像系统曝光并显影。以β-actin 作为内参,进行灰度值分析,计算目的蛋白的相对表达量。
1.2.5An-nexin V/PI 双染流式细胞术检测细胞凋亡根据 An-nexin V-FITC /PI 凋亡检测试剂盒说明操作。 细胞生长同步化后,根据不同分组处理细胞。24 h后胰酶消化后收集细胞,用预冷的 PBS 洗涤细胞 2 次并离心,100 μL 预冷 binding buffer 重悬细胞,加入 5 μL An-nexin V-FITC 和 5 μL PI 混匀,避光室温反应10 min,加入400 μL binding buffer 终止反应,筛网过滤后用流式细胞仪检测细胞凋亡率。 BD Accuri C6 软件分析细胞凋亡。
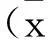
2结果
2.1各组大鼠系膜细胞自噬情况比较透射电子显微镜下观察,对照组、Bufalin组未见典型的双层膜包裹细胞浆及细胞器的自噬体,ALD组可见典型自噬体,ALD+Bufalin组见自噬体但数目小于ALD组,ALD+CQ组见自噬体且数目明显高于ALD组,ALD+Bufalin+CQ组见自噬体且数目高于ALD+Bufalin组但低于ALD+CQ组。见图1。

a.对照组;b.ALD组;c.Bufalin组;d.ALD+Bufalin组;e.ALD+CQ组;f.ALD+Bufalin+CQ组。
图1大鼠系膜细胞自噬的变化
2.2各组自噬相关蛋白P62和LC3表达比较Western Blot结果显示,各组间自噬相关蛋白P62存在显著差异(F=480.11,P=0.000)。b组P62蛋白表达较a组明显升高(P=0.000);c组P62蛋白表达较a组无明显变化,d组P62蛋白表达较b组明显降低(P=0.000),d组P62蛋白表达较c组显著升高(P=0.000),e组P62蛋白表达较b组显著升高(P=0.000),f组P62蛋白表达较e组无明显变化,f组P62蛋白表达较d组显著升高(P=0.000,图2A、B)。
Western Blot结果显示,各组间自噬相关蛋白LC3存在显著差异(F=179.53,P=0.000)。b组LC3蛋白表达较a组明显升高(P=0.000);c组LC3蛋白表达较a组无明显变化,d组LC3蛋白表达较b组明显降低(P=0.000),d组LC3蛋白表达较c组显著升高(P=0.000),e组LC3蛋白表达较b组显著升高(P=0.000),f组LC3蛋白表达较e组无明显变化,f组LC3蛋白表达较d组显著升高(P=0.000,图2A、C)。

a.对照组;b.ALD组;c.Bufalin组;d.ALD+Bufalin组;e.ALD+CQ组;f.ALD+Bufalin+CQ组;*P<0.05vsa组;#P<0.05vsb组;△P<0.05vsc组;&P<0.05vsd组。
图2自噬相关蛋白P62和LC3表达的变化
2.3各组大鼠系膜细胞凋亡的变化流式细胞仪检测细胞凋亡结果显示,a组细胞凋亡率(46.10±3.49)%,b组细胞凋亡率(76.47±3.03)%,c组细胞凋亡率(46.23±5.83)%;d组细胞凋亡率(61.73±2.76)%,e组细胞凋亡率(61.03±3.62)%,f组细胞凋亡率(60.50±1.95)%,各组细胞凋亡率存在显著差异(F=95.55,P=0.000),见图3。c组与a组细胞凋亡率无显著差异,b组细胞凋亡率较a组显著升高(P=0.000),d组、e组和f组细胞凋亡率无显著差异,但与a组和c组比较均有明显升高(P=0.000),和b组比较显著降低(P=0.000)。

a.对照组;b.ALD组;c.Bufalin组;d.ALD+Bufalin组;e.ALD+CQ组;f.ALD+Bufalin+CQ组;*P<0.05vsa组;#P<0.05vsb组;P<0.05vsc组。
图3大鼠系膜细胞凋亡的变化
3讨论
所有肾脏疾病均可能进展为慢性肾衰竭,进而严重影响人类生命长度及质量,寻找新的延缓或阻止肾脏损害的治疗方法十分重要。研究显示醛固酮可能作为一个独立的危险因素直接参与肾损害[5]。系膜细胞、内皮细胞、足细胞等肾脏固有细胞的凋亡(又称Ⅰ型程序性细胞死亡)在肾脏损害的发生发展中,扮演着至关重要的角色[6-9],醛固酮被证实可诱导系膜细胞凋亡[1],但其机制目前研究甚少。
自噬又称Ⅱ型程序性细胞死亡,是一个吞噬自身细胞质或细胞器并使其包被进入囊泡,形成自噬体,并与溶酶体融合形成自噬溶酶体,降解其所包裹的内容物的过程,藉此实现细胞本身的代谢需要和某些细胞器的更新。在细胞饥饿、生长因子缺乏时,自噬对维持细胞的存活起到一定的积极作用,而过度的自噬可引起细胞死亡。自噬参与足细胞、肾小球内皮细胞、肾小管上皮细胞等肾脏固有细胞的损伤[9],在急性和慢性肾脏疾病实验模型中,自噬已被证明对肾细胞的存活至关重要[11],干预肾疾病细胞的自噬,可能会阻止肾脏疾病的进展。自噬与凋亡间关系密切而复杂,某些情况下,细胞因应激诱导自噬增强,从而避免了凋亡;而另一些情况下,过度的自噬又可导致细胞的凋亡[12-13],二者的动态平衡或失衡决定了细胞的生存或死亡,因此阐明自噬和凋亡的动态关系对细胞命运转归的作用机制具有重要意义,也有助于寻找治疗肾脏疾病的有效措施。
在细胞自噬的研究中,用透射电子显微镜观察细胞质中双层膜包裹所形成的自噬体是评估自噬的形态学金标准。LC3是一种自噬相关蛋白,在自噬体形成中起核心作用,胞浆可溶性LC3.Ⅰ在泛素样修饰后,与自噬膜表面的磷脂酰乙醇胺结合,形成LC3.Ⅱ被募集到自噬前体上,形成自噬体,LC3.Ⅱ含量与自噬泡数量成正比,可用于判断自噬水平的高低[14]。P62蛋白,通过LRS结构域与LC3相互作用而定位在自噬体上,其C末端结构域连接泛素化蛋白,并引导自噬体进行选择性降解,当自噬功能障碍时,聚集在自噬体上的P62不能被降解,导致P62蛋白水平明显增高[15]。因此,P62是评价自噬体降解的指标。本研究结果显示ALD组可见典型自噬体,LC3、P62表达量及细胞凋亡率较对照组明显升高;Bufalin组未见典型自噬体,LC3、P62表达量及细胞凋亡率与对照组基本相同,但较ALD组明显下降;ALD+Bufalin组见自噬体较ALD组减少,LC3、P62表达量及细胞凋亡率较ALD组下降;ALD+CQ组见自噬体明显多于ALD组,LC3、P62表达量较ALD组显著升高,但细胞凋亡率较ALD组下降;ALD+ Bufalin+CQ组见自噬体多于ALD+Bufalin组,但少于ALD+CQ组,LC3、P62表达量及细胞凋亡率与ALD+CQ组基本相同。上述结果提示醛固酮可诱导系膜细胞自噬及凋亡,蟾蜍灵可抑制醛固酮诱导系膜细胞自噬及凋亡,自噬抑制氯喹通过组织自噬体与溶酶体结合抑制自噬,造成自噬体堆积,同时抑制凋亡。
综上提示醛固酮可能诱导系膜细胞自噬促进细胞凋亡,这可能是引起的肾损伤的机制之一。因此本研究结果可以为醛固酮在肾损害的作用机制研究中提供一定的实验基础和理论依据,并为醛固酮抑制剂、蟾蜍灵及氯喹治疗相应肾脏疾病提供依据。
【参考文献】
[1]施会敏,张俊,何燕芳,等.体外醛固酮对大鼠系膜细胞凋亡基因的影响[J].医学研究生学报,2015,28(3):260-263.
[2]DE RECHTER S,DECUYPERE JP,IVANOVA E,etal.Autophagy in renal diseases[J].Pediatric Nephrology,2015,30:1-16.
[3]EISENBERG-LERNER A,Bialik S,Simon HU,etal.Life anddeath partners:apoptosis,autophagy and the cross-talk between them[J].Cell Death Differ,2009,16(7):966-975.
[4]郭玉兰,李善文,张爱青,等.蟾蜍灵对体外培养的大鼠肾小球系膜细胞分泌肿瘤坏死因子-α与白细胞介素-1β的影响[J].医药导报,2009,28(9):1114-1116.
[5]NAVANEETHAN S D,NIGWEKAR S U,STRIPPOLI GF. Aldosterone antagonists for preventing the progression of chronic kidney disease[J]. Cochrane Database of Systematic Reviews,2014,4(4):297-301.
[6]QIU W,ZHOU J,ZHU G,etal.Sublytic C5b-9 triggers glomerular mesangial cell apoptosis via XAF1 gene activation mediated by p300-dependent IRF-1 acetylation[J].Cell Death Dis,2014,5 (4):e1176.
[7]CHEN KC,PENG CC,HSIEH CL,etal.Exercise ameliorates renal cell apoptosis in chronic kidney disease by intervening in the in- trinsic and the extrinsic apoptotic pathways in a rat model[J].Evid Based Complement Alternat Med,2013,2013(6):368450.
[8]陈瑜,刘翠萍,茅晓东,等.糖基化终产物诱导小鼠足细胞内质网应激和细胞凋亡的研究[J].医学研究生学报,2013,26(11):1129-1133.
[9]LI X,PABLA N,WEI Q,etal.PKC-delta promotes renal tubular cell apoptosis associated with proteinuria[J].J Am Soc Nephrol,2010,21(7):1115-1124.
[10] 谭进,刘华锋,陈孝文.自噬与肾脏固有细胞损伤[J].中国中西医结合肾病杂志,2012,13(9):836-838.
[11] DONG Z.Introduction:autophagy in kidneys[J].Semin Nephrol,2014,34(1):1.
[12] RUBINSTEIN AD,KIMCHI A.Life in balance-a mechanistic view of the crosstalk between autophagy and apoptosis[J].J Cell Sci,2012,125(11):5259-5268.
[13] DING WX,YIN XM.Mitophagy:mechanisms,pathophysiological roles,and analysis[J].Biol Chem,2012,393(7):547-564.
[14] ROMAO S,MUNZ C.LC3-associated phagocytosis[J].Autophagy,2014,10(3):526-528.
[15] PANKIV S,CLAUSEN TH,LUMARK T,etal.p62/AQATM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy[J].J Biol Chem,2007,282(33):24131-24145.
Effects of autophagy on the apoptosis of mesenteric cells induced by aldosterone in rats with
invitrotechniqueQUGaoting,ZHANGAiqing,SHIHuimin,GANWeihua
Department of Pediatrics,The Second Affiliated Hospital of Nanjing Medical University,Nanjing 210003,China
【Abstract】Objective: To investigate the potential mechanisms of renal injury associated with aldoserone(ALD) through observing the effects of autophagy on the apoptosis of mesenteric cells(MCs) induced by ALD and bufalin in rats with in vitro techniques.Methods:The mesenteric cells in rats were cultured with in vitro technique and induced with ALD,bufalin and chloroquine phosphate(CQ),respectively.Then rats were divided into groups of controls(group A),treated with ALD(10-6mol/L,group B),bufalin(10-8mol/L,group C),ALD+Bufalin(10-6mol/L +10-8mol/L,group D),ALD+CQ(10-6mol/L+10 μmol /L,group E) and ALD+Bufalin+CQ(10-6mol/L+10-8mol/L+10 μmol /L,group F).Transmission electron microscope was used to observe the autophagy in the mesenteric cells in each group of rats,and Western Blot was performed to determine the protein changes of P62 and LC3 suggestive of autophagy.Cellular apoptosis was detected with An-nexin V /PI flow cytometry.Results:Transmission electron microscopy showed no typical autophagosome in group A and C,yet prominent autophagosome in group B.Lower autophagosomes were seen in group D,whereas higher autophagosomes were found in group E and F.Western blotting revealed similar level of P62 and LC3 protein in group C to group A,yet higher level in group A than that of group A and C(P<0.05),lower level in group D than that of group B(P<0.05).Level of P62 and LC3 protein in group E was comparable to that of group F,yet the level was higher than remaining groups(P<0.05).Flow cytometry indicated the rate of cellular apoptosis of (46.23 ±5.83)% in group C and (46.10±3.49)% in group A.Although group B had higher incidence of apoptosis (76.47 ±3.03)% than group group A(P<0.05),yet it remained similar among groups of group D(61.73 ±2.76)%,group E(61.03 ±3.62)% and group F(60.50±1.95)%.However,apoptosis was higher in group A and C,and lower in group B (P<0.05).Conclusion:Aldosterone may induce autophagy as well as apoptosis of the mesenteric cells,and bufalin and chloroquine can inhibit the autophagy and apoptosis induced by ALD.
【Key words】aldosterone;bufalin;mesenteric cells;apoptosis;autophagy
【文献标识码】【中图号】R 692A
作者简介:曲高婷(1990-),女,2014级硕士研究生,(电话)17701596663,(电子信箱)302936422@qq.com;
收稿日期:2015-08-22
文章编号:1002-0217(2016)02-0113-05
甘卫华,女,主任医师,(电子信箱)weihuagan@njmu.edu.cn,通讯作者.
