北京永定河沿河沙地杨树人工林光能利用效率
2016-05-23仇宽彪张志强康满春查同刚蔡永茂赵广亮
仇宽彪, 张志强,*, 康满春, 查同刚, 牛 勇, 蔡永茂, 赵广亮
1 北京林业大学水土保持与荒漠化防治教育部重点实验室,北京 100083 2 北京市八达岭林场,北京 102102
北京永定河沿河沙地杨树人工林光能利用效率
仇宽彪1, 张志强1,*, 康满春1, 查同刚1, 牛勇1, 蔡永茂2, 赵广亮2
1 北京林业大学水土保持与荒漠化防治教育部重点实验室,北京1000832 北京市八达岭林场,北京102102
摘要:光能利用效率(LUE)是影响生态系统生产力大小和质量的主要因素。以位于北京市大兴区永定河沿河沙地的杨树(欧美107/108, Populus euramericana cv.)人工林生态系统作为研究对象,依托涡度相关观测系统,对该生态系统的LUE进行研究,从而确定LUE在不同时间尺度上的影响因子,并确定最大光能利用利用效率(LUEmax)。结果表明:LUE存在明显的季节变化趋势,4月份生长季开始后LUE迅速升高,到7—8月达到最大,而后逐渐降低;在生长季不同阶段, LUE日动态的影响因子不同:4月份气温(Ta)、蒸散比(EF)和饱和水汽压差(VPD)是影响LUE日动态的主要因子,7、8月份光合有效辐射(PAR)和冠层导度(gc)是主要影响因子,5—6月与9—10月LUE日动态则与土壤水分(VWC)有较大关系;而LUE月动态则与月蒸散比(EFm)和月平均土壤温度(Tsm)有关。由于该人工林各月光能利用最适宜环境条件不同,各月LUEmax也各有差异,该生态系统年LUEmax为0.44 gC/MJ PAR,7、8月LUEmax最大,分别为0.66和0.69 gC/MJ PAR。研究结果表明,在利用光能利用模型进行区域乃至全球初级生产力估算时需要根据研究的不同时间尺度确定LUEmax。
关键词:光能利用效率;生态系统总初级生产力;涡度相关;杨树人工林
光能利用效率(LUE)是植被吸收单位光合有效辐射所生产的干物质[1- 2],是影响生态系统生产力大小与质量的主要因素[3- 4]。LUE受到温度、水分、光照辐射以及养分等因素的影响[5- 7]。基于LUE,目前已有多个光能利用效率模型被被广泛应用于不同尺度陆地生态系统总初级生产力(GPP)或净初级生产力(NPP)的估算研究[8- 10]。光能利用效率模型具有如下形式:
GPP=APAR×LUEmax×f(T,W,…)
(1)
式中,APAR为生态系统所吸收的光合有效辐射,LUEmax为生态系统最大光能利用效率,f(T,W,…)表示影响生态系统LUE的环境因子,包括温度、水分等。
早期的估算研究[11- 12]对LUEmax的取值大多基于进化论的观点而对不同陆地生态系统取某一近似值[13- 14]。但近来的研究表明LUEmax也受到诸如植物种类[15- 17]等因素的影响,而单一的LUEmax会增加模型估算结果的不确定性[18]。此外,各光能利用效率模型实际估算中所使用的最大光能利用效率也不尽相同,比如:Veroustraete利用C-fix模型对欧洲森林的碳固定模拟中使用1.1 gC MJ-1[19]; Landsberg在使用3-PG模型估算澳大利亚、新西兰森林生态系统生产力时使用的1.8 gC/MJ[20];在EC-LUE模型中应用2.14 gC MJ-1[21];Li在估算中国生态系统GPP时采用2.25 gC/MJ[22]。伍卫星等选择不同生长季半小时白天的净生态系统交换(NEE)和光量子通量密度(PPFD)数据,采用表观量子效率作为LUEmax,其值在0.054—0.0248 μmol CO2μmol/photon[23]。对于落叶阔叶林,Xiao采用0.528 gC/mol作为LUEmax[8, 24]。LUEmax的不同导致模型估算结果具有较大差异[25]。
不同时间分辨率下,生态系统LUE的变化特征不同。短期内LUE波动较大,而长期LUE波动则较小[26- 28]。在不同的时间尺度内,LUE的影响因子也不同。LUE的逐日变化与PAR及散射辐射比例有关[28- 32],LUE的逐月变化则与温度、水分有关[5, 7, 33]。因此,在应用光能利用效率模型时还需考虑不同时间分辨率下对LUE有显著影响的环境因子。
目前,LUEmax的确定仍依赖于模型的分析。有的研究采用光合响应曲线,通过表观量子效率作为LUEmax的度量。但这种方法无法界定除光照外其他环境气象条件的作用。有的研究采用一定的光能利用效率模型进行LUEmax的估算Potter[12]和Zhu[34- 36],但该方法受到模型结构的影响,不同的光能利用效率模型中,影响LUE的因子也不尽相同。Potter和Zhu均采用CASA模型分别在全球和全国尺度进行NPP的估算,因此采用CASA模型中有关温度和水分的标量估算LUEmax是合理的。
杨树(PopulusL.)是我国主要的造林树种,主要用于用材林、防护林等林种。目前,全国杨树人工林面积达700余万hm2,分布广泛,具有较大的林分生产力和经济效益[37]。目前,对杨树的光能利用效率研究大多集中在不同品种杨树苗的光能利用效率方面[38- 39],对杨树人工林生态系统光能利用效率不同时间尺度上动态变化研究还较少[28],而且,对杨树人工林的LUEmax研究还很缺乏。由于针对不同树种进行GPP或NPP估算可提高区域生态系统生产力的估算精度[6],本文利用位于北京市大兴区永定河沿河沙地杨树人工林生态系统多年连续涡度相关观测数据,研究该人工林光能利用效率不同时间尺度的变化,确定了该人工林不同时间尺度下光能利用效率的影响因素,比较不同时间尺度该人工林最大光能利用效率,以期为区域杨树人工林生产力估算提供依据。
1研究区域
本研究地点位于北京市大兴区榆垡镇大兴林场,东经116 °15 ′07 ″,北纬39 °31 ′50 ″,西距永定河0.8 km,林场东西、南北长均为44 km,总面积1030.6 km2。该地区属于暖温带半湿润气候区,年均气温11.5 ℃,年均无霜期209d,年均日照总时数2772h,多年平均降水568.9 mm,降水年际波动较大,最少降水量261.8 mm,最大降水量1058 mm,降水年内分配不均,全年降水总量的60%—70%集中在7、8、9三个月份。研究地点内土壤为冲积性沙壤土,土质疏松,通气透水性较好,但土壤养分含量低。研究地点植被主要为1998—2003年间营造的杨树人工林,株行距2 m×4 m,2006—2009年胸径分别为10.8 cm、12.2 cm、13.8 cm和14.5 cm,树高分别为11.5、13、14.8 m和16.2 m。林下植被以草本为主,主要包括尖头叶藜(ChenopodiumacuminatumWilld)和菊科的黄花蒿(ArtemisiaannuaL.)以及紫苜蓿(MedicagosativaLinn)等。
2数据来源与研究方法
2.1通量与微气象观测
大兴林场地形平坦,符合涡度相关观测要求。涡度相关观测系统以高度为22.5 m的观测塔为载体,主要观测仪器包括测定辐射的光量子探头(Li190SB-L, Li-Cor, NE)、净辐射仪(Q7.1, REBS),测定水和气体含量的红外气体分析仪(Li-7500, Li-Cor, NE),测定风速的三维超声风速仪(CSAT3, CS, USA),气压计(CS105, CS, USA),自记雨量计(TE525-L, CS, USA),测定林内5、10、15 m和20 m温湿度的温湿度传感器(HMP45AC prob, CS, USA),以及安装在地下5、10和20 cm处的土壤温度传感器(TCAV107, CS, USA)、土壤热通量板(HFT3, Seattle, USA),以及安装在地下5、20 cm处的水分观测仪TDR(CS616, CS, USA)。2006—2009年,由于林分生长,辐射观测仪器、红外气体分析仪及三维风速计安装高度分别为16、18、18和20 m。
2.2通量数据处理与计算
涡度相关观测系统所获得数据需要经过质量控制。涡度相关数据处理流程包括稳态测试、平面坐标拟合、WPL校正以及大气稳定度分析、临界风速μ*确定、异常点剔除、数据插补以及质量控制与分析等步骤。2006—2009年μ*分别为0.1266、0.1174、0.1139和0.1312。采用能量闭合作为质量控制的衡量指标,2006—2009年间在0.7—0.9之间,观测数据可靠[40]。缺失数据按照时长进行数据插补:对小于2h的数据缺口采用线性内插法,对2h到7d的数据缺口采用邻近7d相同时段的观测平均值,对大于7d的数据缺口,通过区分NEE和Re,采用Michaelis-Menten(公式(2))和Lloyd-Taylor方程(公式(3))进行插补[41- 42],GPP可按公式(4)计算得到:
(2)
(3)
GPP=-NEE+Re
(4)
式中,α为表观量子效率(μmol CO2/μmol PAR),Qp为光合有效辐射(μmol m-2s-1),最大光合速率(μmol CO2m-2s-1),Rd表示暗呼吸速率(μmol CO2m-2s-1),R10为某一参考温度下的呼吸值(Tref=283.15,K=10℃),Ea为活化能(kJ mol-1K-1),R为气体常数(8.3134 mol-1K-1),Ta为气温(K)。
逐日LUE与逐月LUE分别采用公式(5)和(6)进行计算:
(5)
(6)
式中,i为每月内天数,j表示月份,GPP为每天总初级生产力(gC m-2d-1),PAR为光合有效辐射(MJ m-2d-1)。
2.3数据分析与统计
为研究生长季内不同时期LUE的主要影响因子,本文采用多重逐步回归方法,对各月内逐日LUE与PAR、气温(Ta)、土温(Ts)、蒸散比(EF)、冠层导度(gc)、饱和水汽压差(VPD)以及土壤含水量(VWC)之间的关系进行研究。其中,VPD、VWC以及EF作为可反映生态系统水分状况的指标。较高的VPD将引起气孔关闭,空气阻力增加,叶片光合速率降低[5]。当出现水分胁迫时,叶片气孔会关闭,导致植物叶片温度升高,表现为显热通量的增加。因此采用EF可表示生态系统水分条件[43-44],并在EC-LUE模型中有所应用[21]。gc及EF的计算方法如下[45]:
(7)
(8)
(9)
式中,Δ为饱和水汽压与温度曲线的斜率 (kPa/K),γ为干湿表常数 (kPa/K),Rn为净辐射(MJ m-2d-1),G为土壤热通量(MJ m-2d-1),LE为潜热通量(MJ m-2d-1),ρa为空气密度(kg/m3),cp为空气定压比热(J kg-1K-1), VPD为饱和水汽压差(kPa),γa为空气动力学阻力 (s/m),zm和zh分别为风速和水分测量高度 (m),d为零平面位移 (m),zom和zoh分别为热量和水汽粗糙度长度 (m),k为冯卡曼常数(0.41),u为平均水平风速。Hs为显热通量(MJ m-2d-1)。
为研究生长季LUE的主要影响因子,本文采用偏相关分析与多重逐步回归方法,研究逐月LUE与月光合有效辐射(PARm)、月平均气温(Tam)、月平均土壤温度(Tsm)、月蒸散比(EFm)、月均土壤水分(VWCm)和月均饱和水气压差(VPDm)和月降水量(Pm)之间的关系。
根据对逐日LUE和逐月LUE有显著影响的环境因子,采用决策树算法确定该人工林LUE达到最大的环境条件组合,然后对GPP和PAR进行标准线性回归,其斜率即为LUEmax。决策树各分支通过复杂度损失修剪的进行剪枝,以确定最优组合。以上数据分析均在R3.0.2中进行[46]。
3结果与分析3.1杨树LUE日变化及其影响因素
图1为研究区2006—2009年环境因子日变化特征图。从图中可见,2006—2009年,日平均气温变化范围为-14—31 ℃,年均气温为12.67 ℃。4a平均降雨量为560 mm,光合有效辐射( PAR) 从冬季到夏季逐渐增加,随后逐渐降低。大气饱和水汽压差( VPD) 在温度较低的冬季达到极小值,在高温干旱条件下达到极大值。土壤水分( VWC) 与降雨量变化较为一致,生长季保持在2%—17%之间。2006—2009年LUE分别为(0.33±0.16)gC/MJ、(0.35±0.23) gC/MJ、(0.39±0.16) gC/MJ和(0.32±0.19) gC/MJ。

图1 2006—2009年环境气象因子以及LUE日动态变化图Fig.1 The daily dynamics of environmental factors and LUE from 2006 to 2009
不同月份LUE日动态的影响因子不同(表1)。逐步回归结果显示,4月份LUE日动态主要受Ta、EF和VPD影响,这3个变量可解释该月LUE变化的67%; 5月份LUE日动态的主要受PAR、VWC、EF和gc的影响,它们可解释LUE的79%;6月份LUE日动态的主要影响因子为PAR、VWC、gc和VPD,此4个变量可解释LUE变化的79%;影响7、8月份LUE日动态的主要因子为PAR和gc,此两个变量可解释7、8月近80%的LUE变化。PAR、Ts、VWC和EF是影响9月份LUE日动态的主要因子,其可解释LUE变化的52%。PAR、VWC、EF、gc和VPD是影响10月份LUE日动态的主要因子,其可解释LUE变化的92%。此外,逐月的LUE日动态回归模型离差(3.27)小于生长季LUE回归模型离差(5.38),生长季LUE回归估算在生长季前期和后期具有较大的误差(图2)。因此在估算日LUEmax时需要按每月分别进行。

表1 2006—2009年LUE日动态与环境气象因子逐步回归分析表
Ta: 日气温(℃),Ts: 日土温(℃),EF: 日蒸散比,PAR: 日光合有效辐射(MJ m-2d-1),VWC: 0—20cm深度土壤平均含水量(%),gc: 冠层导度(mm/s),VPD: 饱和水气压差(kPa)
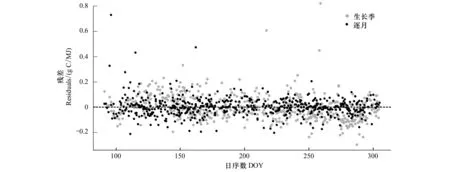
图2 生长季与逐月LUE回归估算残差图Fig.2 The comparison between the residuals of LUE estimates for the growing season and each month虚线表示残差为0
3.2杨树LUE月变化及其影响因素
图3为研究区2006—2009年环境因子月变化特征图。从图中可见,研究区各环境因子均具有明显的季节变化特征。2006—2009年,月平均气温变化范围为-3—26 ℃,4a中降雨主要集中在7、8月,该两月的平均降水量达759 mm和436 mm,光合有效辐射( PAR) 从冬季到夏季逐渐增加,随后逐渐降低。月均VPD在1月达到极小值,在6月达到极大值。土壤水分( VWC) 在1月达最小值,在7月达最大值,生长季保持在7%—9%之间。
3.3LUEmax估算
各月LUEmax估算结果见表4。从标准回归系数来看,LUEmax也与LUE有相似的月动态变化趋势,4—5月份LUEmax迅速升高,7—8月份达到最大值,随后逐渐降低。4月份时,EF≥0.42和EF<0.42时逐日LUE差别最大,4月份LUEmax为0.22 gC/MJ;5月份17≤PAR<27且EF≥0.77时逐日LUE最高,月LUEmax为0.39 gC/MJ;6月份VPD<1.2 kPa且PAR≥21 MJ时逐日LUE高于其他条件,月LUEmax为0.38 gC/MJ;7月以后PAR成为逐日LUE差异的分支点,但不同月份PAR的临界点不同,自7月份到10月份,这一临界值逐渐降低。7—9月份3个月LUEmax没有显著差异(P>0.05)。
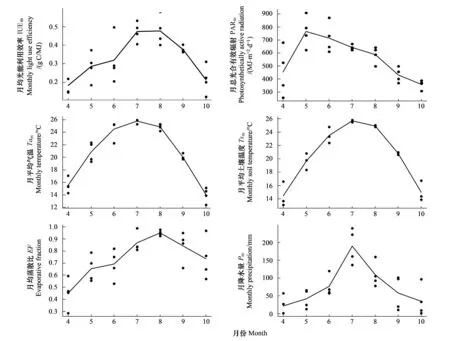
图3 2006—2009年环境气象因子及LUE月动态变化图Fig.3 The monthly dynamics of environmental factors and LUE from 2006 to 2009
LUE月动态与PARm相关系数仅为0.14,没有显著的相关关系(P=0.48),Tam、EFm、Tsm以及Pm与LUE月动态相关程度较高,其相关系数分别为0.63、0.79、0.71和0.69(P<0.01,表2)。多重逐步回归结果显示(表3),Tsm和EFm对LUE月动态有显著影响,两者可解释LUE月动态的71%,两者回归系数分别为0.01和0.39(T=3.19、4.61,P<0.01)。而且两者回归系数均大于0,表明随着Tsm和EFm的升高,月LUE逐渐升高。以上分析表明,月平均土壤温度和蒸散比是影响LUE月动态的主要因素。

表2 2006—2009年LUE月动态与环境气象因子相关分析表
gcm: 0.41(0.03), vpdm:-0.35(0.07)
生长季内逐月LUE主要与EFm有关。EFm=0.77可作为LUE月动态显著差异的分支点。EFm≥0.77时,生长季内LUEmax为0.44 gC/MJ。

表3 2006—2009年LUE月动态与环境气象因子逐步回归系数

表4 每月LUEmax估算表
4讨论
从叶片光合速率的变化来看,在叶片伸展完毕到叶片面积达到最大这段时间内,其光合速率增加较迅速,随着叶面积停止生长,叶龄继续增长时,净光合速率则有所下降[28]。由于春季降水较少,叶面积迅速增大的杨树林对水分需求量增加,水分成为制约杨树林LUE的主导因子,这种状况一直持续到雨季来临。8月后,LUE则持续下降,这与降水减少导致的VWC减少以及温度降低有关[47],还有的研究也指出这也与叶片养分含量下降、叶片年龄等因素相关[31]。2006和2009年5月份LUE较4月份的增幅大于2007和2008年,在2009年5月份LUE甚至高于6月份LUE。这是因为在2006年和2009年5月份实验地对林地进行了灌溉。水分条件的改善提高了生态系统的LUE。
尽管在生长季前期和后期,水分因子对LUE的影响较显著(表1),但在决策树分析中水分不再是影响后期月LUE的主要节点(表4),这表明生长季前期降水对杨树人工林生态系统GPP的影响更为显著。
相比于土壤温度,气温对LUE的影响更为显著。在4月份,气温与LUE呈正相关,表明气温对生长季杨树人工林生长季开始的重要意义,这与Chen的研究类似[48]。但在北方针叶林、落叶阔叶林生态系统,土壤温度却是生态系统生长期开始的主要影响因子[49- 50]。
光能利用效率模型中使用的LUEmax多根据植物所吸收的PAR(PAR×fPAR,fPAR为植物吸收的光合有效辐射比例,fraction of absorbed PAR)进行计算。而本文之所以直接使用PAR是因为本文的研究目的在于估算站点尺度的LUEmax,而关于fPAR的遥感数据产品空间分辨率较大,易引起混合像元问题。但为了与已有植被类型的LUEmax进行比较,本文根据MOD15A2数据产品,选取通量塔所在像元,提取fPAR并粗略计算LUEmax,结果见表5。由于fPAR与叶面积有关,因此呈现明显的季节变化趋势。4—5月间fPAR迅速升高,5—6月间fPAR变化不大,6—8月间fPAR增加并达到生长季内最大值,9月后逐渐降低。为便于比较各月LUEmax,通过fPAR进行修正。推算出年LUEmax在1.09 gC/MJ。根据研究,不同生态系统具有不同的光能利用效率均值和范围[31]。作物的LUE集中在1.1—1.4 gC/MJ,但对于自然生态生态系统,LUE的变化范围更大[51]。在哈佛森林,GPP的光能利用效率大约在1.1 gC/MJ[52]。我国落叶阔叶林NPP合成LUEmax为0.692 gC/MJ[34],根据GPP-LUE与NPP-LUE大体为2:1推算,我国落叶阔叶林LUEmax当在1.38 gC/MJ。但各月的LUEmax有所差异,在对生长季内月度GPP进行估算时需要考虑各月LUEmax。

表5 站点像元每月fPAR平均值
表中fPAR和LUEmax分别表示植物吸收的光合有效辐射比例(fraction of absorbed PAR)和最大光能利用效率(Light use efficiency)
5结论
通过研究沙地杨树人工林不同时段LUE的变化,确定不同时间分辨率下LUE的主导影响因子,并估算沙地杨树人工林的LUEmax。研究发现:(1)生长季开始后LUE迅速升高,到7月中旬时达到最大值,随后GPP逐渐降低,11月开始进入休眠期。生长季早期的灌溉可提高杨树人工林逐月LUE;在生长季不同阶段,逐日LUE的影响因子不同,4—5月与9—10月逐日LUE则与土壤水分(VWC)有较大关系;而逐月LUE则与月蒸散比(EFm)和月平均土壤温度(Tsm)有关;沙地杨树人工林年LUEmax为0.44 gC/MJ PAR,由于各月对沙地杨树人工林LUE最适宜的环境条件不同,据此估算的LUEmax也各异,各月中以7、8月LUEmax最高,分别为0.66和0.69 gC/MJ PAR。研究结果表明,在利用光能利用模型进行区域乃至全球初级生产力估算时需要根据研究的不同时间尺度确定LUEmax。
参考文献(References):
[1]Prince S D. A model of regional primary production for use with coarse resolution satellite data. International Journal of Remote Sensing, 1991, 12(6): 1313- 1330.
[2]Running S W, Nemani R R, Heinsch F A, Zhao M, Reeves M, Hashimoto H. A continuous satellite-derived measure of global terrestrial primary production. BioScience, 2004, 54(6): 547- 560.
[3]吴俊铭. 贵州光能资源的基本特征、时空分布规律及其合理利用. 贵州气象, 2001, 25(4): 13- 18.
[4]同小娟, 李俊, 王玲. 农田光能利用效率研究进展. 生态学杂志, 2008, 27(6): 1021- 1028.
[5]同小娟, 李俊, 于强. 农田生态系统光能利用效率及其影响因子分析. 自然资源学报, 2009, 24(8): 1393- 1401.
[6]张秀娟, 韩海荣, 王军邦. 亚热带人工针叶林光能利用效率的差异. 东北林业大学学报, 2010, 38(12): 12- 14.
[7]隋雪梅, 辛晓平, 张宏斌, 海全胜, 包刚, 闫瑞瑞, 包玉海. 呼伦贝尔贝加尔针茅草甸草原光能利用率变化规律分析. 中国农业资源与区划, 2013, 34(5): 27- 35.
[8]Xiao X M, Zhang Q Y, Braswell B, Urbanski S, Boles S, Wofsy S, Moore B III, Ojima D. Modeling gross primary production of temperate deciduous broadleaf forest using satellite images and climate data. Remote Sensing of Environment, 2004, 91(2): 256- 270.
[9]Potter C S, Davidson E A, Klooster S A, Nepstad D C, De Negreiros G H, Brooks V. Regional application of an ecosystem production model for studies of biogeochemistry in Brazilian Amazonia. Global Change Biology, 1998, 4(3): 315- 333.
[10]Goetz S J, Prince S D, Goward S N, Thawley M M, Small J, Johnston A. Mapping net primary production and related biophysical variables with remote sensing: Application to the BOREAS region. Journal of Geophysical Research: Atmospheres, 1999, 104(D22): 27719- 27734.
[11]Monteith J L. Solar radiation and productivity in tropical ecosystems. Journal of Applied Ecology, 1972, 9(3): 747- 766.
[12]Potter C S, Randerson J T, Field C B, Matson P A, Vitousek P M, Mooney H A, Klooster S A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Global Biogeochemical Cycles, 1993, 7(4): 811- 841.
[13]Field C B. Ecological scaling of carbon gain to stress and resource availability //Winner W E, Pell E J, Roy J eds. Response of Plants to Multiple Stresses. San Diego: Academic Press, 1991: 35- 65.
[14]Goetz S J, Prince S D. Modelling terrestrial carbon exchange and storage: evidence and implications of functional convergence in light-use efficiency. Advances in Ecological Research, 1999, 28: 57- 92.
[15]Xiao X M, Hollinger D, Aber J, Goltz M, Davidson E A, Zhang Q Y, Moore B III. Satellite-based modeling of gross primary production in an evergreen needleleaf forest. Remote Sensing of Environment, 2004, 89(4): 519- 534.
[16]Li A N, Bian J H, Lei G B, Huang C Q. Estimating the maximal light use efficiency for different vegetation through the CASA model combined with time- series remote sensing data and ground measurements. Remote Sensing, 2012, 4(12): 3857- 3876.
[17]Prince S D, Goward S N. Global primary production: a remote sensing approach. Journal of Biogeography, 1995, 22(4/5): 815- 835.
[18]Goerner A, Reichstein M, Tomelleri E, Hanan N, Rambal S, Papale D, Dragoni D, Schmullius C. Remote sensing of ecosystem light use efficiency with MODIS- based PRI. Biogeosciences, 2011, 8(1): 189- 202.
[19]Veroustraete F, Sabbe H, Eerens H. Estimation of carbon mass fluxes over Europe using the C-Fix model and Euroflux data. Remote Sensing of Environment, 2002, 83(3): 376- 399.
[20]Landsberg J J, Waring R H. A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. Forest Ecology and Management, 1997, 95(3): 209- 228.
[21]Yuan W P, Liu S G, Zhou G S, Zhou G Y, Tieszen L L, Baldocchi D, Bernhofer C, Gholz H, Goldstein A H, Goulden M L, Hollinger D Y, Hu Y M, Law B E, Stoy P C, Vesala T, Wofsy S C. Deriving a light use efficiency model from eddy covariance flux data for predicting daily gross primary production across biomes. Agricultural and Forest Meteorology, 2007, 143(3/4): 189- 207.
[22]Li X L, Liang S L, Yu G R, Yuan W P, Cheng X, Xia J Z, Zhao T B, Feng J M, Ma Z G, Ma M G, Liu S M, Chen J Q, Shao C L, Li S G, Zhang X D, Zhang Z Q, Chen S P, Ohta T, Varlagin A, Miyata A, Takagi K, Saiqusa N, Kato T. Estimation of gross primary production over the terrestrial ecosystems in China. Ecological Modelling, 2013, 261- 262: 80- 92.
[23]伍卫星, 王绍强, 肖向明, 于贵瑞, 伏玉玲, 郝彦宾. 利用MODIS影像和气候数据模拟中国内蒙古温带草原生态系统总初级生产力. 中国科学D辑: 地球科学, 2008, 38(8): 993- 1004.
[24]张丽景, 葛宏立. 利用MODIS数据估测毛竹林总初级生产力. 浙江农林大学学报, 2014, 31(2): 178- 184.
[25]Ruimy A, Kergoat L, Bondeau A, Intercomparison T P O F T P N M. Comparing global models of terrestrial net primary productivity (NPP): analysis of differences in light absorption and light-use efficiency. Global Change Biology, 1999, 5(S1): 56- 64.
[26]Norman J M, Arkebauer T J. Predicting canopy light-use efficiency from leaf characteristics//Handks J, Ritchie J T eds. Modeling Plant and Soil Systems. Madison: American Society of Agronomy, Inc., 1991: 125- 143.
[27]牛铮, 王长耀. 碳循环遥感基础与应用. 北京: 科学出版社, 2008: 206- 210.
[28]李泽晖, 王云龙, 魏远, 马悦, 陈润芝. 湖南岳阳杨树人工林光能利用率动态特征分析. 资源科学, 2012, 34(10): 1832- 1838.
[29]Sims D A, Rahman A F, Cordova V D, Baldocchi D D, Flanagan L B, Goldstein A H, Hollinger D Y, Misson L, Monson R K, Schmid H P, Wofsy S C, Xu L K. Midday values of gross CO2flux and light use efficiency during satellite overpasses can be used to directly estimate eight-day mean flux. Agricultural and Forest Meteorology, 2005, 131(1/2): 1- 12.
[30]魏远, 张旭东, 江泽平, 周金星, 汤玉喜, 吴立勋, 黄玲玲, 高升华. 湖南岳阳地区杨树人工林生态系统净碳交换季节动态研究. 林业科学研究, 2010, 23(5): 656- 665.
[31]Turner D P, Urbanski S, Bremer D, Wofsy S C, Meyers T, Gower S T, Gregory M. A cross-biome comparison of daily light use efficiency for gross primary production. Global Change Biology, 2003, 9(3): 383- 395.
[32]Jenkins J P, Richardson A D, Braswell B H, Ollinger S V, Hollinger D Y, Smith M L. Refining light-use efficiency calculations for a deciduous forest canopy using simultaneous tower-based carbon flux and radiometric measurements. Agricultural and Forest Meteorology, 2007, 143(1/2): 64- 79.
[33]Runyon J, Waring R H, Goward S N, Welles J M. Environmental limits on net primary production and light-use efficiency across the Oregon transect. Ecological Applications, 1994, 4(2): 226- 237.
[34]Zhu W Q, Pan Y Z, He H, Yu D Y, Hu H B. Simulation of maximum light use efficiency for some typical vegetation types in China. Chinese Science Bulletin, 2006, 51(4): 457- 463.
[35]康婷婷, 高苹, 居为民, 黄金龙. 江苏省农作物最大光能利用率时空特征及影响因子. 生态学报, 2014, 34(2): 410- 420.
[36]陈静清, 闫慧敏, 王绍强, 高艳妮, 黄玫, 王军邦, 肖向明. 中国陆地生态系统总初级生产力VPM遥感模型估算. 第四纪研究, 2014, 34(4): 732- 742.
[37]刘文国, 张旭东, 黄玲玲, 刘玲, 张鹏. 我国杨树生理生态研究进展. 世界林业研究, 2010, 23(1): 50- 55
[38]刘磊, 庞洪影, 杨峰, 黄凤兰, 孟凡娟. 3种引种杨树和3种乡土杨树苗叶的光合特性比较. 西部林业科学, 2010, 39(3): 14- 19.
[39]周永斌, 马学文, 姚鹏, 崔建国. 不同生长速度杨树品种的光合生理特性研究. 沈阳农业大学学报, 2007, 38(3): 336- 339.
[40]方显瑞. 杨树人工林生态系统碳交换及其环境响应. 北京: 北京林业大学, 2011.
[41]Lloyd J, Taylor J A. On the temperature dependence of soil respiration. Functional Ecology, 1994, 8(3): 315- 323.
[42]Noormets A, Chen J Q, Crow T R. Age-dependent changes in ecosystem carbon fluxes in managed forests in northern wisconsin, USA. Ecosystems, 2007, 10(2): 187- 203.
[43]Suleiman A, Crago R. Hourly and daytime evapotranspiration from grassland using radiometric surface temperatures. Agronomy Journal, 2004, 96(2): 384- 390.
[44]付刚, 沈振西, 张宪洲, 石培礼, 何永涛, 孙维, 武建双, 周宇庭. 利用蒸散比和气温模拟藏北高寒草甸的光能利用效率. 自然资源学报, 2012, 27(3): 450- 459.
[45]Monteith J L. Evaporation and environment. Proceedings of the 19th Symposia of the Society for Experimental Biology. Cambridge: University Press, 1965, 205- 234.
[46]R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: R Foundation for Statistical Computing, 2013.
[47]陈效逑, 周萌, 郑婷, 张荣菊. 呼伦贝尔草原羊草(Leymuschinensis)光合速率的季节变化——以鄂温克旗牧业气象试验站为例. 生态学报, 2008, 28(5): 2003- 2012.
[48]Chen X Q, Hu B, Yu R. Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Global Change Biology, 2005, 11(7): 1118- 1130.
[49]Jarvis P, Linder S. Constraints to growth of boreal forests. Nature, 2000, 405(6789): 904- 905.
[50]Baldocchi D D, Xu L K, Kiang N. How plant functional-type, weather, seasonal drought, and soil physical properties alter water and energy fluxes of an oak-grass savanna and an annual grassland. Agricultural and Forest Meteorology, 2004, 123(1/2): 13- 39.
[51]Russell G P, Jarvis P G, Monteith J L. Absorption of radiation by canopies and stand growth// Russell G, Marshall B, Jarvis P G eds. Pland Canopies: Their Growth, Form and Function. Cambridge: Cambridge University Press, 1989, doi: 10.1017/CBO9780511752308.003.
[52]Wofsy S C, Goulden M L, Munger J W, Fan S M, Bakwin P S, Daube B C, Bassow S L, Bazzaz F A. Net exchange of CO2in a mid-latitude forest. Science, 2007, 260(5112): 1314- 1317.
The dynamics of light use efficiency at a poplar plantation in Beijing
QIU Kuanbiao1, ZHANG Zhiqiang1,*, KANG Manchun1, ZHA Tonggang1, NIU Yong1, CAI Yongmao2, ZHAO Guangliang2
1KeyLaboratoryofSoilandWaterConservationandDesertificationCombating,MinistryofEducation,CollegeofSoilandWaterConservation,BeijingForestryUniversity,Beijing100083,China2BadalingForestFarm,Beijing102102,China
Abstract:Light use efficiency (LUE) is a major limiting factor of gross ecosystem productivity (GPP). Various LUE models have been established to evaluate regional GPP.The various maximum light use efficiency (LUEmax) values used in these models are critical variables that influence model uncertainty. Since the dynamics and influential factors affecting LUE at different temporal resolutions vary, it is unclear whether the values of LUEmaxat these temporal resolutions differ. Therefore, we examined the dynamics of LUE and LUEmaxusing data from a poplar plantation (Populus euramericana cv.) in the Daxing district, Beijing. Eddy covariance measurements were taken at this study site. A multiple stepwise regression procedure and recursive partitioning methods were applied at both monthly and annual scales. The results indicate that the averagely daily LUE values from 2006 to 2009 were (0.33 ± 0.16) gC/MJ, (0.35 ± 0.23) gC/MJ, (0.39 ± 0.16) gC/MJ,and (0.32 ± 0.19) gC/MJ, respectively. The daily LUE varied seasonally, with a rapid increase occurring in April and May, a peak from Jun to Aug, and a gradual decrease after September. The factors influencing daily LUE were different during different parts of the growing season. Air temperature (Ta), evaporative fraction (EF), and vapor pressure deficit (VPD) were the main factors in affecting LUE in April. In May, photosynthetically active radiation (PAR), volumetric water content (VWC), EF, and canopy conductance (gc) were the factors with the greatest influence. PAR, VWC, gc, and VPD had large impacts on LUE in June. In July and August, LUE was controlled by PAR and gc. In September, PAR, soil temperature (Ts), VWC, and EF were the main influencing factors, while PAR, VWC, EF, gc, and VPD influenced LUE in October. PAR was the most important factor regularizing LUE in the middle of the growing season, while moisture conditions were the main influencing factors early and late in the growing season. However, monthly PAR (PARm) was not a main factor affecting monthly LUE (LUEm). In contrast, 71% of LUEm variations were explained by the monthly evaporative fraction (EFm) and monthly soil temperature (Tsm). Because of various influential factors, LUEmaxwere not identical among temporal resolutions. Recursive partitioning analysis showed that EF = 0.42 was the node for LUE in April. Correspondingly, LUEmaxin Apr was 0.22 gC/MJ, when EF ≥ 0.42. PAR and EF were the nodes for LUE in May, LUEmaxin May was 0.39 gC/MJ,when 17 ≤ PAR < 27 MJ and EF ≥ 0.77. In June, LUEmaxwas 0.38 gC/MJ when VPD < 1.2 kPa and PAR ≥ 21 MJ. From July to October, PAR was the main node for LUE, when LUEmaxwas 0.66 gC/MJ, 0.69 gC/MJ, 0.61 gC/MJ, and 0.44 gC/MJ, respectively. LUEmaxin July, August, and September was slightly larger than that in other months. The average annual LUEmaxwas approximately 0.44. We concluded that iLUE models should incorporate different LUEmaxat different temporal scales to better model GPP.
Key Words:light use efficiency; ecosystem gross primary productivity; eddy covariance; poplar plantation
DOI:10.5846/stxb201408241671
*通讯作者
Corresponding author.E-mail: zhqzhang@bjfu.edu.cn
收稿日期:2014- 08- 24; 网络出版日期:2015- 07- 29
基金项目:林业公益性行业科研专项“森林经营对生态系统碳水耦合变化的影响机理研究”(201204102); 北京市教育委员会科学研究与研究生培养共建项目
仇宽彪, 张志强, 康满春, 查同刚, 牛勇, 蔡永茂, 赵广亮.北京永定河沿河沙地杨树人工林光能利用效率.生态学报,2016,36(6):1571- 1581.
Qiu K B, Zhang Z Q, Kang M C, Zha T G, Niu Y, Cai Y M, Zhao G L.The dynamics of light use efficiency at a poplar plantation in Beijing.Acta Ecologica Sinica,2016,36(6):1571- 1581.
