陕北水蚀风蚀交错带柠条适应不同生境的形态和生理可塑性
2016-05-06陈丽茹陈伟月李秧秧陈佳村
陈丽茹,陈伟月,李秧秧,陈佳村
(1 西北农林科技大学 林学院,陕西杨陵 712100;2 西北农林科技大学 水土保持研究所,陕西杨陵 712100;3 中国科学院水利部水土保持研究所,陕西杨陵 712100)
陕北水蚀风蚀交错带柠条适应不同生境的形态和生理可塑性
陈丽茹1,陈伟月1,李秧秧2,3*,陈佳村3
(1 西北农林科技大学 林学院,陕西杨陵 712100;2 西北农林科技大学 水土保持研究所,陕西杨陵 712100;3 中国科学院水利部水土保持研究所,陕西杨陵 712100)
摘要:以陕北水蚀风蚀交错带4个不同坡位与土壤质地生境[沟底+坝淤绵沙土(A)、梁坡+红黄土(B)、坡顶+绵沙土(C)、坡顶+风沙土盖绵沙土(D)]中生长的柠条为研究对象,研究了其生长、光合和水力性状的可塑性变化,以揭示柠条对不同生境的适应机制。结果表明:(1)生境D和生境A样地1~3 m和3 m土层的平均含水量明显高于生境B和C;(2)与生境B和C相比,生境A和D中生长的柠条冠幅、株高和新枝长显著增加,叶厚度减小,正午叶水势、净光合速率、气孔导度和蒸腾速率亦显著增加,但不同生境中柠条枝比导水率、Huber值和栓塞程度无显著差异;(3)柠条各测定指标中,新枝长、净光合速率、气孔导度和蒸腾速率表现出较大的可塑性。研究认为,柠条可能主要通过新枝长或光合生理特征的改变来适应不同的水分生境。
关键词:柠条;生境;光合特征;水力性状;可塑性
表型可塑性是植物器官在复杂的环境中产生一系列不同的相对适合的表现型的潜能[1-2],是个体适应环境的方式。可塑性越大,在短时段内可促进植物的快速定居,长时段内则通过基因的改变促使植物适应新的环境条件,可塑性增加了植物在高度变化环境中的适应性[3]。
在黄土高原水蚀风蚀交错带,地面梁峁起伏,切割破碎,沟壑纵横,土壤类型、质地和地形等下垫面条件变化很大,因而造成植物生长的生境明显不同。柠条是豆科锦鸡儿属的一种旱生落叶灌木,具有很强的抗旱、抗寒、抗风沙等特性,是干旱半干旱地区防风固沙和保持水土的主要树种,其在黄土高原水蚀风蚀交错带的沟底、梁峁坡和坡顶都有分布。由于水蚀或风沙沉积厚度不同,不同坡位土体的空间构型亦不同,从而导致此地区柠条生长的生境差别很大。揭示柠条对这种不同生境的适应性机制对其合理栽植和经营管理具有重要的理论指导价值。
水分是影响黄土高原植物生长的主要限制因子,不同生境下水分的有效性直接决定着植物的适应性。植物生长依赖于光合过程同化的碳,而光合过程又受植物水分传输能力的直接影响,研究表明:气孔导度、光合速率与土壤-植物-大气连续体(SPAC)水力导度之间经常有很显著的线性相关关系[4-5],因而气体交换和水力性状是植物重要的水分生理特征,已被广泛用来阐释大尺度上植物的地理分布[6-8],是否亦可解释同一地理位置不同生境下植物的适应性?为此,本试验进行了黄土高原水蚀风蚀交错带不同生境下柠条生长、光合和水力性状的可塑性研究,试图揭示柠条对不同生境的适应机制。
1材料和方法
1.1试验区概况
研究区位于陕西省神木县六道沟流域( 38°46′~38°51′N,110°21′~110°23′E,海拔1 094~1 274 m),属于典型的水蚀风蚀交错带,流域面积约为6.89 km2。该地区属中温带半干旱气候,冬春季干旱少雨,夏秋季多雨,多年平均降水量为437 mm,70%以上的降水集中在7~9月份,年均气温8.4 ℃,全年无霜期153 d,年日照时数2 836 h,年总太阳辐射5 922 MJ·m-2。主要土壤类型有绵沙土、新黄土、红土及在沙地发育起来的风沙土和沙地淤土。植被为沙生植物和黄土区植物交错出现,属于森林草原带向干草原过渡地带[9]。流域内除现存少量天然灌木如山榆(Ulmusmacrocarpa)、黄刺玫(Rosexanthina)、文冠果(Xanthocerassorbifolia) 和绣线菊(Spiraeasalicifolia)等之外,主要以人工林为主,包括柠条(Caraganakorshinskii)、沙柳(Salixpsammophila)、紫穗槐(Amorphafruticosa) 等灌木林,及以旱柳(Salixmatsudana)和小叶杨(Populussimonii)为主的乔木林。
1.2试验设计
试验在六道沟流域的一个支沟进行,坡向为东北向,从沟底沿坡向布设4个样地,样地A位于沟底,土质为坝於绵沙土;样地B位于梁坡,土质为红黄土;样地C位于坡顶,土质为绵沙土;样地D位于坡顶,土质为风沙土盖绵沙土。其中,绵沙土属黄绵土的一个亚类,土壤颗粒组成相对均一,细砂粒在42.4%~57.9%,粗粉砂为27.9%~41.1%,物理性粘粒为14.1%~17.6%,粘粒为11.1%~14.1%,土壤容重为1.35~1.48 g·cm-3,总孔隙度为45%~50%,毛管孔隙度在32%~38%之间;红黄土是指新黄土被剥蚀后,老黄土出露地表,其颗粒组成极不均一,细砂粒含量为10.0%~45.9%,粗粉砂为22.9%~48.4%,物理性粘粒为12.7%~67.0%,粘粒为9.0%~22.3%,土壤容重为1.31~1.74 g·cm-3,总孔隙度为35.6%~51.2%,非毛管孔隙度在9.5%~14.8%之间;风沙土颗粒组成相对均一,以细砂为主,细砂粒在59.0%~71.1%之间[10]。4个样地柠条栽植年限均为30年左右,每样地分别打3钻,深度为3 m,每20 cm间隔取土,烘干法测定土壤含水量,同时选取长势良好的柠条6丛,用于植物形态指标(植株生长状况和叶性状)和生理指标(叶水势、氮含量、光合、导水率、茎栓塞程度等)的测量。测定在2014年7~8月份进行。
1.3测定项目及方法
1.3.1植株生长状况包括冠幅、株高、分枝数和当年生新枝长,均用常规方法测量。
1.3.2叶性状和氮含量主要包括叶厚度、叶密度、比叶重。每丛剪取3枝当年生枝条,除新叶外的所有叶组成混合样用于叶性状研究。样品先称鲜重,再用扫描仪(台湾明基5520型扫描仪)扫描,后用Image-J软件分析计算叶面积(LA),最后将样品置于75 ℃烘箱中烘干48 h,称其干重。烘干质量与叶面积之比即为比叶重(LMA),鲜重与叶面积之比用来衡量叶厚度大小(LT,g·cm-2),烘干质量与鲜重之比用来衡量叶密度(LD,%)大小。烘干样品粉碎过筛后,用硫酸-双氧水消煮,采用全自动凯氏定氮仪测定叶氮含量。
1.3.3叶水势叶水势用压力室( 美国PMS仪器公司,PMS 1000型)测定,黎明前叶水势测定时间为4:30~5:30,正午叶水势为晴天12:00~14:00。每丛上测定2~3片叶。
1.3.4光合气体交换参数叶片光合气体交换参数用CI-340光合仪(美国CID公司)来测定,测定时间为9:30~11:30之间,测定时的光强、温湿度和CO2浓度与外界大气相同,测定分两个连续晴天进行,每丛上测定2片叶。
1.3.5茎导水率及正午原位栓塞程度黎明前采集柠条2~3年生枝条,在水下从基部剪取,然后基部放入水中带回到实验室。在实验室中,水下剪取用作测定导水率的茎段,茎段长度必须超过木质部最大导管长度,同时水下去除叶,然后茎段与木质部栓塞测定仪 (XYL’EM,法国Bronkhorst公司)相连,在大约3 kPa压力下用过0.22 μm滤膜的20 mmol/L KCl+1 mmol/L CaCl2的溶液测定茎导水率(Kh),茎导水率与木质部横截面积之比即为比导水率(Ks),与叶面积之比为叶比导水率(Kl),茎木质部横截面积与其上承载的所有叶面积之比即为Huber值。
正午原位栓塞程度采样时间为12:00~14:00,采样方法与导水率的基本相同。枝条带回到实验室后,在距基端约120%木质部最大导管长度处水下取5~6个约3 cm长的茎段,与XYL’EM仪器相连,测定初始导水率(Ki),然后在0.15 MPa压力下冲刷3 min,测定冲刷后的最大导水率(Kmax),测定压力和所用溶液与导水率测定相同。栓塞程度(PLC)用导水率损失的百分数表示,PLC=(1-Ki/Kmax)×100%。
1.4数据统计分析
不同生境中柠条形态与生理功能之间的差异显著性用单因素方差分析进行,当方差分析达到显著后,用LSD法进行多重比较。
2结果与分析
2.1不同生境土壤含水量比较
4个生境样地的土壤含水量在50 cm土层内以生境D最低,而50~100 cm土层之间又以生境A样地较低(图1);在1 m土层内,各样地平均土壤含水量差异不大,生境A、B、C和D样地分别为4.75%、6.04%、5.49%和5.66%;而在1~3 m土层内,生境A~D 4个样地平均土壤含水量分别为9.15%、5.34%、6.85%和13.56%,生境D和生境A样地土壤含水量明显高于其它2个样地。3 m以内土层的平均含水量以生境D最高(9.61%),生境A次之(6.95%),而生境C和B较低(6.17%和5.69%)。从整个土壤剖面看,生境C样地土壤含水量随土壤深度变化不大,生境B样地呈现“上湿下干”的趋势,而生境A和D表现出明显的“上干下湿”现象。
2.2不同生境柠条植株形态指标比较
2.2.1生长状况4个生境中柠条植株的冠幅、株高、分枝数和新枝长显著不同(表1)。其中,柠条植株冠幅和株高在生境A和生境D更大;其分枝数在生境B和生境D最多,而在生境A中最少;柠条新枝长在生境A、C和D间无显著差异,而在生境B下最小。从生长情况看,生境A和D生长的柠条冠幅、株高和新枝长相对高于其它2个生境,这与其较高的立地土壤含水量有关,而分枝数与立地土壤含水量的关系不很紧密。
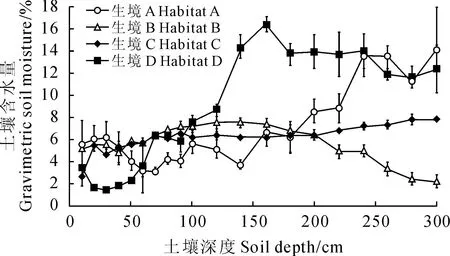
A.沟底+坝淤绵沙土;B.梁坡+红黄土;C.坡顶+绵沙土;
2.2.2叶性状不同生境柠条的叶性状参数见表2。不同生境中柠条叶片比叶重和叶密度无显著差异,但叶厚度有显著差异,并以生境C最高,生境D最小,且两者间有显著差异,其它2种生境介于中间;枝叶比以生境B最高,其它3个生境差异不显著,而生境C显著低于生境B;叶氮含量则以生境A最高,而其它生境之间差异不显著,生境C和D的叶氮含量显著低于生境A。以上结果说明生境B和C趋向于通过增加叶厚度及/或枝叶比来抵御其不良的水分状况。
2.3不同生境柠条生理指标比较
2.3.1水力性质4个生境中柠条黎明前叶水势差异不大,平均为-0.42 MPa,而正午叶水势有显著差异,生境D的叶水势最高并与其他生境间差异显著,其次为生境A和生境B,而生境C的最低;4个生境的柠条茎比导水率、比叶导水率、胡伯尔值和正午时茎原位栓塞程度均差异不显著(表3)。表明不同生境茎水分传输能力并无不同,造成不同生境正午叶水势差异的原因可能主要在于根系吸水和蒸腾能力的差异。
2.3.2光合作用参数由表4可知,柠条叶片的净光合速率、气孔导度和蒸腾速率在4个生境中均存在显著差异,并以生境A最高,生境D、B次之,而生境C则最低,生境C的净光合速率、气孔导度和蒸腾速率分别低于生境A 48.6%、47.9%和56.2%,且这种差异达到显著水平;4个生境的叶片瞬时WUE无显著差异(表4)。以上结果说明生境A和D具有高的光合和蒸腾能力。
2.4不同生境柠条形态与生理特征的可塑性
利用Valladares等提出的可塑性指数(PI)来评价某个形态或生理特征的表型变异[11]:PI=(Xmax-Xmin)/Xmax,其中Xmax和Xmin分别为不同生境中某个特征的最大值和最小值。PI值的变化范围为0~1,该值越大,表明某个特征的可塑性越大。
在测定的8个柠条形态指标中,分枝数和新枝长的可塑性指数较大,分别为0.469和0.427,而其它指标的可塑性指数都相对较小;在11个生理指标中,叶净光合速率、气孔导度、蒸腾速率和比导水率的可塑性指数较高,均大于0.42(表5)。但通过前面的分析可知,柠条分枝数与立地土壤水分并不密切相关(表1),而不同生境的比叶导水率也并不存在显著差异(表3),表明柠条主要依靠新枝长和光合生理参数(净光合速率、气孔导度和蒸腾速率)的变化来适应不同的水分生境。

表1 不同生境下柠条植株生长状况
注:同列不同小写字母表示生境间差异达到显著(P<0.05);下同。
Note:Different normal letters indicate significance between habitats at 0.05 level;The same as below.
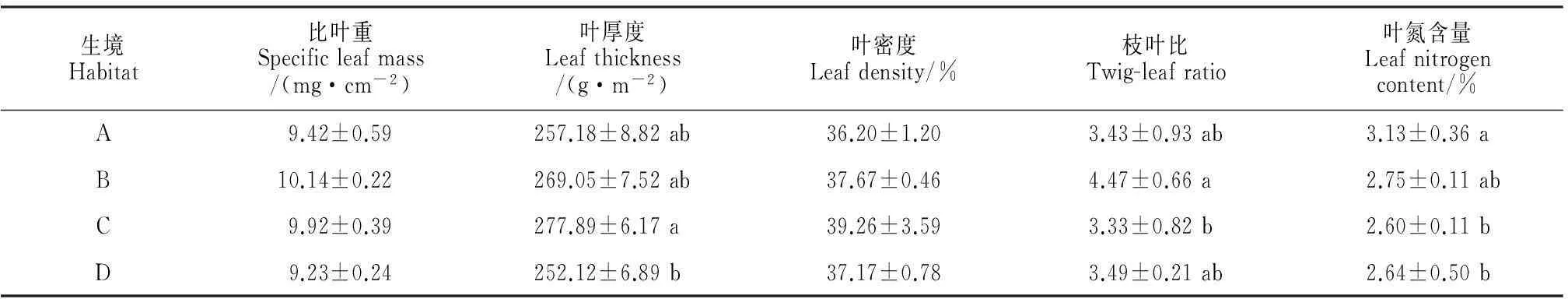
表2 不同生境下柠条叶性状参数

表3 不同生境下柠条叶水势和导水率

表4 不同生境柠条叶光合参数
3讨论
本试验中不同生境的土壤含水量垂直分布明显不同,生境B、生境C样地整个剖面的平均土壤含水量明显低于生境A和生境D。生境B中红黄土是指新黄土被剥蚀后,老黄土出露地表,土壤中有料礓石,土壤保水能力差;绵沙土属于黄绵土的一个亚类,位于坡顶的绵沙土(生境C)由于风和光照的作用,蒸发强烈,因而土壤含水量也较低,而沟底的绵沙土(生境A)由于风小,光照时间短,因而土壤蒸发量小,土壤能维持相对高的含水量。位于坡顶的风沙土盖绵沙土(生境D),尽管上层风沙土的含水量很低,但其孔隙度高,有很高的渗透性,而下层绵沙土的保水能力强,上粗下细的这种土壤质地可降低土壤蒸发[12],因而导致其下层含水量显著高于其他处理。对层状土壤水分的模拟结果表明上部细砂能增加下层粗质地土壤的有效水含量,导致松树和杨树叶面积指数的增加和净初级生产力的提高[13]。
土壤质地对植物水分状况有很大的影响。生长在沙土中的植物比细质地土壤中的植物具有高的叶水势,尽管沙土中根的水力导度小于重质地土壤,但其依靠更多的吸收根增加根系吸水,两种质地叶比水力导度并无显著差异[14]。杨树和云杉在轻质地土壤上根密度要高于重质地土壤[15]。砂质壤土上刺槐蒸腾速率对气象条件变化的敏感度要高于壤质粘土,蒸腾速率下降的临界含水量在砂质壤土上也要高于壤质粘土[16],表明土壤质地影响根系吸水和蒸腾失水过程。本研究中,梁坡+红黄土(生境B)和坡顶+绵沙土(生境C)土壤的质地相对较粗,土壤含水量低,导致柠条在形态和生理结构上发生明显改变。在形态上,柠条灌丛的冠幅、株高和新枝长显著降低,叶厚度增加;在生理上,柠条正午叶水势显著降低,同时其净光合速率、气孔导度和蒸腾速率亦显著降低。其中,柠条形态特征新枝长和光合生理参数(净光合速率、气孔导度和蒸腾速率)表现出较大的可塑性,表明柠条主要通过这些形态或生理结构的改变来适应低水分生境的粗质地土壤。
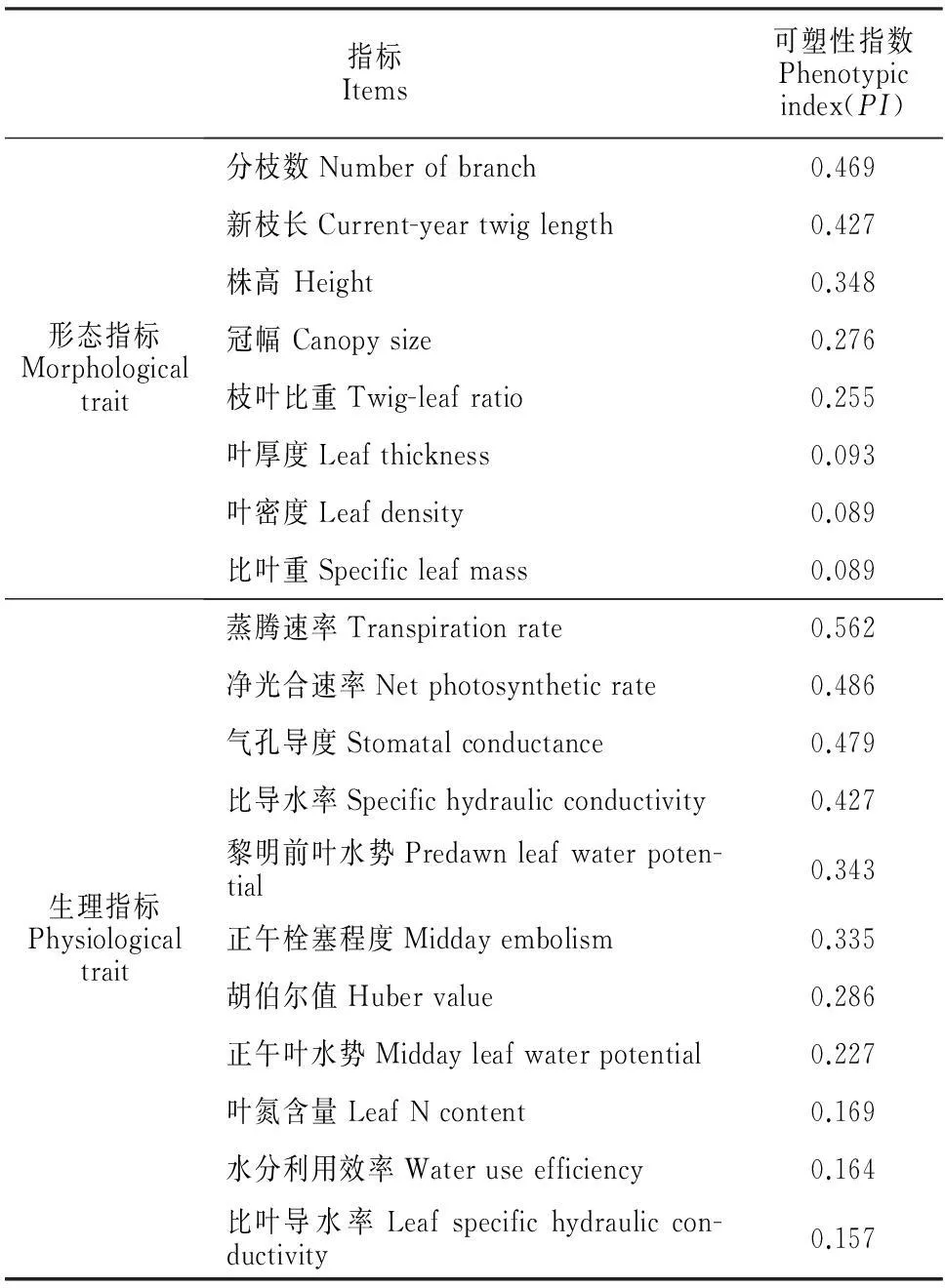
表5 柠条形态和生理指标的可塑性指数
土壤质地改变也可能影响植物的水分传输效率和其安全性。Holste等[17]的研究发现,增加土壤颗粒直径,植物的茎导水率、单位叶面积导水率显著降低;粗质地土壤中生长的火炬松抵抗空穴化能力明显降低[18]。但本研究并未发现粗质地土壤上柠条茎导水率和水分传输安全性有显著差异,一些植物依靠抵抗栓塞能力的增加来适应干旱生境,而另一些植物则不然,柠条可能依靠强的木质部抵抗栓塞的形成,因而其茎水分传输效率和抵抗栓塞能力在不同生境并未表现出明显不同。
除水分因素外,养分也可能是造成不同生境柠条生长差别的原因。尽管生境D样地3 m土层土壤含水量高于生境A,正午叶水势也表现出同样的趋势,但生境A和D的柠条灌丛冠幅、株高和当年生新枝长并无显著差异,且生境A的净光合速率、气孔导度和蒸腾速率更高,原因可能与生境A中柠条具有高的叶N含量有关。坝於绵沙土由于来自于侵蚀的表层土壤,因而养分含量较高,从而导致柠条叶氮含量高,从另一方面证明其它3个生境柠条生长可能受到养分的限制。因而养分限制也可能影响柠条形态和生理特性的可塑性,对此值得进一步研究。
参考文献:
[1]DEWITT T J,ANDREW S,DAVID S W.Costs and limits of phenotypic plasticity[J].TreePhysiology,1998,13(2):77-81.
[2]关保华,葛滢,樊梅英,等.华荠苧响应不同土壤水分的表型可塑性[J].生态学报,2003,23(2):259-263.
GUAN B H,GE Y,etal.Phenotypic plasticity of growth and morphology inMoslachinensisresponds to diverse relative soil water content[J].ActaEcologiaSinica,2003,23(2):259-263.
[3]NICOTRA A B,DAVIDSON A.Adaptive phenotypic plasticity and plant water use[J].FunctionalPlantBiology,2010,37(2):117-127.
[4]BRODRIBB T J,HOLBROOK N M,etal.Hydraulic and photosynthetic co-ordination in seasonally dry tropical forest trees[J].PlantCellandEnvironment,2002,25:1 435-1 444.
[5]HUBBARD R M,STILLER V,RYAN M G,etal.Stomatal conductance and photosynthesis vary linearly with plant hydraulic conductance in ponderosa pine[J].PlantCellandEnvironment,2001,24:113-121.
[6]蒙乾财,张晓飞,刘晓,等.陕北黄土高原辽东栎林分布区北部边界形成的光合作用机制[J].应用生态学报,2011,6:1 409-1 415.
MENG Q C,ZHANG X F,LIU X,etal.Photosynthetic mechanisms of northern boundary formation ofQuercuswutaishanicaforest in Loess Plateau of Northern Shaanxi[J].ChineseJournalofAppliedEcology,2011,22(6):1 409-1 415.
[7]POCKMAN W T,SPERRY J S.Vulnerability to xylem cavitation and the distribution of Sonoran desert vegetation[J].AmericanJournalofBotany,2000,87:1 287-1 299.
[8]BRODRIBB T,HILL R S.The importance of xylem constraints in the distribution of conifer species[J].NewPhytologist,1999,143:365-372.
[9]王建国,樊军,王全九,等.黄土高原水蚀风蚀交错区植被地上生物量及其影响因素[J].应用生态学报,2011,22(3):556-564.
WANG J G,FAN J,WANG Q J,etal.Vegetation above-ground biomass and its affecting factors in water/wind erosion crisscross region on Loess Plateau[J].ChineseJournalofAppliedEcology,2011,22(3):556-564 .
[10]贾恒义,雍绍萍,王富乾.神木试区的土壤资源[J].中国科学院水利部西北水土保持研究所集刊,1993,18(2):36-46.
JIA H Y,YONG S P,WANG F Q.The soil resource in the Shenmu Experimental Area[J].MemoirNISWC,ChineseAcademyofSciences&MinistryofWaterResources,1993,18(2):36-46.
[11]VALLADARES F,ARRIETA S,ARANDA I,etal.Shade tolerance,photoinhibition sensitivity and phenotypic plasticity ofIlexaquifoliumin continental-Mediterranean sites[J].TreePhysiology,2005,25:1 041-1 052.
[12]任利东,黄明斌.砂性层状土柱蒸发过程实验与数值模拟[J].土壤学报,2014,51(6):1 282-1 289.
REN L D,HUANG M B.Experiment and numerical simulation of soil evaporation from layered sandy soil columns[J].ActaPedologicaSinica,2014,51(6):104-110.
[13]HUANG M,BARBOUR S L,ELSHORBAGY A,etal.Effects of variably layered coarse textured soils on plant available water and forest productivity[J].PeocediaEnvironmentalSciences,2013,19:148-157.
[14]LI Y,XU H,COHEN S.Long-term hydraulic acclimation to soil texture and radiation load in cotton[J].PlantCellandEnvironment,2005,28(4):492-499.
[15]PLANTE P M,RIVEST D,VÉZINA A,etal.Root distribution of different mature tree species growing on contrasting textured soils in temperate windbreaks[J].PlantandSoil,2014,380(1-2):429-439.
[16]WU Y Z,HUANG M B,WARRINGTON D N.Black Locust transpiration responses to soil water availability as affected by meteorological factors and soil texture[J].Pedosphere,2015,25(1):57-71.
[17]HOLSTE E K,JERKE M J,MATZNER S L.Long-term acclimatization of hydraulic properties,xylem conduit size,wall strength and cavitation resistance inPhaseolusvulgarisin response to different environmental effects[J].PlantCellandEnvironment,2006,29(5):836-843.
[18]HACKE U G,SPERRY J S,EWERS B E,etal.Influence of soil porosity on water use inPinustaeda[J].Oecologia,2000,124(4):495-505.
(编辑:裴阿卫)
Plasticity of Morphological and Physiological Traits in >Caraganakorshinskiiunder Different Habitats of Water-Wind Erosion Crisscross Region of the Loess Plateau
CHEN Liru1,CHEN Weiyue1,LI Yangyang2,3*,CHEN Jiacun3
(1 College of Forestry,Northwest A&F University,Yangling,Shaanxi 712100,China;2 Institute of Soil and Water Conservation,Northwest A&F University,Yangling,Shaanxi 712100,China;3 Institute of Soil and Water Conservation,CAS & MWR,Yangling,Shaanxi 712100,China)
Abstract:The plasticity of plant morphological and physiological traits is the foundation to probe species adaptation.Hence,the plasticity of growth,photosynthetic and hydraulic traits of Caragana korshinskii growing under different slope positions with different soil texture,including A:gully channel+loessial soil,B:slope+loessial soil,C:slope top+loessial soil and D:slope top+layered soil,were studied in order to explore the mechanism of the extensive distribution of this species under fragmented landscapes on the Loess Plateau.The results indicated that:(1)the soil moisture within 1-3 m and 3 m layer under habitat D and A were higher than those under habitat B and C.(2)C.korshinskii growing under habitat A and D had relatively larger canopy size,plant height and current-year twig length and thinner leaf thickness than those plants under habitat B and C.The midday leaf water potential,net photosynthetic rate,stomatal conductance and transpirational rate were also increased,but no difference existed for specific hydraulic conductivity,Huber value and midday native embolism.(3)Among those measured plant traits,current-year twig length,net photosynthetic rate,stomatal conductance and transpirational rate showed relative larger plasticity,indicting C.korshinskii mainly depend on variation of current-year twig length and photosynthetic traits to adapt to different water habitats.
Key words:Caragana korshinskii;habitat;photosynthetic traits;hydraulic traits;plasticity
中图分类号:Q945.79
文献标志码:A
作者简介:陈丽茹(1988-),女,硕士,主要从事树木水分生理生态研究。E-mail:493705238@qq.com*通信作者:李秧秧,研究员,主要从事植物生理生态研究。E-mail:yyli@ms.iswc.ac.cn
基金项目:国家自然科学基金(41371507)
收稿日期:2015-11-11;修改稿收到日期:2016-02-14
文章编号:1000-4025(2016)03-0573-06
doi:10.7606/j.issn.1000-4025.2016.03.0573
