长羽毛恐龙及翼龙研究新发现
2016-04-26吕君昌中国地质科学院地质研究所北京100037国土资源部地层与古生物重点实验室北京100037
吕君昌中国地质科学院地质研究所,北京 100037;国土资源部地层与古生物重点实验室,北京 100037
长羽毛恐龙及翼龙研究新发现
吕君昌
中国地质科学院地质研究所,北京 100037;国土资源部地层与古生物重点实验室,北京 100037
摘 要:在江西赣州地区晚白垩世地层中发现的窃蛋龙类化石——赣州华南龙,属于进步窃蛋龙类,其独特的头部特征为研究窃蛋龙类恐龙的颅面演化、古地理分布及古生态环境提供重要信息;在辽西早白垩世义县组地层中发现的大型、前肢短的驰龙类恐龙——孙氏振元龙,为大型、短前肢类型的驰龙类提供羽毛形态学方面的重要信息,为研究驰龙类的多样性、鸟类羽毛以及飞行起源提供了重要依据;在辽西发现的喙嘴龙类翼龙——朝阳东方颌翼龙,是该地区晚侏罗世地层中发现的第一件翼龙标本,不仅填补了时代上的空白,同时对于印证晚侏罗世喙嘴龙类的辐射演化具有重要作用。这些重要发现对于研究窃蛋龙类的演化、驰龙类羽毛演化及鸟类羽毛起源等热点与难点问题提供了重要的参考依据,尤其在发现华南龙的基础上,命名了赣州恐龙动物群。
关键词:华南龙;晚白垩世;江西赣州;振元龙;早白垩世;东方颌翼龙;晚侏罗世;辽西
www.cagsbulletin.com www.地球学报.com
本文由中国国家自然基金委(编号:41272022)、中国地质调查局(编号:12120114026801)和中国地质科学院基本业务费项目(编号:JB1504)联合资助。获中国地质调查局、中国地质科学院2015年度地质科技十大进展第十名。
1 赣州恐龙动物群
近年来,在江西省赣州地区晚白垩世红色砂岩地层中发现大量保存极其精美的恐龙骨骼(兽脚类恐龙及蜥脚类恐龙)、恐龙蛋(窝)、恐龙胚胎蛋及其他爬行动物(蜥蜴类及龟类)化石,尤其是窃蛋龙类骨骼和蛋化石,无论在种类、数量及保存状态上,在世界上都是独一无二的,它们共同构成了一个晚白垩世丰富的恐龙动物群,被命名为“赣州恐龙动物群”(Lü et al.,2015a)。目前在江西省赣州地区已命名的窃蛋龙类化石有:斑脊龙(徐星和韩凤禄,2010),赣州龙(Wang et al.,2013),江西龙(Wei et al.,2013),南康龙(Lü et al.,2013a)及华南龙(Lü et al.,2015a),并且还有一些新的种类正在在研究中。窃蛋龙类属于兽脚类恐龙,主要分布于北美(加拿大、美国)及亚洲的蒙古国及中国(Osmólska et al.,2004;Lü et al.,2004,2005;Zanno and Sampson,2005;Lamanna et al.,2014)。在中国主要分布在以下3个地区:北方地区(辽宁省西部的早白垩世、内蒙古晚白垩世的地层中)(Ji et al.,1998;季强等,2012;Xu et al.,2002a,2007;徐星等,2013;Longrich et al.,2010),中原地区(河南省洛阳地区—汝阳盆地早白垩世晚期及栾川盆地晚白垩世的地层中)(Lü et al.,2009,2013b)以及南方地区(广东省河源和南雄盆地晚白垩世,江西省赣州地区上晚白垩世的红色砂岩沉积中)(吕君昌,2005;Lü,2002;Lü et al.,2013a;Wei et al.,2013;徐星和韩凤禄,2010;吕君昌和张宝堃,2005;Wang et al.,2013;Lü et al.,2015a)。原始的窃蛋龙类比如切齿龙(Xu et al.,2002a)、尾羽龙(Ji et al.,1998)以及宁远龙(季强等,2012)都具有牙齿。尾羽龙还具有鸟类一样的尾羽,因此推断进步的窃蛋龙类也应该具有羽毛,只不过由于沉积环境原因(多数为沉积颗粒粗的红色砂岩,无法保存细微的结构),没有保存下来罢了。原始的窃蛋龙类主要发现在我国辽宁省西部地区的早白垩世义县组地层中,其他地区均为没有牙齿的进步窃蛋龙类。进步的窃蛋龙类具有如下独有特征:头骨高,多孔,吻短,无齿而可能具角质喙,下颌外孔大,下颌前背缘上凸,齿骨后端具两个长的突起,前部尾椎椎体具有椎体侧孔等(Osmólska et al.,2004;Barsbold et al.,2000)。
1923年由美国科学家安德鲁斯领导的美国自然历史博物馆中亚科学考察团,在蒙古国西南戈壁的纳摩盖特盆地的巴音扎克(火焰崖)发现了第一件窃蛋龙标本(Andrews,1923)。由于当时发现时,化石骨架正好趴在一窝恐龙蛋上,而在附近又发现许多原角龙类恐龙骨架,因此标本的研究者奥斯本认为这些蛋是原角龙的,这一恐龙是在偷吃原角龙蛋时遭到灭顶之灾的。因此他把它命名为偏食角龙蛋窃蛋龙(Oviraptor philoceratops)。Oviraptor意思是“抢蛋者、窃蛋者”,philoceratops意味着“溺爱角龙类蛋”(Osborn,1924)。值得提及的是奥斯本曾经意识到“这一动物的属种名…可能会完全误导我们对其食性的理解和掩盖其真正的特点”(Osborn,1924)。1994年,在蒙古国发现一恐龙胚胎化石,其蛋大小、形态、表面刻饰及显微构造均与奥斯本1924年认为的原角龙蛋的长形蛋一样,而对这一胚胎化石研究证明,它却是窃蛋龙的(Norell et al.,1994,1995),因此,以前被认为是原角龙蛋其实是窃蛋龙的蛋。从而说明原来认为是偷吃角龙蛋的恐龙,其实是在孵蛋或者是在保护它们自己的蛋(Norell et al.,1994,2001),印证了奥斯本的预言。但是根据古生物命名法则,名称一旦发表,不能更改,虽然对窃蛋龙的行为进行了平反昭雪,但是名字是不会改的了。在蒙古国(Clark et al.,1999)和中国的内蒙古(Dong and Currie,1996)以及江西省赣州地区(Lü et al.,2013a)发现的数具窃蛋龙骨架趴在蛋窝之上,说明了窃蛋龙类像鸟类一样,具有孵蛋行为。
虽然窃蛋龙类具有许多鸟类的特征,比如头骨高度气腔化,颅顶的顶骨部分增大,方骨的耳突双头,方骨上与方轭骨关节处具有侧关节窝(Maryańska and Osmólska,1997);薄或棒状的轭骨(Elżanowski,1999),进步的类群没有牙齿,一些窃蛋龙尾部具有类似鸟类的尾综骨结构(Barsbold et al.,2000),具有鸟类一样的羽毛,尾椎的数目较少等(Ji et al.,1998),它们曾经被认为是次生的、失去飞行能力的鸟类(Elzanowski,1999;Lü,2000;Lü et al.,2002;Maryańska et al.,2002;Feduccia,2012),也就是说它们是中生代的平胸类。最近,Feduccia 和Czerkas(2015)通过对尾羽龙的研究,进一步说明了尾羽龙为次生、失去飞行能力的鸟类,也就是说窃蛋龙类应为不会飞的鸟类;并提出驰龙类、伤齿龙类以及窃蛋龙类等这些核心手盗龙类是处于滑翔、飞行及失去飞行能力的不同阶段的鸟类(Feduccia and Czerkas,2015)。而其它一些研究显示窃蛋龙类与鸟类的关系较远(Sereno,1999;Holtz,2000;Xu et al.,2002b;Zelenitsky et al.,2012)。Xu等通过对在中国辽西早白垩世义县组发现的窃蛋龙类最原始种类——切齿龙的研究,认为切齿龙不具有鸟类特征,这些鸟类特征在进步窃蛋龙类中出现,是趋同演化的结果(Xu et al.,2002a),Balanoff等(2014)通过对Conchoraptor头骨构造的CT研究,认为Conchoraptor所具有的鸟类特征更像是趋同而不是同源的产物。然而,正如Feduccia和Czerkas(2015)所指出的“窃蛋龙类所具有的大量而高度复杂的鸟类特征,如果是由恐龙衍生而来已经很难解释”。窃蛋龙类除了长有一个长的尾巴外,其它形态特征与平胸类相似,它们本身可能就是中生代次生失去飞行能力的鸟类。不管窃蛋龙类的分类位置如何,它们是一独特类群,对于研究鸟类大脑演化的复杂历史,羽毛起源与演化以及窃蛋龙类的生活习性、古地理环境等都具有重要意义。
2 赣州华南龙
目前,在赣州南雄组发现有5个属的窃蛋龙类。赣州的南康地区在40 km2的地域内就发现有超过200个窃蛋龙类的蛋窝(一些具有胚胎化石)。其中,赣州华南龙就是最近发现的窃蛋龙类(Lü et al.,2015a)。
赣州华南龙属于进步的窃蛋龙类,它发现于赣州火车站扩建工地,是最近江西省赣州地区发现的、保存相对完整的标本之一,除了它的头骨和下颌都保存得很好之外,还有7个连续的前部颈椎椎体保存,其中前4个保存完整,后3个部分或者以印痕形式保存,以及部分前肢及后肢保存(Lü et al.,2015a)(图1)。它具有以下综合特征:下颌的方骨关节髁位于枕髁之后;头骨的颈横向脊显著;隅骨占下颌外孔边缘的大部分;齿骨气腔化;第一掌骨长而细,直径为其长度的20%;齿骨的后腹支扭曲,因而其侧面有些朝向腹面;下颌缝合部大于下颌长度的20%而小于25%;每一前指爪子的前背伸肌边缘突出;圆形的上颞颥窝小于下颞颥窝;前上颌骨的后背突与泪骨接触,其末端的后腹角有一个明显的孔;下颌外孔之上的齿骨背边缘部分强烈凹陷。华南龙在以下几个方面不同于其他多数窃蛋龙类:枕髁和方骨向前背方倾斜(与葬火龙一样)以及外鼻孔的前腹角稍微位于穿过眶前窝后背角的水平线之下(这一特征与葬火龙、可汗龙以及豫龙共享);背视,第四颈椎椎体的两后关节突形成一直边缘(该特征与豫龙共享)。
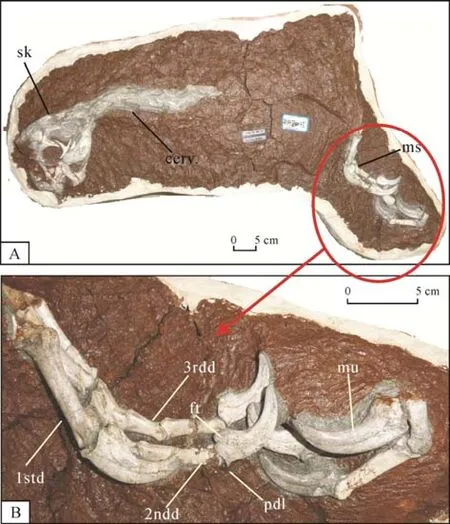
图1 赣州华南龙(HGM41HIII-0443)的正型标本(A)和左右手部的放大(B)(改自Lü et al.,2015a)Fig.1 The holotype of Huanansaurus ganzhouensis(HGM41HIII-0443)(A);a close up of the phalanges of the right and left hand(B)(modified after Lü et al.,2015a)
虽然华南龙与始兴龙(吕君昌和张宝堃,2005)只有胫骨可以比较,但是它们之间区别是很明显的:始兴龙胫骨近端内表面具有一个明显的孔,而华南龙胫骨的该部位没有孔。
华南龙与河源龙区别在于:河源龙的齿骨愈合部分强烈下翻,颈椎椎体的神经弓及颈肋上具有气孔,并且第三掌骨强烈减小(Lü,2002;吕君昌2005),而华南龙的齿骨愈合部分没有下翻,其颈椎椎体的神经弓及颈肋上也不具有气孔,以及第三掌骨大小正常(Lü et al.,2015a)。
从华南龙下颌的构造上看,它区别于赣州龙(Wang et al.,2013)、江西龙(Wei et al.,2013)及南康龙(Lü et al.,2013a)在于它的齿骨前部背边缘的强烈凹陷,而上述后3个窃蛋龙中,该部位几乎平直。
华南龙在以下几个方面区别于发现于内蒙古的乌拉特龙(徐星等,2013):华南龙的前肢爪子前背方具有明显的唇突,而乌拉特龙没有,其外鼻孔的前腹角位于穿过眶前窝后背角的水平线之下。
华南龙区别于斑脊龙在于斑脊龙(徐星和韩凤禄,2010)的外鼻孔相对于眶前孔较大,而在华南龙相对较小;华南龙的下颞颥孔近于梯形,而斑脊龙的近于正方形。斑脊龙的外鼻孔前腹角远远位于穿过眶前窝后背角的水平线之下。
华南龙在以下几个方面区别于蒙古国的葬火龙(Clark et al.,2002):沿头骨中线,其顶骨比额骨短,轭骨的上升突向后背方延伸,下颌外孔之上的齿骨背边缘部分强烈凹陷。葬火龙的下颞颥孔的上边缘和下边缘几乎等长,因而使得下颞颥孔近于不规则四边形,而华南龙的下颞颥孔接近梯形或者近三角形。
华南龙在以下几个方面与纳摩盖特慈母龙(Lü et al.,2004,2005)不同:外鼻孔前腹角远远位于穿过眶前窝后背角的水平线之下,上颞颥孔圆形以及头骨的最高处位于眼眶之上;而纳摩盖特慈母龙的外鼻孔前腹角远远位于穿过眶前窝后背角的水平线之上,上颞颥孔亚圆形,头骨的最高处位于外鼻孔的上方。
华南龙在以下几个方面不同于Rinchenia mongoliensis(Barsbold,1997)(=蒙古窃蛋龙,Osmólska et al.,2004):华南龙没有额骨和顶骨形成的头骨脊,其下颌外孔之上的齿骨背边缘部分强烈凹陷,外鼻孔前腹角稍微位于穿过眶前窝后背角的水平线之下,而Rinchenia mongoliensis的头骨具有明显的由额骨和顶骨形成的脊,其外鼻孔前腹角远远位于穿过眶前窝后背角的水平线之上。
华南龙区别于可汗龙(Balanoff and Norell,2012)在于华南龙的前肢爪子上近端关节处具有明显的唇突,可汗龙的下颞颥孔的腹边缘和后边缘几乎形成一个直角,而华南龙则形成一锐角。

图2 基于42个分类单元和230个特征,由TNT程序分析所获得的60颗最简约树构成的严格合意树,显示了赣州华南龙的系统发育位置(树长为533,一致性指数为0.510,保留指数为0.691)Fig.2 Strict consensus of 60 most parsimonious trees obtained by TNT,based on analysis of 42 taxa and 230 characters,showing the phylogenetic position of Huanansaurus ganzhouensis(tree length = 533,consistency index = 0.510 and retention index = 0.691)
系统发育分析显示华南龙属于窃蛋龙科,与发现于蒙古国的葬火龙关系密切,构成一个分枝。同时显示了发现于南雄组的6个窃蛋龙属嵌入窃蛋龙科的3个分枝中。它们下颌的结构完全不同,南康龙的下颌缝合部的吻端没有下翻,江西龙和华南龙的稍微下翻,而斑脊龙的强烈下翻,这说明了这些窃蛋龙之间具有不同的摄食策略,它们在同一群落生境中很可能占据不同的生态灶。江西省赣州地区发现大量的窃蛋龙类化石表明了这一生物群包含了在白垩纪晚期,亚洲地理上更广泛分布的,具有分类学和形态学意义上的窃蛋龙类的多样性。在中生代末期,这些窃蛋龙类在亚洲陆相生态系统中曾昌盛一时。应该指出的是在中国中原地区的河南省栾川地区晚白垩世晚期发现的窃蛋龙——迷你豫龙(Lü et al.,2013b)与蒙古国发现的Nomingia和江西省赣州地区发现的南康龙均产于马斯赫里特期,且构成一个分枝,而通过研究,南康龙及迷你豫龙是植食性的(Lü et al.,2013a,b),从而推断Nomingia可能也是植食性的。这使我们能够提出这样一个假设,在白垩纪末期,中国可能识别出至少2个独立的窃蛋龙组合。然而,在两个组合之间的物种形成存在一些有趣的细节。北方的12种窃蛋龙发现于从坎潘期到马斯赫里特期的6个组中,而时间和空间受到很好约束的南雄组却发现了6种窃蛋龙。而且,与戈壁地区相比,中国的南方和中原地区仍然有一些相对没有调查研究区域。因此,我们在这里提出的所有关于亚洲窃蛋龙古生物地理学的假设,伴随着新的标本、新的分类单元及新地点的发现,将被进一步检验。
60颗最简约树构成的严格合意树(图2)显示了蒙古国纳摩盖特盆地的窃蛋龙和中国南方南雄组的窃蛋龙存在一定的关系,而北美的新颌龙类形成一单独的分枝,是北美独特的一枝窃蛋龙类。这与白垩纪晚期,亚洲中部和东部的窃蛋龙类持续传播以及后来由于地理隔离造成的物种形成相符合。这好像是窃蛋龙类多个分枝的代表由于适应不同生态灶分区及对各种觅食策略的特化,居住在重叠的地理区域中。因为中国南方南雄组的窃蛋龙和蒙古国纳摩盖特盆地的窃蛋龙在合意树中几乎随意分布,所以这意味着自然扩散的障碍是低的,比如在贯穿整个地理区域具有相似的栖息地。然而,基因流动后来一定受到限制,这是因为祖先亚种群之间的距离最大化最终导致它们应对新的、演化的古环境的隔离和特化。

图3 窃蛋龙类的头骨比较,显示眶前孔的后背角与外鼻孔的前腹角之间的位置关系Fig.3 Skull comparisons of oviraptorosaurs showing relative positions of the posterodorsal corner of the antorbital fenestra and the anteroventral corner of the external narial opening
赣州华南龙发现的意义:赣州华南龙的发现不仅说明了中国南方是窃蛋龙类多样性较高的地区,同时也进一步说明了中国江西省赣州地区是窃蛋龙类化石发现最多的地区之一。它的头骨形态学特征,尤其是外鼻孔与眶前窝的相对位置关系(图3),为了解窃蛋龙类恐龙的颅面演化提供了重要信息。华南龙与其它报道的中国南方发现的窃蛋龙类显著不同,而是与3000 km之外的蒙古国发现的葬火龙十分相似,说明白垩纪末期窃蛋龙在亚洲分布范围更为广泛,并在晚白垩世达到繁盛,这为研究窃蛋龙类的古地理分布及古生态学提供重要证据(中国地质科学院,2016)。
3 孙氏振元龙
辽宁地区以发现长羽毛的恐龙、翼龙类、鸟类及原始哺乳类而著称。尤其是长羽毛恐龙的发现,对研究鸟类起源、小型兽脚类恐龙的演化具有重要的意义。而长羽毛的驰龙类恐龙种类繁多,保存精美,为研究鸟类及羽毛起源提供重要素材。
驰龙类恐龙一般是小到中型长羽毛的兽脚类恐龙,第二脚趾长有大而弯曲的爪子,尾部由于被极度加长的棒状前关节突、后关节突以及脉弧所包裹而显得僵硬,背椎椎体的副突呈凸起的茎状。虽然推断它们起源于晚侏罗世之前,但是化石记录主要发现于白垩纪,它们的身体大小范围从小至知更鸟,大到鸸鹋(Kirkland et al.,1993;Xu et al.,2000;Zheng et al.,2010;Norell and Makovicky,2004;Senter et al.,2012;Turner et al.,2012;Evans et al.,2013)一样。
在过去20年来,中国辽西及其周边地区发现著名的长羽毛恐龙已成为古生物学中最具标准性的化石。这些早白垩世生物群中身披绒毛保存精美的标本完美地说明了由肉食的兽脚类恐龙到鸟类的过渡阶段。最有趣的标本是这些具有恐龙到鸟类演化的基干副鸟类群,其中许多具有前肢形成大的翅膀或者一些情况下后肢的腿上附着的“现代”的大飞羽。目前在辽西地区共发现驰龙类有6个属,它们是长羽盗龙(Han et al.,2014),纤细盗龙(徐星和汪筱林,2004),小盗龙(Xu et al.,2000;Xu et al.,2003),中国鸟龙(Xu et al.,1999;Ji et al.,2001),天宇盗龙(Zheng et al.,2010)和振元龙(Lü and Brusatte,2015);除了天宇盗龙和振元龙外,其他驰龙类的个体均相对较小,体长小于家猫或者中等大小的狗,具有长的前肢和广泛排列的廓羽(Lü and Brusatte,2015)。
振元龙的鉴定特征:振元龙具有以下独特的组合特征,在驰龙类中的自近裔性状以单星号而在辽宁驰龙类中以双星号表示:极度细长的桡骨,其主干比第一手指第一指节细*;第二掌骨变短,比第一掌骨和第一手指第一指节总长还短**;具有6个荐椎椎体**;前肢短,是后肢长度的一半,肱骨和股骨的长度之比小于0.65,尺骨和股骨的长度之比小于0.55;以及耻骨主干侧面中部缺少明显的侧结节(这一特征同样在天宇盗龙和非辽宁驰龙类也缺失,但是出现在长羽盗龙、小盗龙及中国鸟龙中)。振元龙不同于天宇盗龙在于其眶前窝的腹面由明显的边缘所限定(与中国鸟龙的情况一样);肠骨后突末端尖;以及耻骨具有向前凸出的主干(与长羽盗龙、小盗龙及中国鸟龙的情形一样,但是不像天宇盗龙那样直)。振元龙也缺少天宇盗龙和小盗龙所具有的骨化的钩突,但是这不是一个决定性的特征差异,因为在许多辽宁兽脚类恐龙中,一些保存有钩突而另一些没有保存。振元龙和天宇盗龙之间的肢骨比例也稍微不同:前肢稍微短于后肢(振元龙为0.48而天宇盗龙的为0.53)以及手部相对股骨要短(振元龙为0.76而天宇盗龙为0.86)。然而,由于只有两件标本相比较,如果没有种间差异及可能由于埋藏变形引起的不同等更多信息,这些不同鉴定特征的重要性是有问题的。由于在具有许多标本的两个副鸟类—小盗龙(Dececchi and Larsson,2013)和始祖鸟(Mayr et al.,2007;Foth et al.,2014)中,前肢和后肢的比例不同处于种间变化范围之内,因而,这一不同可能不是天宇盗龙和振元龙之间明显的区别特征。
3.1 振元龙标本简要描述
振元龙的详细描述见Lü和Brusatte(2015)。在此只是对其进行简短记述。振元龙是辽宁驰龙类中大的恐龙,缺失部分尾部,保存的体长为126.6 cm(图4),通过与天宇盗龙(Zheng et al.,2010)比较,估计振元龙活着时的体长为165 cm(Lü and Brusatte,2015)。由于兽脚类恐龙股骨的长度与其体重具有很强的对应关系(Christiansen and Fariña,2004),并且辽宁的驰龙类的各个类群至少有一根股骨保存,因此股骨长度是比较辽宁驰龙类的实用指标(Lü and Brusatte,2015)。在目前发现的7个辽宁驰龙类的属中,振元龙的股骨长度仅短于天宇盗龙,而比其他辽宁驰龙的大。振元龙的股骨长度为193.4 mm,天宇盗龙的股骨长度为212.5 mm(Zheng et al.,2010),而其他的辽宁驰龙类的股骨长度均小于160 mm(Lü and Brusatte,2015)。
振元龙除了头骨后部侧颞颥孔及脑腔周围受到强烈挤压外,其他部分保存较好,和中国鸟龙(Xu and Wu,2001)一样,上颌骨的眶前窝的腹面由突出的边缘所限定,而与天宇盗龙和其他的驰龙类不一样。前颌骨孔(Promaxillary fenenstra)大,几乎与颌骨孔一样大,并且背部位于与颌骨孔几乎同一水平线上。大的前颌骨孔同样出现在其他辽宁驰龙类中,比如中国鸟龙和天宇盗龙,但是在疾走龙和查干龙中要小的多。
T形的泪骨侧面上好像具有气腔化凹陷。方轭骨具有突出的后突和细的背支。鼻骨和额骨有明显的侧脊而形成颅顶的侧边缘。额骨伸长,上颞颥孔向前延伸到额骨的背表面上。顶骨破碎,但是保存的部分显示了在其活着的时候,左右顶骨是愈合的。在齿骨中部侧面有一系列大的孔,但是这些孔没有联合成一个明显的沟槽。齿骨、隅骨和上隅骨构成大的下颌外孔的边界。
颈部完整,颈椎椎体的神经弓背视呈X形,小的、薄片状的神经棘位于中部,关节突强烈分开。背椎椎体具有前后加长的四边形神经棘,其末端没有膨胀。荐椎椎体6个,其中4个明显地愈合在一起。尾部由相互交织的骨质棒包裹着,使其更加坚固,这也是驰龙类的特征之一。这些骨棒向前几乎延伸至尾巴的根部。中部尾椎椎体的长度大约是背椎椎体的3倍,与多数的辽宁驰龙类类似,但是要比其它们的成比例加长,尾椎椎体长度大约是背椎的2倍。
振元龙的最重要特性是其前肢相当短,整个前肢(肱骨+尺骨+第二手指)与后肢(股骨+胫骨+第三脚趾)的长度比率为0.48,是辽宁驰龙类中最小的。相比之下,天宇盗龙的这一比率是0.53,在其他辽宁驰龙类标本这一比率大约为0.80(Zheng et al.,2010;Han et al.,2014)。振元龙肱骨的长度相对于股骨的长度来说,不论在辽宁驰龙类还是除了非常基干类群大黑神龙(Mahakala)(Turner et al.,2011)和畸变的半鸟龙亚科的南方盗龙(Novas et al.,2009)之外,是最短的。肱骨与股骨的长度之比为0.63,而天宇盗龙的为0.66,其他的辽宁驰龙这一比率均大于0.75(Han et al.,2014;Brusatte et al.,2013)。
肱骨三角脊短,尺骨强壮弯曲,具有明显的鹰嘴突。振元龙最明显的一个特征是桡骨极度细长,比尺骨细的多,并且其主干中部也比手指第一指节的主干细。振元龙在辽宁驰龙类还有一个独特的特征是第二掌骨比第一掌骨和第一手指的第一指节加起来总长还短。
腰带和后肢基本完整。肠骨前后长,背腹矮,肠骨前突比肠骨后突稍微长。髋臼小,上部由明显的上髋臼脊为边界,后部以肠骨的坐骨柄上大的对转子为边界。耻骨主干向前凸起,类似于长羽盗龙、小盗龙及中国鸟龙中的情形,而不同于天宇盗龙的情形,它的耻骨主干直。耻骨主干前边缘没有明显的侧结节。坐骨比耻骨短的多。股骨比胫跗骨短。胫跗骨与股骨的长度之比约为1.30,除了长羽盗龙(1.10)外,与其它辽宁驰龙类一样。跗蹠骨细长,中间的3个紧密靠在一起。脚趾上的爪子大而弯曲。

图4 大型的,前肢短的辽宁驰龙类孙氏振元龙(JPM-0008)Fig.4 The photograph of the large-bodied,short-armed Liaoning dromaeosaurid Zhenyuanlong suni(JPM-0008)

图5 孙氏振元龙的生活情景图(由赵闯绘制)Fig.5 The living scene of Zhenyuanlong suni(drawn by ZHAO Chuang)
羽毛保存在身体的许多部位,特别是前肢和尾部。前臂上有由大的正羽所形成的、宽的翅膀,这些正羽具有羽轴和钩突。虽然由于埋藏时遭到变形,翅膀的具体形状不清楚,但是很明显翅膀很大,右侧翅膀保存的面积大约800 cm2,而左侧的翅膀的面积为1120 cm2。在右侧的翅膀上可以看到羽毛的详细结构,有覆羽、初级飞羽及次级飞羽。翅膀的大小、形状和羽毛的结构一般与小盗龙(Xu et al.,2003;Li et al.,2012),长羽盗龙(Han et al.,2014),近鸟龙(Hu et al.,2009),始中国羽龙(Godefroit et al.,2013)以及基干鸟类,比如始祖鸟(Wellnhofer,2008;Longrich et al.,2012;Foth et al.,2014)的类似。
振元龙的尾部也有正羽,它们分布在背部。羽轴以一系列细长(约1 mm粗,可达8 mm长)的直线形式保存,它们以与尾部长轴夹角45度的方向,向背后方延伸。由于其余部分保存情况很差,很难确定这些羽毛是对称还是非对称(图5)。
3.2 比较和讨论
天宇盗龙和振元龙体长均大于2 m,并且都具有短的前肢,其长度为后肢的一半。很遗憾,天宇盗龙没有保存羽毛(Zheng et al.,2010),因而这类短前肢的驰龙类是否具有像小型辽宁驰龙类那样的皮肤类型及大的飞羽是个未解的问题。虽然振元龙比天宇盗龙稍微小些,但是同样具有短的前肢和一些特征说明它不同于天宇盗龙。振元龙具有保存完好的身体覆盖物,并且首次提供了短前肢的驰龙类的羽毛形态及其分布。像其他的辽宁驰龙类一样,振元龙具有由多组飞羽构成宽的翅膀和尾部具有大的飞羽,但是不像许多基干副鸟类和鸟类类群那样好像后肢缺少廓羽。振元龙的发现,增加了辽宁驰龙类的多样性,进一步提供在早白垩世长前肢和短前肢的驰龙类共存的证据,同时显示了使得驰龙类系统发育变得复杂的大量的同源性,表明辽宁的驰龙类可能不能构成它们自己独特的分枝(Lü and Brusatte,2015)。
振元龙在以下方面区别于小盗龙(Xu et al.,2000,2003):其第三指的第一指节小于第二指节的2倍,肠骨前突具有尖的末端(与天宇盗龙的一样,而在小盗龙中强烈凸出)。振元龙也缺少小盗龙的2个自近裔性状:远端的前上颌牙齿具有明显的收缩以及非常弯曲且具有突出屈肌突的纤细脚爪。
振元龙区别于中国鸟龙(Xu et al.,1999;Ji et al.,2001)在于拥有比第二和第三相当小的第一前上颌齿,中后部尾椎椎体更加长,是背椎椎体长度的3倍或者更大,肠骨后突尖形(像小盗龙和天宇盗龙一样),而不是成方形。振元龙也缺少中国鸟龙所特有的眶前孔之前侧方的、点状刻蚀密集的眶前窝。
振元龙区别于长羽盗龙(Han et al.,2014)在于其第二前上颌牙齿远远大于第三和第四牙齿(与中国鸟龙一样);尾椎的过渡点在于第7和第10尾椎之间(与小盗龙和天宇盗龙一样,而长羽盗龙位于第7尾椎椎体前部);小的半月形腕骨覆盖第一和第二掌骨的几乎一半(与小盗龙和天宇盗龙一样,不像长羽盗龙那样,半月形腕骨大,且几乎覆盖整个第一和第二掌骨的近端);第三掌骨直(与纤细盗龙和中国鸟龙的一样,而不是像长羽盗龙那样具有弯曲的形态);第四蹠骨与第二和第三蹠骨粗细相当(与小盗龙和中国鸟龙的一样,不同于长羽盗龙,其第四蹠骨要粗得多);第二蹠骨和第四蹠骨远端处于同一水平(与中国鸟龙一样,但是不像小盗龙和长羽盗龙那样第四蹠骨远长于第二和第三蹠骨)。
振元龙区别于纤细盗龙(徐星和汪筱林,2004)在于其缺少所特有的极度加长和纤细的胫跗蹠骨。
振元龙的发现,增强了已经充实的辽宁驰龙类的多样性,振元龙是著名的热河生物群发现的第6件明确的驰龙类。如此高的多样性看起来虽然使人惊奇,但是这些辽宁驰龙类发现于不同的组中(义县组和九佛堂组),也不清楚这些驰龙是不是恰好生活在同一时期,因此可能(确实可能)不是所有的这些物种生活在一起。不管怎样,即使这6个属的驰龙与类似鸟类的类群确实在时间和空间上重叠,如果考虑到支持许多种鸟类的无数现代生态系统背景的情形,这是很正常的。
与其它大多数辽宁驰龙类相比,由于它具有大的体型和相对短的前肢,使其成为异常和稀有动物。它是第二件已发表的体型大而前肢短的驰龙类。
孙氏振元龙发现的意义:孙氏振元龙在前肢和后肢均覆盖有类似现生鸟类羽毛及丝状结构,但与其它驰龙类相比,其体型较大且前肢较短,它的发现为大型、短前肢类型的驰龙类恐龙提供了有关羽毛形态学方面的重要信息。这一新类型,除了前肢覆有羽毛,尾部也覆盖羽毛。孙氏振元龙的发现充分说明,早在白垩世纪早期,短前肢类型的驰龙与长前肢类型同时生存在同一地区。这些生活在一起的物种揭示了驰龙类恐龙系统发育的复杂性,这也进一步说明了,辽宁地区的这些驰龙类可能并没有形成独特的分枝。
4 朝阳东方颌翼龙
翼龙类是已绝灭生物中最迷人和最充满神秘色彩的脊椎动物。众所周知,在地球历史上的中生代时期,由于爬行动物非常繁盛,被称之为爬行动物时代。在这段时期,爬行动物的辐射演化异常剧烈,不同类型的爬行动物占据不同的生存空间,无论在陆地上、水体中(包括河流、湖泊以及海洋),还是空中都有它们活动的痕迹。陆地上主要以形形色色的恐龙类为主,河流湖泊中以龟鳖类、鳄类为主,海洋中主要以鱼龙、蛇颈龙及沧龙为主,而空中则由翼龙类所占据,掌握着当时的制空权。翼龙类是一群飞行爬行动物,它们与同时代的陆地霸王——恐龙类几乎同时产生于晚三叠世末期(约2.3亿年前),且开始适应空中生活,是地球历史上最早获得动力飞行的脊椎动物。它们在地球上生活了将近1.6亿年,最后与其同时代的其他爬行动物如陆地上的非鸟恐龙类、水生爬行动物沧龙类等绝灭于6600万年前的白垩纪末期。翼龙类前肢(翅膀)发达,第5手指退化,而第4手指通常有4个指节,每个指节均极度加长,和前肢其它部分构成司飞行的翼的前缘,翼由翼膜构成。翼龙的后肢退化,有的几乎不能在陆地上行走。翼龙的头骨多孔,骨骼中空,骨壁很薄,所有这些特征均适用于飞行。虽然翼龙类与恐龙类同时代,但是它们不是恐龙,虽然它们会飞且具有翅膀,但是它们也不是鸟类。最近几年来,在我国辽西及其周边地区的中侏罗世到早白垩世的地层中发现了大量种类繁多,且保存精美的翼龙化石,尤其是带胚胎的翼龙蛋化石、过渡类型——达尔文翼龙,以及与其蛋保存在一起的雌性达尔文翼龙化石骨架的发现,使得该地区成为世界上翼龙最丰富的产地之一,可以和德国晚侏罗世的索伦霍芬及巴西的翼龙产地相媲美,甚至超过它们。这些发现为我们进一步了解翼龙的生殖生理学、翼龙的演化、性别的鉴定及对其多样性的探讨提供了重要信息。我国目前发现翼龙化石约有12科,近50个属种,几乎占全世界翼龙属种的一半,并且随着研究的不断深入,还将会发现一些新的属种,使我国成为世界上翼龙化石最丰富的国家(吕君昌等,2013)。
虽然在辽西地区早白垩世的热河生物群发现了大量保存精美的翼龙标本(吕君昌等,2006,2013),但是在更早时期的翼龙主要发现于中侏罗世比如达尔文翼龙,青龙翼龙以及蛙嘴龙科的树翼龙等(Lü et al.,2010,2012;Lü and Hone,2012),而晚侏罗世却很少发现。
朝阳东方颌翼龙是2015年发现的、为目前辽西地区最大的喙嘴龙类翼龙,这也是该地区第一件发现于晚侏罗世的翼龙标本(Lü et al.,2015b)。标本产于辽宁省朝阳市胜利乡东山村。朝阳颌翼龙具有以下鉴定特征:翼掌骨与肱骨的长度之比为0.38(比其它的基干翼龙类要小);尺骨短于第四指每一指节(与基干翼龙双形齿翼龙类相似,与船颌翼龙的相反);尺骨短于胫骨(比率为0.92,而在其它喙嘴翼龙类中,该比率大于1),胫骨与股骨几乎等长以及翼小骨与肱骨的长度比率为0.21。

图6 朝阳东方颌翼龙(41HIII-0418)Fig.6 The holotype of Orientognathus chaoyangensis(41HIII-0418)

图7 朝阳东方颌翼龙生活情景图(由赵闯绘制)Fig.7 The living scene of Orientognathus chaoyangensis(drawn by ZHAO Chuang)
朝阳东方颌翼龙保存不完整,头后部分在发掘中破坏缺失,而头骨在埋藏过程中就已经受到破坏,但是其吻端及牙齿部分保存较好(图6)。基于以下几个方面的特征将东方颌翼龙归入喙嘴龙类翼龙:下颌吻部愈合成短的联合部,向前凸出呈船头状;许多大而犬牙状、伏卧的牙齿构成一个捕鱼的装置(Unwin,2003);吻端没有牙齿(与Rhamphorhynchus muensteri(Wellnhofer,1975,1978;Bennett,1995)以及Bellubrunnus rothgaengeri(Hone et al.,2012)的情况一样);前部的牙齿长且以一定角度伸向前侧方;股骨比肱骨短(与Rhamphorhynchus muensteri(Wellnhofer,1975,1978;Bennett,1995)以及郭氏青龙翼龙的情况一样(Lü et al.,2012)).
朝阳东方颌翼龙两翼展宽1.1 m,第一翼指骨的伸肌突与第一翼指骨、肩胛骨-乌喙骨以及腰带没有愈合现象,说明该个体没有达到性成熟。东方颌翼龙下颌前端的形态介于喙嘴龙和船颌翼龙之间,但更接近于船颌翼龙,而牙齿的形态则类似于喙嘴龙。支序系统学分析表明,该物种属于喙嘴龙类,可能以鱼类为食(图7)。
朝阳东方颌翼龙发现的意义:朝阳东方颌翼龙是目前中国东北晚侏罗世沉积中发现最大的喙嘴龙类翼龙。它的发现填补了中侏罗世到早白垩世翼龙动物群的空白,为以后在晚侏罗世发现更多的翼龙化石提供依据,以及在发现东方颌翼龙的地区进一步工作,指明了方向。
Acknowledgements:
This study was supported by National Natural Science Foundation of China(No.41272022),ChinaGeological Survey(No.12120114026801),and Central Public-interest Scientific Institution Basal Research Fund(No.JB1504).
参考文献:
季强,吕君昌,魏雪芳,王旭日.2012.中国辽西建昌义县组一新的窃蛋龙[J].地质通报,31(12):2012-2017.
吕君昌.2005.中国南方窃蛋龙类化石[M].北京:地质出版社.吕君昌,张宝堃.2005.记中国南方广东省南雄盆地晚白垩世一新的窃蛋龙类化石[J].古生物学报,44(3):412-422.
吕君昌,姬书安,袁崇禧,季强.2006.中国的翼龙类化石[M].北京:地质出版社:147.
吕君昌,金幸生,高春玲,杜天明,丁明,盛益明,魏雪芳.2013.空中之龙——中国翼龙化石研究最新进展[M].杭州:浙江科学技术出版社:127.
徐星,韩凤禄.2010.中国上白垩统窃蛋龙科一新属种(兽脚类:窃蛋龙类)[J].古脊椎动物学报,48(1):11-18.
徐星,汪筱林.2004.辽宁西部早白垩世义县组一新驰龙类[J].古脊椎动物学报,42(2):111-119.
徐星,谭庆伟,王烁,舒柯文,洪大卫,韩凤禄,马擎宇,谭琳,肖东.2013.内蒙古上白垩统窃蛋龙科一新属种及其地层学意义[J].古脊椎动物学报,51(2):85-101.
中国地质科学院.2016.中国地质调查局、中国地质科学院2015年度地质科技十大进展新鲜出炉[J].地球学报,37(1):1-5.
References:
ANDREWS C W.1923.The new conquest of Central Asia:a narrative of the explorations of the Central Asiatic expeditions in Mongolian and China,1921-1930[M].New York:American Museum of Natural History:678.
BALANOFF A M,BEVER G S,NORELL M A.2014.Reconsidering the Avian Nature of the Oviraptorosaur Brain(Dinosauria:Theropoda)[J].PLoS ONE,9(12):e113559.
BALANOFF A M,NORELL M A.2012.Osteology of Khaan mckennai(Oviraptorosauria:Theropoda)[J].Bulletin of the American Museum of Natural History,372:1-77.
BARSBOLD R,OSMÓLSKA H,WATABE M,CURRIE P J,TSOGTBAATAR K.2000.A new oviraptorosaur(Dinosauria,Theropoda)from Mongolia:The first dinosaur with a pygostyle[J].Acta Palaeontologica Polonica,45:97-106.
BARSBOLD R.1997.Oviraptorosauria.Encyclopedia of Dinosaurs[M].CURRIE P J and PADIAN K(eds.).San Diego:Academic Press:505-508.
BENNETT S C.1995.A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany:year classes of a single large species[J].Journal of Paleontology,69:569-580.
BRUSATTE S L,VREMIR M,CSIKI-SAVA Z,TURNER A H,WATANABE A,ERICKSON G M,NORELL M A.2013.The osteology of Balaur bondoc,an island-dwelling dromaeosaurid(Dinosauria:Theropoda)from the Late Cretaceous of Romania[J].Bulletin of the American Museum of Natural History,374:1-100.
Chinese Academy of Geological Sciences.2016.Top Ten Geological Sci-tech Progresses of China Geological Survey(CGS)and Chinese Academy of Geological Sciences(CAGS)in the Year 2015 Unveiled[J].Acta Geoscientica Sinica,37(1):1-5(in Chinese with English abstract).
CHRISTIANSEN P,FARIÑA R A.2004.Mass prediction in theropod dinosaurs[J].Historical Biology,16:85-92.
CLARK J M,NORELL M A,CHIAPPE L M.1999.An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod,Mongolia,preserved in an avian-like brooding position over an oviraptorid nest[J].American Museum Novitates,3265:1-36.
CLARK J M,NORELL M A,ROWE T.2002.Cranial anatomy of Citipati osmolskae(Theropoda,Oviraptorosauria),and a reinterpretation of the holotype of Oviraptor philoceratops[J].American Museum Novitates,3364:1-24.
DECECCHI T A,LARSSON H C E.2013.Body and limb size dissociation at the origin of birds:uncoupling allometric constraints across a macroevolutionary transition[J].Evolution,67:2741-2752.
DONG Z M,CURRIE P J.1996.On the discovery of an oviraptorid skeleton on a nest of eggs at Bayan Mandahu,Inner Mongolia,People’s Republic of China[J].Canadian Journal of Earth Sciences,33:631-636.
ELZANOWSKI A.1999.A comparison of the jaw skeleton in theropods and birds,with a description of the palate in the Oviraptoridae[J].Smithsonian Contributions to Paleobiology,89:311-323.
EVANS D C,LARSON D W,CURRIE P J.2013.A new dromaeosaurid(Dinosauria:Theropoda)with Asian affinities from the latest Cretaceous of North America[J].Naturwissenschaften,100:1041-1049.
FEDUCCIA A,CZERKAS S A.2015.Testing the neoflightless hypothesis:propatagium reveals flying ancestry of oviraptorosaurs[J].Journal of Ornithology,156:1067-1074.
FEDUCCIA A.2012.Riddle of the feathered dragons[M].New Haven:Yale University Press.
FOTH C,TISCHLINGER H,RAUHUT O W M.2014.New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers[J].Nature,511:79-82.
GODEFROIT P,DEMUYNCK H,DYKE G,HU D Y,ESCUILLIÉ F,CLAEYS P.2013.Reduced plumage and flight ability of a new Jurassic paravian theropod from China[J].Nature Communications,4:1394 doi:10.1038/ncomms2389.
HAN G,CHIAPPE L M,JI S A,HABIB M,TURNER A H,CHINSAMY A,LIU X L,HAN L Z.2014.A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid flight performance[J].Nature Communications,5:4382,doi:10.1038/ncomms5382.
HOLTZ T R J.2000.A new phylogeny of the carnivorous dinosaurs[J].Gaia,15:5-61.
HONE D W E,TISCHLINGER H,FREY E,ROPER M.2012.A new non-pterodactyloid pterosaur from the Late Jurassic of Southern Germany[J].PLoS ONE,7:e39312.
HU D,HOU L H,ZHANG L,XU X.2009.A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus[J].Nature,461:640-643.
JI Q,CURRIE P J,NORRELL M A,JI S A.1998.Two feathereddinosaurs from northeastern China[J].Nature,393:753-761.
JI Q,NORELL M A,GAO K Q,JI S A,REN D.2001.The distribution of integumentary structures in a feathered dinosaur[J].Nature,410:1084-1088.
JI Qiang,LÜ Jun-chang,WEI Xue-fang,WANG Xu-ri.2012.A new oviraptorosaur from the Yixian Formation of Jianchang,western Liaoning Province,China[J].Geological Bulletin of China,31:2102-2107(in Chinese with English abstract).
KIRKLAND J I,BURGE D,GASTON R.1993.A large dromaeosaur(Theropoda)from the Lower Cretaceous of eastern Utah[J].Hunteria,2(10):1-16.
LAMANNA M C,SUES H D,SCHACHNER E R,LYSON T R.2014.A new large-bodied oviraptorosaurian theropod dinosaur from the Latest Cretaceous of Western North America[J].PLoS ONE,9:e92022.
LI Q,GAO K Q,MENG Q J,CLARKE J A,SHAWKEY M D,D'ALBA L,PEI R,ELLISON M,NORELL M A,VINTHER J.2012.Reconstruction of Microraptor and the evolution of iridescent plumage[J].Science,335:1215-1219.
LONGRICH N R,CURRIE P J,DONG Z M.2010.A new oviraptorid(Dinosauria:Theropoda)from the Upper Cretaceous of Bayan Mandahu,Inner Mongolia[J].Palaeontology,53:945-960.
LONGRICH N R.VINTHER J,MENG Q J,LI Q G,RUSSELL A P.2012.Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi[J].Current Biology,22:1-6.
LÜ J C,BRUSATTE S L.2015.A large,short-armed,winged dromaeosaurid(Dinosauria:Theropoda)from the Early Cretaceous of China and its implications for feather evolution[J].Scientific Reports,5:11775,doi:10.1038/srep11775.
LÜ J C,CURRIE P J,XU L,ZHANG X L,PU H Y,JIA S H.2013b.Chicken-sized oviraptorid dinosaurs from central China and their ontogenetic implications[J].Naturwissenschaften,100:165-175.
LÜ J C,DONG Z,AZUMA Y,BARSBOLD R,TOMIDA Y.2002.Oviraptorosaurs compared to birds[M]//Zhou Z,Zhang F(eds)Proceedings of the 5thSymposium of the Society of Avian Paleontology and Evolution.Beijing:Science Press:175-189.
LÜ J C,HONE D W E.2012.A new Chinese anurognathid pterosaur and the evolution of Pterosaurian tail lengths[M].Acta Geologica Sinica,86(6):1317-1325.
LÜ J C,PU H Y,KOBAYASHI Y,XU L,CHANG H L,SHANG Y H,LIU D,LEE Y-N,KUNDRÁT M,SHEN C Z.2015a.A new oviraptorid dinosaur(Dinosauria:Oviraptorosauria)from the Late Cretaceous of Southern China and its paleobiogeographical implications[J].Scientific Reports,5:11490,doi:10.1038/srep11490.
LÜ J C,PU H Y,XU L,WEI X F,CHANG H L,KUNDRÁT M.2015b.A new rhamphorhynchid pterosaur(Pterosauria)from Jurassic deposits of Liaoning Province,China[J].Zootaxa,3911(1):119-129.
LÜ J C,TOMIDA Y,AZUMA Y,DONG Z M,LEE Y N.2004.New oviraptorid dinosaur(Dinosauria:Oviraptorosauria)from the Nemegt Formation of southwestern Mongolia[J].Bulletin of the National Science Museum,Tokyo,Series C 30:95-130.
LÜ J C,TOMIDA Y,AZUMA Y,DONG Z M,LEE Y N.2005.Nemegtomaia gen.nov.,a replacement name for the oviraptorosaurian dinosaur Nemegtia Lü et al.,2004,a preoccupied name[J].Bulletin of the National Science Museum,Tokyo,Series C:31,51.
LÜ J C,UNWIN D M,JIN X S,LIU Y Q,JI Q.2010.Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull[J].Proceedings of the Royal Society B,227:383-389.
LÜ J C,UNWIN D M,ZHAO B,GAO C L,SHEN C Z.2012.A new rhamphorhynchid(Pterosauria:Rhamphorhynchidae)from the Middle/ Upper Jurassic of Qinglong,Hebei Province,China[M].Zootaxa,3158:1-19.
LÜ J C,XU L,JIANG X J,JIA S H,LI M,YUAN C X,ZHANG X L,JI Q.2009.A preliminary report on the new dinosaurian fauna from the Cretaceous of the Ruyang Basin,Henan Province of central China[J].Journal of Palaeontological Society of Korea,25:43-56.
LÜ J C,YI L P,ZHONG H,WEI X F.2013a.A new oviraptorosaur(Dinosauria:Oviraptorosauria)from the Late Cretaceous of Southern China and its paleoecological implications[J].PLoS ONE,8:e80557.
LÜ J C.2000.Oviraptorosaurs compared to birds.In:L.Shi and F.Zhang(eds.),5th International Meeting of the Society of Avian Paleontology and Evolution and the Symposium on Jehol Biota[J].Vertebrata PalAsiatica,38(Supplement):18.
LÜ J C.2002.A new oviraptorosaurid(Theropoda:Oviraptorosauria)from the Late Cretaceous of southern China[J].Journal of Vertebrate Paleontology,22:871-875.
LÜ Jun-chang,JI Shu-an,YUAN Chong-xi,JI Qiang.2006.Pterosaurs from China[M].Beijing:Geological Publishing House(in Chinese).
LÜ Jun-chang,JIN Xing-sheng,GAO Chun-ling,DU Tian-ming,DING Ming,SHENG Yi-ming,WEI Xue-fang.2013.Dragons of the skies—Recent advances on the study of pterosaurs from China[M].Hangzhou:Zhejiang Science and Technology Press(in Chinese).
LÜ Jun-chang,ZHANG Bao-kun.2005.A new oviraptorid(Theropoda:Oviraptorosauria)from the Upper Cretaceous of the Nanxiong Basin,Guangdong Province of southern China[J].Acta Palaeontologica Sinica,44:412-422(in Chinese with English abstract).
LÜ Jun-chang.2005.Oviraptorid dinosaurs from Southern China[M].Beijing:Geological Publishing House(in Chinese).
MARYAŃSKA T,OSMÓLSKA H,WOLSAN M.2002.Avialan status for Oviraptorosauria[J].Acta Palaeontologica Polonica,47(1):97-116.
MARYAŃSKA T,OSMÓLSKA H.1997.The quadrate of oviraptorid dinosaurs[J].Acta Palaeontologica Polonica,42:361-371.
MAYR G,POHL B,HARTMAN S,PETERS D S.2007.The tenth skeletal specimen of Archaeopteryx[J].Zoological Journal of the Linnean Society,149:97-116.
NORELL M A,CLARK J M,CHIAPPE L M,CHIAPPE L M.2001.An Embryonic Oviraptorid(Dinosauria:Theropoda)from the Upper Cretaceous of Mongolia[J].American Museum Novitates,3315:1-12.
NORELL M A,CLARK J M,CHIAPPE L M,DASHZEVEG D.1995.A nesting dinosaur[J].Nature,378:774-776.
NORELL M A,CLARK J M,DASHZEVEG D,BARSBOLD R,CHIAPPE L M,DAVIDSON A R,MC-KENNA M C,ALTANGEREL P,NOVACEK M J.1994.A theropod dinosaur embryo,and the affinities of the Flaming Cliffs Dinosaur eggs[J].Science,266:779-782.
NORELL M A,MAKOVICKY P J.2004.Dromaeosauridae[M]// WEISHAMPEL D B,DODSON P,OSMÓLSKA H(eds).The Dinosauria,2nd edn.Berkeley:University of California Press:196-209.
NOVAS F E,POL D,CANALE J I,PORFIRI J D,CALVO J O.2009.A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids[J].Proceedings of the Royal Society of London B,276:1101-1107.
OSBORN H F.1924.Three new theropoda,protoceratops zone,central Mongolia[J].American Museum Novitates,144:1-12.
OSMÓLSKA H,CURRIE P J,BRASBOLD R.2004.Oviraptorosauria[M]//The Dinosauria.2nd edn(eds Weishampel D,Dodson P,Osmólska H)Ch.8.California:University of California Press:165-183.
SENTER P,KIRKLAND J I,DEBLIEUX D D,MADSEN S,TOTH N.2012.New dromaeosaurids(Dinosauria:Theropoda)from the Lower Cretaceous of Utah,and the Evolution of the dromaeosaurid Tail[J].PLoS ONE,7(5):e36790.doi:10.1371/journal.pone.0036790.
SERENO P C.1999.The evolution of dinosaurs[J].Science,284:2137-2147.
TURNER A H,MAKOVICKY P J,NORELL M A.2012.A review of dromaeosaurid systematics and paravian phylogeny[J].Bulletin of the American Museum of Natural History,371:1-206.
TURNER A H,POL D,NORELL M A.2011.Anatomy of Mahakala omnogovae(Theropoda:Dromaeosauridae),Tögrögiin Shiree,Mongolia[J].American Museum Novitates,3722:1-66.
UNWIN D M.2003.On the phylogeny and evolutionary history of pterosaurs:In:Buffetaut,E.&Mazin,J.M.(Eds.),Evolution and Palaeobiology of Pterosaurs[M].London:Geological Society:139-190.
WANG S,SUN C,SULLIVAN C,XU X.2013.A new oviraptorid(Dinosauria:Theropoda)from the Upper Cretaceous of southern China[J].Zootaxa,3640:242-257.
WEI X F,PU H Y,XU L,LIU D,LÜ J C.2013.A new oviraptorid dinosaur(Theropoda:Oviraptorosauria)from the Late Cretaceous of Jiangxi Province,southern China[J].Acta Geologica Sinica,87:899-904.
WELLNHOFER P.1975.Die Rhamphorhynchoidea(Pterosauria)der oberjura-plattenkalke süddeutschlands.Teil I.allgemeine skelletmorphologie[J].Paläontographica A,148:1-33.
WELLNHOFER P.1978.Handbuch der paläoherpetologie.Teil 19.Pterosauria[M].Stuttgart:Verlag Gustav Fischer:82.
WELLNHOFER P.2008.Archaeopteryx[M].Munchen:Der Urvogel von Solnhofen.
XU X,CHENG Y N,WANG X L,CHANG C H.2002a.An unusual oviraptorosaurian dinosaur from China[J].Nature,419:291-293.
XU X,NORELL M A,WANG X L,MAKOVICKY P J,WU X C.2002b.A basal troodontid from the Early Cretaceous of China[J].Nature,415:780-784.
XU X,TAN Q W,WANG J M,ZHAO X J,TAN L.2007.A gigantic bird-like dinosaur from the Late Cretaceous of China[J].Nature,447:844-847.
XU X,WANG X L,WU X C.1999.A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China[J].Nature,401:262-266.
XU X,WU X C.2001.Cranial morphology of Sinornithosaurus millenii Xu et al.1999(Dinosauria:Theropoda:Dromaeosauridae)from the Yixian Formation of Liaoning,China[J].Canadian Journal of Earth Sciences,38:1739-1752.
XU X,ZHOU Z H,WANG X L,KUANG X W,ZHANG F C,DU X K.2003.Four-winged dinosaurs from China[J].Nature,421:335-340.
XU X,ZHOU Z H,WANG X L.2000.The smallest known non-avian theropod dinosaur[J].Nature,408:705-708.
XU Xing,HAN Feng-lu.2010.A new oviraptorid dinosaur(Theropoda:Oviraptorosauria)from the Upper Cretaceous of China[J].Vertebrata PalAsiatic,48(1):11-18(in Chinese with English abstract).
XU Xing,TAN Qing-wei,WANG Shuo,SULLIVAN C,HONE D W E,HAN Feng-lu,MA Qing-yu,TAN Lin,XIAO Dong.2013.A new oviraptorid from the Upper Cretaceous of Nei Mongol,China,and its stratigraphic implications[J].Vertebrata PalAsiatic,51:85-101.
XU Xing,WANG Xiao-lin.2004.A new dromaeosaur(Dinosauria:Theropoda)from the Early Cretaceous Yixian Formation of Western Liaoning[J].Vertebrata PalAsiatic,42:111-119(in Chinese with English abstract).
ZANNO L E,SAMPSON S D.2005.A new oviraptorosaur(Theropoda;Maniraptora)from the Late Cretaceous(Campanian)of Utah[J].Journal of Vertebrate Paleontology,25:897-904.
ZELENITSKY D K,THERRIEN F,ERICKSON G M,DEBUHR C L,KOBAYASHI Y,EBERTH D A,HADFIELD F.2012.Feathered non-avian dinosaurs from North America provide insight into wing origins[J].Science,338:510-514.
ZHENG X T,XU X,YOU H L,ZHAO Q,DONG Z M.2010.A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution[J].Proceedings of the Royal Society of London B,277:211-217.
New Findings in the Study of Feathered Dinosaurs and Pterosaurs
LÜ Jun-chang
Institute of Geology,Chinese Academy of Geological Sciences,Beijing 100037;
Key Lab of Stratigraphy and Paleontology,Ministry of Land and Resources,Beijing 100037
Abstract:An oviraptorid dinosaur Huanansaurus ganzhouensis,belonging to the derived oviraptorid dinosaur,was discovered from the Late Cretaceous deposits in Ganzhou,Jiangxi Province.Its unique skull characteristics provide a new insight into the craniofacial evolution of oviraptorosaurid dinosaurs,paleogeographical distribution and paleoecological environments.Zhenyuanlong suni from the Early Cretaceous Yixian Formation of western part of Liaoning Province is a large,short-armed,winged dromaeosaurid dinosaur.It provides the evidence of feather morphologies and distribution in a short-armed dromaeosaurid dinosaur.The discovery of Zhenyuanlong also provides important evidence for the study of the diversity,the origin of feathers and the flight of Liaoning dromaeosaurid dinosaurs.Orientognathus chaoyngensis is the first Late Jurassic pterosaur discovered in western Liaoning and it is the largest rhamphorhynchinae pterosaur.The discovery of Orientognathus not only fills the temporal gap between the Middle Jurassic and Early Cretaceous pterosaur faunas of China but also plays an important role in the understanding of radiation evolution of rhamphorhynchids from the Late Jurassic.The above-mentioned important discoveries play a key role in our understanding of the hot and difficult issues such as the evolution of oviraptorid dinosaurs,the feather evolution in dromaeosaurid dinosaurs and the origin of bird feathers.In particular,the Ganzhou Dinosaurian Fauna is named based on the discovery of Huanansaurus.
Key words:Huanansauurs;Late Cretaceous;Ganzhou of Jiangxi Province;Zhenyuanlong;Early Cretaceous;Orientognathus;Late Jurassic;western part of Liaoning Province
作者简介:第一吕君昌,男,1965年生。研究员,博士生导师。主要从事中生代爬行动物(尤其是恐龙类和翼龙类)及其地层学研究。
通讯地址:100037,北京市西城区百万庄大街26号。电话:010-68999707。E-mail:Lujc2008@126.com。
收稿日期:2016-02-15;改回日期:2016-02-22。责任编辑:闫立娟。
中图分类号:Q915.2;Q915.86
文献标志码:A
doi:10.3975/cagsb.2016.02.01
