舟山群岛海域网采浮游植物分布与多样性
2016-04-23刘晶晶江志兵曾江宁
陈 悦,刘晶晶,江志兵,黄 伟,寿 鹿*,曾江宁
(1. 国家海洋局 海洋生态系统与生物地球化学重点实验室,浙江 杭州 310012;
2. 国家海洋局 第二海洋研究所,浙江 杭州 310012)
舟山群岛海域网采浮游植物分布与多样性
陈悦1,2,刘晶晶1,2,江志兵1,2,黄伟1,2,寿鹿*1,2,曾江宁1,2
(1. 国家海洋局 海洋生态系统与生物地球化学重点实验室,浙江 杭州 310012;
2. 国家海洋局 第二海洋研究所,浙江 杭州 310012)
摘要:于2014年夏、秋两季在舟山海域进行了浮游植物拖网采集和环境理化因子测定,共鉴定浮游植物6门92属184种,其中硅藻门127种,甲藻门30种,为调查海域主要浮游植物类群。夏季骨条藻Skeletonema spp.为绝对优势种,秋季优势种由琼氏圆筛藻Coscinodiscus jonesianus、虹彩圆筛藻Coscinodiscus oculus-iridis、蛇目圆筛藻Coscinodiscus argus、星脐圆筛藻Coscinodiscus asteromphalus、骨条藻Skeletonema spp.、尖刺伪菱形藻Pseudo-nitzschia pungens、并基角毛藻Chaetoceros decipiens、伏氏海毛藻Thalassiothrix frauenfeldii及中间肋缝藻Frustulia interposita等9种硅藻构成。调查海域夏季浮游植物平均丰度(1 713.58×104个/m3)高于秋季(1 430.30×104个/m3)。冗余分析表明,温度、盐度、悬浮物、溶解无机磷及溶解无机氮等环境因子对夏、秋两季浮游植物群落影响较大。结合历史数据分析可知,舟山海域近30 a来富营养化程度严重且日益加剧,富营养化指数明显升高;同时浮游植物群落变化也十分显著,主要体现为细胞丰度增加和优势种更替。
关键词:舟山群岛;浮游植物;群落特征;环境因子
0引言
舟山群岛位于我国东部海岸线和长江出海口交界,陆地面积1 440 km2,内海海域面积2.08×104km2,是我国首个以海洋经济为主题的国家战略层面新区,自2011年3月14日批准成为中国第一个群岛新区以来,舟山群岛新区凭借其重要的地理位置,丰富的海洋资源已成为浙江省海洋开发的重点岛群[1]。然而,由于长江、钱塘江等陆地径流的影响,大量营养盐的输入使得舟山群岛呈现富营养化的状态[2]。
浮游植物作为海洋生态系统中最主要的初级生产者,其群落结构以及生物量的改变都将通过食物网影响到海洋生态系统的功能和结构[3-4]。与此同时,浮游植物又凭借其对环境的敏感性对海洋生态系统的变化起着指示性作用[5-7]。过去几十年,对舟山海域浮游植物群落结构及生物量的调查从未间断,然而以往调查基本是针对舟山群岛某特定区域(如舟山渔场附近海域[8-10]、舟山-宁波港[11])的浮游植物或是针对该海域某些特定藻种(如赤潮藻种[12])的影响研究,对于舟山群岛及其周围海域(包括金塘岛和册子岛)浮游植物的研究则相对较少。鉴于此,本文根据2014年7月(夏季)和11月(秋季)2个航次所采集到的浮游植物样品,分析了舟山群岛新区浮游植物的种类组成、优势种、丰度分布和多样性指数等,并通过冗余分析法分析了环境因子对群落结构的影响,另外还与历史资料进行了对比分析,以期对该海域浮游植物群落结构变化有更为全面的认识。
1材料与方法
1.1调查海域与站位设置
于2014年7月29日至8月1日和2014年11月9日至11月11日,对舟山群岛海域进行了2个航次的调查。根据舟山群岛的岛群分布,环舟山本岛及册子、金塘两岛共布设25个采样站位(图1)。
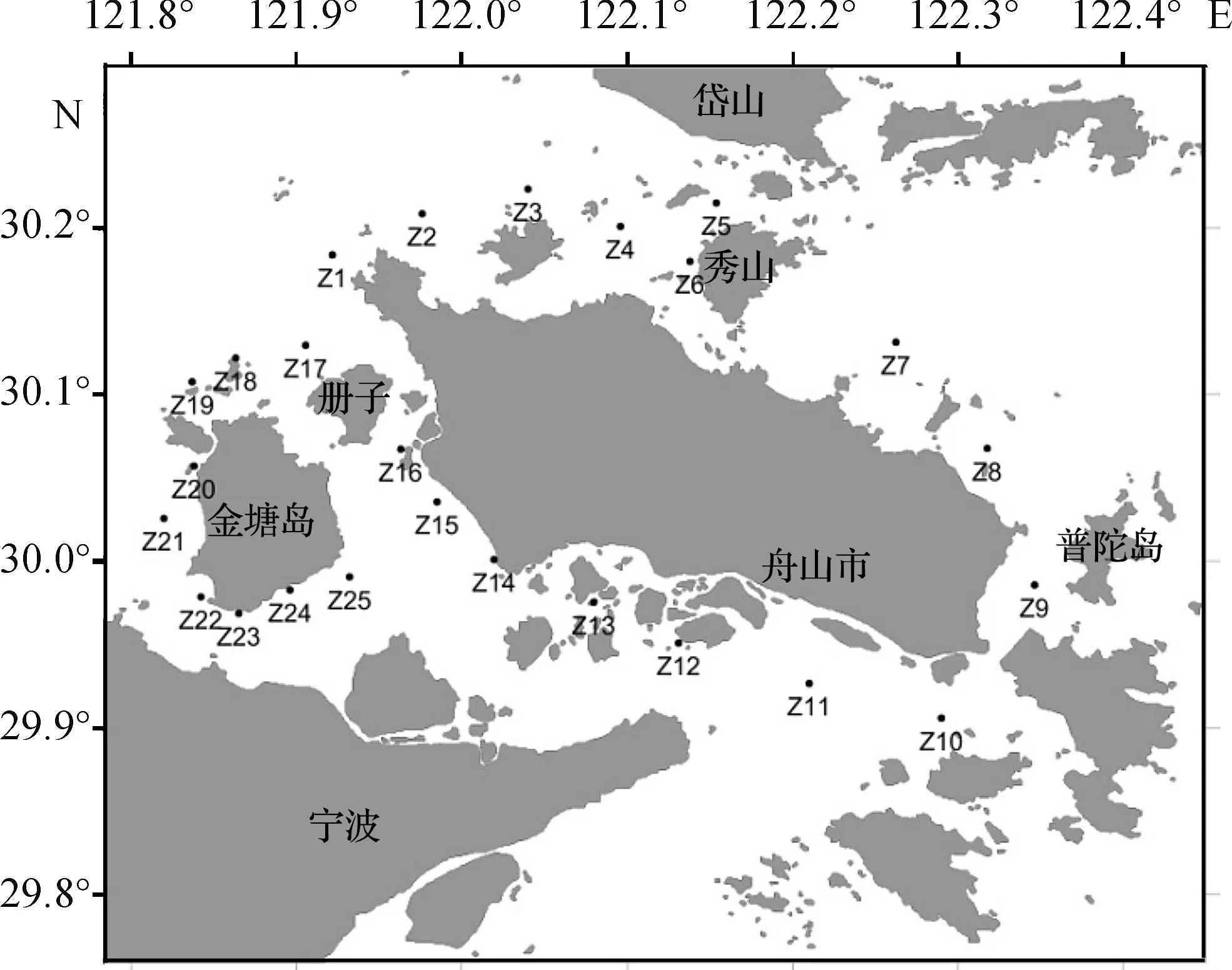
图1 舟山群岛海域采样站位Fig.1 Sampling stations in the waters nearby Zhoushan Islands
1.2样品采集与分析
根据《海洋监测规范》[13],采用网目为76 μm的浮游生物网自底至表进行垂直拖网采集,并用Hydro-bios流量计记录滤水量。所采集样品用4%中性甲醛进行固定,但夏季航次因水流原因未采集Z4、Z6和Z7站位的浮游植物样品。经浓缩制片后用ZEISS Scope A1显微镜进行观察、鉴定和计数。采集表层(0.5 m)水样测定得到盐度、COD、溶解无机氮(DIN)以及溶解无机磷(DIP)等理化数据。
1.3数据分析及处理
根据浮游植物群落结构指数的分析方法[14-15],采用以下公式进行计算:
Shannon-Wiener指数(H′):
(1)
Pielou均匀度指数(J):
(2)
Margalef丰度指数(d):
(3)
优势度(Y):
Y=(ni/N)fi
(4)
式中:ni为第i种的个体数;N为总个体数;Pi为第i种生物量与样方中总生物量的比值;S为样品中种类的总数;fi为第i种在各样品当中出现的频率,如果某物种的Y≥2%时,则定为优势种。
本文采用营养指数法计算海域富营养化指数[16],公式如下:
(5)
式中:COD,无机氮和无机磷含量的单位均为mg/L,E≥1时,即可认为水体呈富营养化,且E与水体富营养化程度呈正比。
采用Canoco4.5软件对浮游植物群落和环境进行除趋势对应分析(DetrendedCorrespondenceAnalysis,DCA),分析结果表明各轴梯度最大值小于3,故采用冗余分析(RedundanceAnalysis,RDA)。在RDA排序图中,箭头表示环境因子,箭头连线的长短表示各站位浮游植物群落与环境因子相关性的大小。各站位的藻类群落结构指数使用PRIMER6.0软件进行处理,在处理前,先对数据进行四次方根转化。
2结果
2.1群落组成
2次调查共鉴定浮游植物6门92属184种,其中硅藻49属127种(占69.0%),甲藻18属30种(占16.3%),绿藻13属14种(占7.6%),蓝藻8属9种(占4.9%),裸藻和隐藻各2属2种(各占1.1%)。
夏季共鉴定浮游植物6门64属105种,其中硅藻40属74种(占70.4%),甲藻10属17种(占16.2%),绿藻7属7种(占6.7%),蓝藻3属3种(占2.9%),隐藻和裸藻各2属2种(各占1.9%);秋季共鉴定浮游植物5门55属106种,其中硅藻38属82种(占77.3%),甲藻9属16种(占15.1%),绿藻5属5种(占4.7%),蓝藻2属2种(占1.9%),裸藻1属1种(占1.0%)。
2.2优势种组成
本次调查,舟山海域夏季浮游植物优势种有7种,包括并基角毛藻Chaetoceros decipiens、蛇目圆筛藻Coscinodiscus argus、琼氏圆筛藻Coscinodiscus jonesianus、虹彩圆筛藻 Coscinodiscus oculus-iridis、尖刺伪菱形藻Pseudo-nitzschia pungens、骨条藻Skeletonemaspp.以及裸藻Euglenasp.,其中骨条藻占绝对优势(Y=0.31)。秋季优势种有9种,包括并基角毛藻、蛇目圆筛藻、星脐圆筛藻Coscinodiscus asteromphalus、虹彩圆筛藻、琼氏圆筛藻、中间肋缝藻Frustulia interposita、尖刺伪菱形藻、骨条藻以及伏氏海毛藻Thalassiothrix frauenfeldii;各优势种的优势度较为平均(表1)。
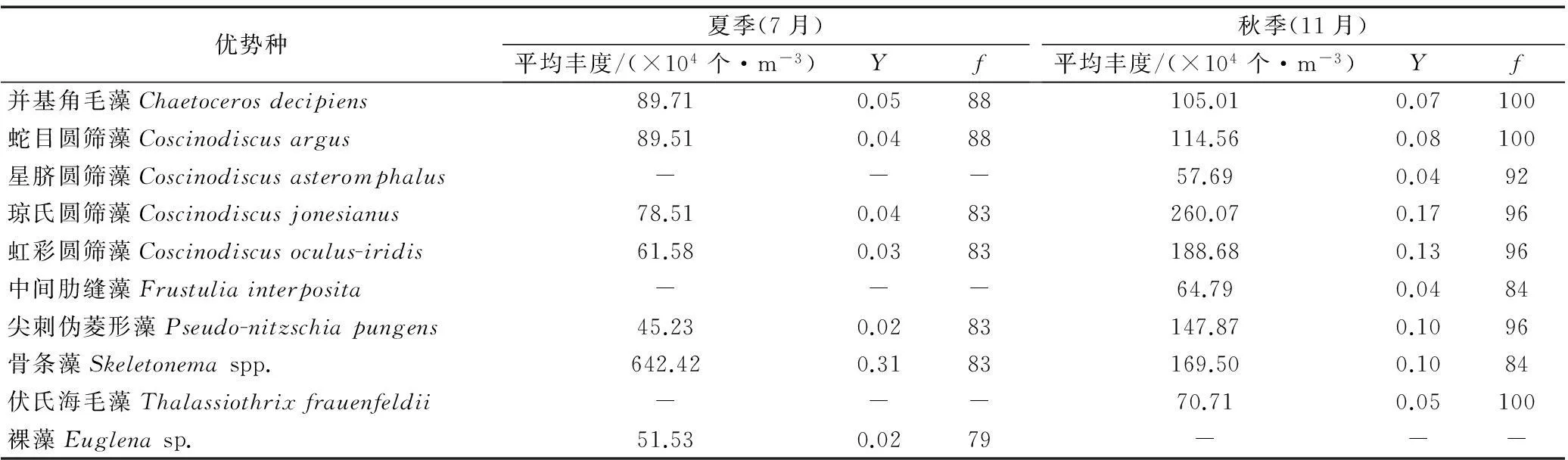
表1 夏、秋两季浮游植物优势种平均丰度、优势度(Y)和频率(f)
注:“-”表示非优势种
2.3细胞丰度分布
夏季,调查海域浮游植物细胞丰度为18.94×104~5 996.67×104个/m3,平均细胞丰度为1 713.58×104个/m3;秋季为195.06×104~ 3 764.91×104个/m3,平均细胞丰度为1 430.30×104个/m3。两季浮游植物群落均以硅藻占绝对优势(分别占总丰度的81.2%和98.5%)、甲藻次之(分别占总丰度的5.1%和0.6%),且夏季甲藻比例(5.1%)高于秋季(0.6%),可见夏季调查海域浮游植物类群以硅藻和甲藻为主,秋季硅藻占绝对优势(图2)。
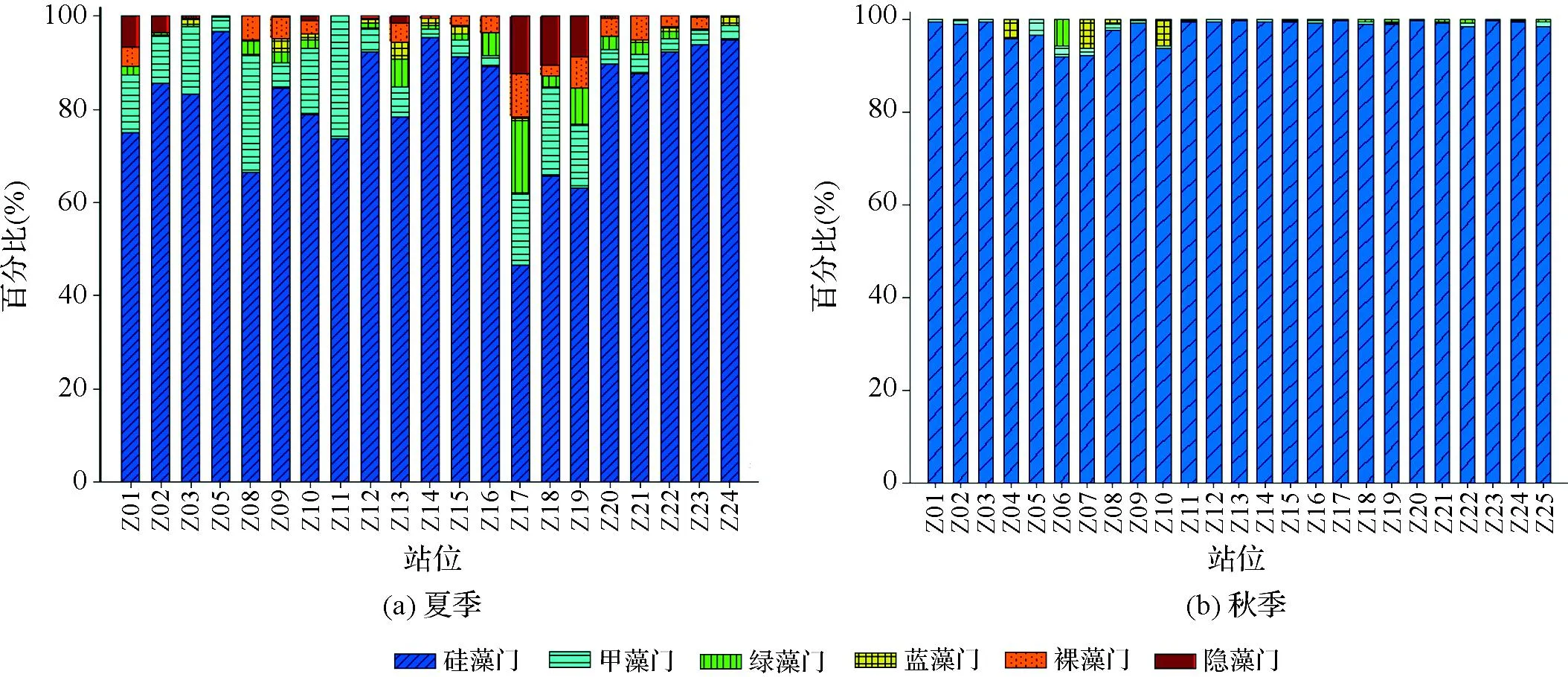
图2 夏、秋两季各站位浮游植物类群的比例Fig.2 The ratio of phytoplankton genera at different stations in summer and autumn
2.4群落多样性分析
夏季调查海域浮游植物的多样性指数为0.92~2.94,平均值为2.52,最低值出现在Z25站位;秋季为3.04~3.58,平均为3.28,最低值出现在Z5站位。夏季丰富度指数、均匀度指数和多样性指数(分别为4.33、0.69、2.52)均低于秋季(分别为6.92、0.98、3.28)(表2)。

表2 夏、秋两季调查海域平均丰富度指数(d)、均匀度指数(J)
2.5理化因子
夏季舟山海域海水中溶解无机氮平均质量浓度为0.65±0.18 mg/L、无机磷为0.038 0±0.007 2 mg/L;秋季溶解无机氮和无机磷含量分别为1.07±0.17和0.051 4±0.005 8 mg/L(表3)。根据中华人民共和国海水水质标准(GB 3097—1997)[17],舟山群岛附近海域溶解无机氮含量超过四类水标准;夏季溶解无机磷含量属于四类水范围,秋季则超四类水范围。根据公式5可计算得到夏季富营养化指数E为4.79,秋季为21.04,因此,调查海域水质富营养化程度严重。
此外,本次调查夏季平均水深、温度、盐度、pH及悬浮物含量分别为16.00±6.38 m、25.44±1.06 ℃、25.06±2.46、7.99±0.06和135.83±146.38 mg/L,秋季分别为12.92±5.37 m、20.17±0.45 ℃、21.21±2.13、8.08±0.01和652.00±377.73 mg/L。

表3 夏、秋两季舟山群岛附近海域环境理化因子(平均值±标准差)
2.6相关性分析
除pH外,其他水质参数(水深、温度、盐度、悬浮物、溶解无机磷及溶解无机氮)在夏、秋两季均有显著差异,其中悬浮物含量差异十分明显,秋季(652.00±377.73 mg/L)比夏季(135.83±146.38 mg/L)高出近4倍。夏、秋两季RDA分析中的6个环境因子分别可解释各站位浮游植物群落总变量的36.8%及34.2%。夏季第1轴和第2轴的特征值分别为0.138和0.073,分别解释了37.4%和19.9%的站位丰度变量,站位群落-环境相关系数分别为0.907和0.846;秋季第1轴和第2轴的特征值分别为0.120和0.079,分别解释了35.2%和22.9%的站位丰度变量,站位群落-环境相关系数分别为0.895和0.868。表明这6个环境因子与两季各站位浮游植物群落均存在较好的相关性(图3)。
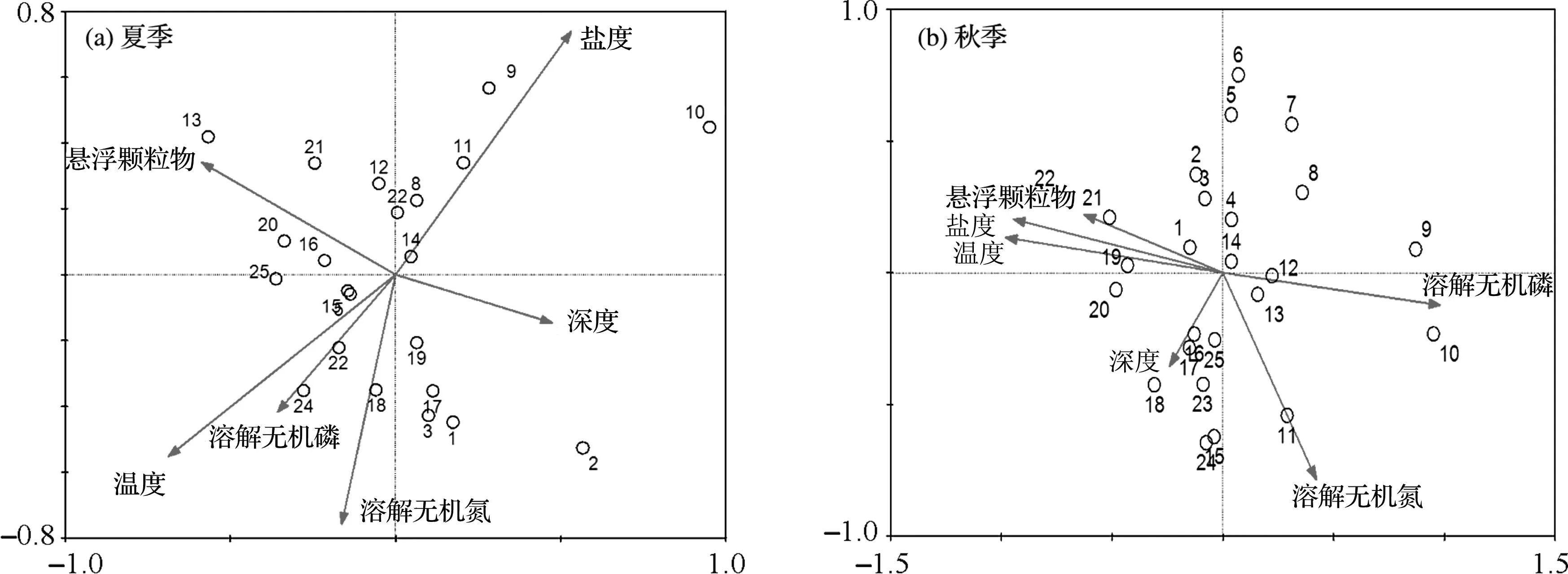
图3 夏、秋两季各站位浮游植物丰度与环境因子间的RDA排序图Fig.3 RDA ordination of phytoplankton abundance and environmental factors at different stations in summer and autumn
3讨论
3.1群落结构与环境因子的关系
调查期间,舟山海域主要受长江冲淡水、台湾暖流水影响[9],长江冲淡水呈现低盐特征,台湾暖流则呈现高盐的外海水特征,两个水团交汇,使得该海域盐度适中,适合浮游植物大量繁殖[18]。因此,该海域浮游植物平均丰度一直处于较高水平且种类复杂,不仅有长江径流带入的盘星藻Pediastrum、桥弯藻Cymbella等淡水种及骨条藻等半咸水种,台湾暖流带入的拟旋链角毛藻Chaetocerospseudocurvisetus等常见于暖水的近岸种,还有较多如大角角藻Ceratiummacroceros等外海水携带而来的外海高盐种[6,19]。同时,该海域受到两水团交汇的影响,近外海侧盐度较高适应于咸水种,近冲淡水径流处适应于半咸水种及少量淡水种,这种浮游植物类群对海水盐度适应性不同可能造成了各站位群落差异(图2)。
调查海域营养盐来源一方面由长江冲淡水携带大量陆源营养,另一方面台湾暖流也为浮游植物提供了丰富的磷酸盐[20],因此2个调查季节的营养盐均较丰富,夏季溶解无机氮及溶解无机磷含量分别为0.65和0.038 0 mg/L、秋季分别为1.07和0.051 4 mg/L(表3),不构成该海域浮游植物生长的最主要限制因子。
两季调查期间,海水悬浮物含量均较高,夏、秋两季分别为135.83和652.00 mg/L(表3),这可能是由于舟山群岛海域处于混浊区。黄小平 等[21]指出最大浑浊带附近会形成重力环流, IRIGOIEN et al[22]、JONGE et al[23]研究表明,受浑浊带重力环流作用,微型底栖硅藻会再悬浮至表面,而再悬浮的硅藻对该区域的藻类群落有显著影响;同时,该环流会富集来自冲淡水及外海水两个方向滞留的颗粒物,这种“捕捉效应”集聚了部分浮游植物的同时也延长了其在混浊带区域的停留时间[21]。此外,高悬浮物质量浓度使得该海域水体浑浊,透明度较低,水环境由于径流量较大而极不稳定,多数种类很难生长,而骨条藻等适应于大径流量水域环境的藻种大量繁殖成为该海域的优势种[24],这与前人的研究结论相符合[ 8,19,25]。
因此,舟山海域浮游植物群落受到悬浮物含量、盐度、溶解无机氮、溶解无机磷和重力环流等诸多环境因素的综合影响(图3),丰度较高且群落结构复杂。
3.2多样性分析
陈碧鹃 等[26]指出,浮游植物群落的多样性指数(H′)和均匀度指数(J)可以作为环境评价的依据。根据《水生生物监测手册》[27],当0
此外,根据PINKNEY[30]的观点,当调查区域优势种较多且各优势种之间优势度相近,无绝对优势种,则此时群落结构更加稳定,因此,舟山海域秋季浮游植物群落稳定性优于夏季。
3.3舟山海域浮游植物长期变化
近30 a来,舟山海域浮游植物优势种构成变化显著(表4)。自20世纪80年代初以来,硅藻一直占据绝对优势,但20世纪80年代初及90年代初主要优势属为圆筛藻属,进入21世纪,骨条藻开始占据绝对优势,2008年优势种仅为中肋骨条藻,直至本次调查,骨条藻依然处于优势种地位。此外,1982年舟山海域细胞丰度仅为212.5×104个/m3,至此次调查,平均丰度最高已达1 713×104个/m3,期间历年数据显示,该海域细胞丰度大致处于上升趋势。
根据刘雪芹[2]的结论推测这种优势种的演替及浮游植物细胞丰度爆发性增长与舟山近年来富营养化日益严重有关,舟山海区自1996年(E=3.88)至2000年(E=11.49)富营养化指数上升近3倍,本次调查秋季富营养化指数达到21.04,说明舟山海域一直处于严重富营养化状态且富营养化日趋严重[16]。营养盐不再成为约束浮游植物生长的条件,导致其大量繁殖;而中肋骨条藻属于赤潮藻种,海域的富营养化为该种提供了繁殖所需的大量营养盐,因此骨条藻大量生长,自21世纪来一直处于优势地位。
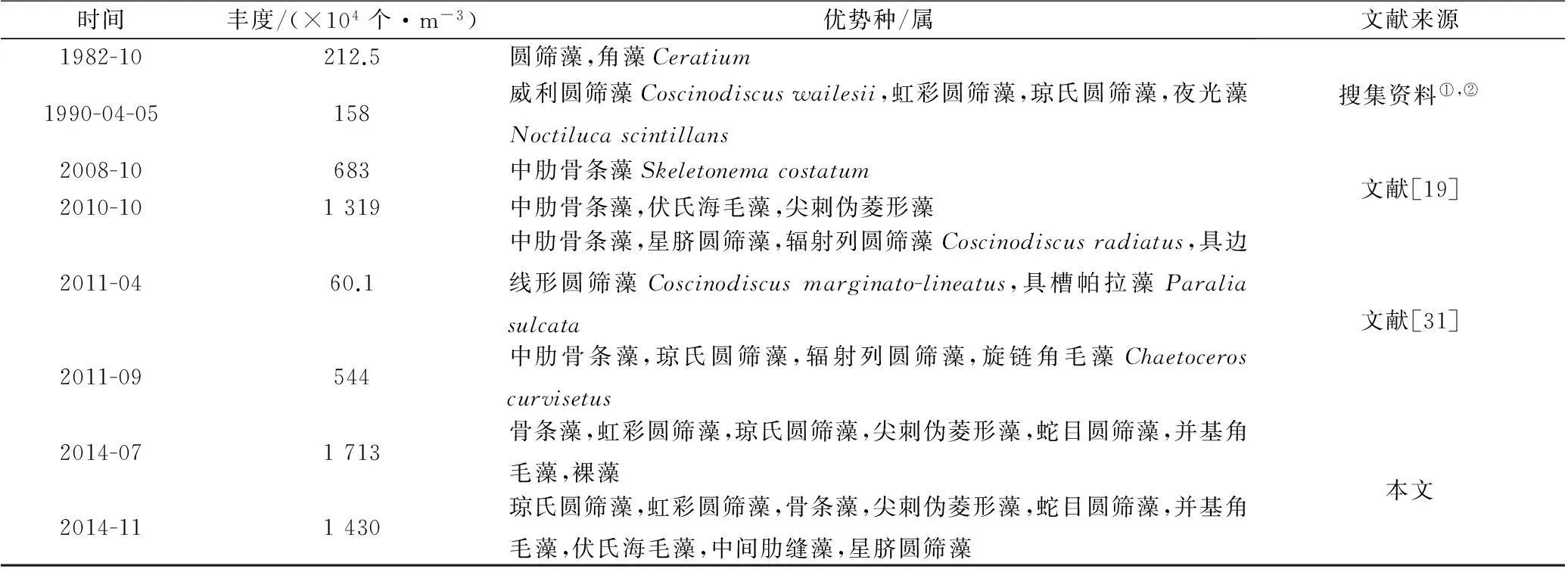
表4 舟山群岛附近海域网采浮游植物丰度和优势种/属的变化
注:①国家海洋局第二海洋研究所,浙江省海岸带资源综合调查专业报告,1985;②国家海洋局第二海洋研究所,浙江省海岛调查报告,1993
4结论
本文于2014年夏、秋两季在舟山海域进行了2个航次的调查,得到以下结论:
(1)夏、秋两季共鉴定出浮游植物6门92属184种,其中,硅藻49属127种(占69.0%),甲藻18属30种(占16.3%),以骨条藻、蛇目圆筛藻、琼氏圆筛藻等半咸水和近岸种为主。夏季优势种有7种,秋季有9种,其中并基角毛藻、蛇目圆筛藻、琼氏圆筛藻、虹彩圆筛藻、尖刺伪菱形藻和骨条藻在两季均为优势种。
(2)夏季浮游植物平均丰度(1 713.58×104个/m3)高于秋季(1 430.30×104个/m3);该海域浮游植物群落稳定性较好,且秋季较夏季稳定性更好。
(3)温度、盐度、悬浮物、溶解无机磷及溶解无机氮等环境因子对夏、秋两季浮游植物群落影响较大,但存在季节差异。
(4)近30 a来,舟山海域富营养化程度严重,且日益加剧。同时浮游植物群落变化显著,体现为细胞丰度增加和优势种更替。
致谢感谢国家海洋局第二海洋研究所刘小涯、胡佶、蔡小霞和张海峰提供调查航次环境理化因子等数据!
参考文献(References):
[1] PENG Bo. Port locational potential evaluation and development strategy research on Zhoushan Islands new district-based on empirical analysis of Zhoushan, Ningbo and Shanghai Port locational Potential[J]. Economic Geography,2013,33(6):114-131.
彭勃.舟山群岛新区港口区位势评价及其发展战略研究——基于舟山、宁波、上海三港区位势的实证分析[J].经济地理,2013,33(6):114-131.
[2] LIU Xue-qin. Eutrophication assessment in coastal waters of Zhoushan[J]. Transaction of Oceanology and Limnology,2005(2):55-60.
刘雪芹.舟山近岸海域富营养化评价[J].海洋湖沼通报,2005(2):55-60.
[3] YE Shu-feng, DING De-wen, WANG Wen-hua. Large-scale estuarine engineering and estuarine habitat fragmentation of water body in the Yang River Estuary[J]. Acta Ecologica Sinica,2005,25(2):268-272.
叶属峰,丁德文,王文华.长江河口大型工程与水体生境破碎化[J].生态学报,2015,25(2):268-272.
[4] WANG Jin-hui, HUANG Xiu-qing, LIU A-cheng, et al. Tendency of the biodiversity variation nearby Changjiang Estuary[J]. Marine Science Bulletin,2004,23(1):32-39.
王金辉,黄秀清,刘阿诚,等.长江口及邻近水域的生物多样性变化趋势分析[J].海洋通报,2004,23(1):32-39.
[5] CLERN J E, DUFFORD R. Phytoplankton community ecology: principles applied in San Francisco Bay[J]. Marine Ecology Series,2005,285(1):11-28.
[6] JIANG Zhi-bing, LIU Jing-jing, LI Hong-liang, et al. Distribution and long-term changes of net-phytoplankton in the tidal freshwater estuary of Changjiang during wet season[J]. Acta Ecological Sinica,2013,18(33):5 917-5 925.
江志兵,刘晶晶,李宏亮,等.丰水期长江感潮河口段网采浮游植物的分布与长期变化[J].生态学报,2013,18(33):5 917-5 925.
[7] GUO Pei-yong, SHEN Huan-ting. Research advance in estuarine phytoplankton ecology[J]. Chinese Journal of Applied Ecology,2003,14(1):139-142.
郭沛勇,沈焕庭.河口浮游植物生态学研究进展[J].应用生态学报,2003,14(1):139-142.
[8] WEI Na, HU Jing-yan, MAO Hong-yue,et al. Survey and study of phytoplankton ecology in Zhoushan fishing ground and adjacent waters[J]. Marine Environmental Science,2010(2):170-183.
魏娜,胡颢琰,毛宏跃,等.舟山渔场及其邻近海域浮游植物生态调查与研究[J].海洋环境科学,2010(2):170-183.
[9] ZHANG Qi-long, WANG Fan, ZHAO Wei-hong,et al. Seasonal characteristics in the water masses in Zhoushan fishing ground and adjacent region[J]. Acta Oceanologica Sinica,2007,29(5):1-9.
张启龙,王凡,赵卫红,等.舟山渔场及其邻近海域水团的季节特征[J].海洋学报,2007,29(5):1-9.
[10] ZHANG Bin, ZHANG Fei-jun, GUO Yuan-ming,et al. Micronutrients distributions and their limitation on phytoplankton in the Zhoushan fishery ground in spring and summer[J]. Journal of Zhejiang Ocean University:Natural Science,2011,30(3):185-189.
张宾,章飞军,郭远明,等.舟山渔场海域春夏季氮磷分布及其对浮游植物的限制[J].浙江海洋学院学报:自然科学版,2011,31(3):185-189.
[11] WANG Qi. Study on phytoplankton community structure and seasonal variation in Ningbo-Zhoushan Port water[D]. Ningbo: Ningbo University,2014.
王歧.宁波—舟山港海域浮游植物群落结构及季节变化[D].宁波:宁波大学,2014.
[12] ZHU Gen-hai, YAMANOTO T, OHTANI S, et al. A study on nano- and microphytoplankton and causative species of red tide in adjacent waters off the Zhoushan Islands, Zhejiang[J]. Donghai Marine Science,18(1):28-35.
朱根海,山本民次,大谷修司,等.浙江舟山群岛邻近海域微、小型浮游植物与赤潮生物研究[J].东海海洋,18(1):28-35.
[13] GB/T 17378.4-2007 The specification of marine monitoring [S]. Beijing: China Standard Press,2007.
GB/T 17378.4-2007海洋监测规范[S].北京:中国标准出版社,2007.
[14] PIELOU E C. An introduction to mathematical ecology[M]. New York: Wiley-Interscience, 1969.
[15] MAGURRAN A E. Ecological diversity and its measurement[M]. New Jersey: Princeton University Press,1988:1-179.
[16] ZOU Jing-zhong, DONG Li-ping, QIN Bao-ping, et al. The preliminary discussion of eutrophication and red tide in Bohai bay[J]. Marine Environmental Science,1983,5(2):41-54.
邹景忠,董丽萍,秦保平,等.渤海湾富营养化和赤潮问题的初步探讨[J].海洋环境科学,1983,5(2):41-54.
[17] GB3097-1997 Sea water quality standard[S]. Beijing: China Standard Press,1997.
GB3097-1997中华人民共和国海水水质标准[S].北京:中国标准出版社,1997.
[18] TANG Feng, JIANG Xia-min, WANG Qi, et al. Dynamic of phytoplankton in typical sea areas of Zhoushan[J]. Marine Environmental Science,2013,32(1):67-72.
唐峰,蒋霞敏,王歧,等.舟山典型海区浮游植物的动态变化[J].海洋环境科学,2013,32(1):67-72.
[19] GAO Yu, JIANG Zhi-bing, ZHU Xu-yu, et al. Community composition of phytoplankton in Qushan Island in autumn and related affecting factors[J]. Marine Science Bulletin,2012,12(6):654-660.
高瑜,江志兵,朱旭宇,等.衢山岛海域秋季浮游植物群落组成及影响因素[J].海洋通报,2012,12(6):654-660.
[20] CHUNG S W, JAN S, LIU K K, et al. Nutrients fluxes through the Taiwan strait in spring and summer[J]. Journal of Oceanography,1999,57(1):47-53.
[21] HUANG Xiao-ping, HUANG Liang-min. Progress in research on dynamical processes of phytoplankton ecology in maximum turbidity zone of estuary[J]. Acta Ecologica Sinica,2002,22(9):1 528-1 531.
黄小平,黄良民.河口最大浑浊带浮游植物生态动力过程研究进展[J].生态学报,2002,22(9):1 528-1 531.
[22] IRIGOIEN X, CASTEL J. Light limitation and distribution of chlorophyll pigments in a highly turbid estuary: Gironde ( SW France) [J]. Estuarine Coastal and Shelf Science,1997,44(4):507-517.
[23] De JONGE V N, BENUSEKOM J E. Contribution of resuspended microphytotobenthos to total phytoplankton in the Emsestuary and its possible role for grazers [J]. Netherlands Journal of Sea Research,30(12):91-105.
[24] WU Yu-lin, FU Yue-na, ZHANG Yong-shan, et al. Phytoplankton distribution and its relation to the runoff in the Changjiang (Yangzte) estuary[J]. Oceanologia et Limnologia Sinica,2004,35(3):246-251.
吴玉霖,付月娜,张永山,等.长江口海域浮游植物分布及其与径流的关系[J].海洋与湖沼,2004,35(3):246-251.
[25] TANG Feng-hua, WU Yu-mei, FAN Wei, et al. Preliminary discussion on phytoplankton distribution and its relation to the runoff in the Yangtze River estuary[J]. Ecology and Environment,2010,19(12):2 934-2 940.
唐峰华,伍玉梅,樊伟,等.长江口浮游植物分布情况及与径流关系的初步探讨[J].生态环境学报,2010,19(12):2 934-2 940.
[26] CHEN Bi-juan, CHEN Ju-fa, CUI Yi, et al. The ecological characteristics of phytoplankton in the eastern marine culture area of Laizhou Bay[J]. Marine Fisheries Research,2001,22(3):64-70.
陈碧娟,陈聚法,崔益,等.莱州湾东部养殖区浮游植物的生态特征[J].海洋水产学研究,2001,22(3):64-70.
[27] The State Environmental Protection Administration. Aquatic biological monitoring[M]. Nanjing: Southeast University Press,1993.
国家环保局水生生物监测手册编委会.水生生物监测手册[M].南京:东南大学出版社,1993.
[28] MA Jian-xin, ZHEN Zhen-hu, LI Yun-ping, et al. The distribution characteristics of phytoplankton in Laizhou bay[J]. Transactions of Oceanology and Limnology,2002(4):63-67.
马建新,郑振虎,李云平,等.莱州湾浮游植物分布特征[J].海洋湖沼通报,2002(4):63-67.
[29] CAI Li-zhe, MA Li, GAO Yang, et al. Analysis on assessing criterion for polluted situation using species diversity index of marine macrofauna[J]. Journal of Xiamen University:Natural Science,2002,41(5):641-646.
蔡立哲,马丽,高阳,等.海洋底栖动物多样性指数污染程度评价标准的分析[J].厦门大学学报:自然科学版,2002,41(5):641-646.
[30] PINELENY J L, PAERL H W, HARRINGTON M B, et al. Annual eyeles of phytoplankton community-structure and bloom dynamics in the Neuse River Estuary, North Carolina[J]. Marine Biology,1998,131(2):371-381.
[31] LI Tie-jun, ZHANG Yu-rong, DING Yue-ping, et al. Analysis of phytoplankton Near Zhoushan Luomen Island[J]. Journal of Zhejiang Ocean University: Natural Science,2013,4(32):304-308.
李铁军,张玉荣,丁跃平,等.舟山螺门海域浮游植物现状分析[J].浙江海洋学院学报:自然科学版,2013,4(32):304-308.
Distribution and diversity of net-collected phytoplankton in the waters nearby Zhoushan Islands
CHEN Yue1,2, LIU Jing-jing1,2, JIANG Zhi-bing1,2, HUANG Wei1,2, SHOU Lu*1,2, ZENG Jiang-ning1,2
(1.LaboratoryofMarineEcosystemandBiogeochemistry,SOA,Hangzhou310012,China; 2.TheSecondInstituteofOceanography,SOA,Hangzhou310012,China)
Abstract:The net-collected phytoplankton community and physicochemical properties of the waters nearby Zhoushan Islands during summer and autumn of 2014 were investigated. A total of 6 phyla, including 92 genera and 184 species were indentified, of which 127 diatom and 30 dinoflagellat species were found in this survey area. The species Skeletonema spp. is dominant in summer, while the dominant species in autumn consists of nine diatom species (i.e., Coscinodiscus jonesianus, Coscinodiscus oculus-iridis, Coscinodiscus argus, Coscinodiscus asteromphalus, Skeletonema spp., Pseudo-nitzschia pungens, Chaetoceros decipiens, Thalassiothrix frauenfeldii and Frustulia interposita). The phytoplankton abundance in summer (1 713.58×104 ind/m3) is higher than that in autumn (1 430.30×104 ind/m3). According to the results of Redundancy Analysis, the temperature, salinity, suspended particulate matter, dissolved inorganic phosphorus and nitrogen are the main environmental factors affected on the phytoplankton community in summer and autumn. Based on the analysis of the integrated historical data, the eutrophication index at the research area has increased clearly since 1996, and the structure of phytoplankton community has also changed significantly. It reflects on the cells abundance increase and the dominant species substitution.
Key words:Zhoushan Islands; phytoplankton; community composition; environmental factor
Doi:10.3969/j.issn.1001-909X.2016.01.010
中图分类号:Q948.8
文献标识码:A
文章编号:1001-909X(2016)01-0076-08
作者简介:陈悦(1989—),女,浙江杭州市人,研究实习员,主要从事海洋生态学方面的研究。E-mail: chanyue0803@sina.com*通讯作者:寿鹿(1979—),男,副研究员,主要从事海洋生物实验生态学方面的研究。E-mail: shoulu981@sina.com
基金项目:国家海洋公益性行业科研专项项目资助(201305009,201405007);国家海洋局第二海洋研究所基本科研业务费专项项目资助(JG1311,JG1412);浙江省自然科学基金项目资助 (LY13D060004,Y14D060031)
收稿日期:2015-08-04修回日期:2015-11-17
陈悦,刘晶晶,江志兵,等. 舟山群岛海域网采浮游植物分布与多样性[J]. 海洋学研究, 2016,34(1):76-83, doi:10.3969/j.issn.1001-909X.2016.01.010.
CHEN Yue, LIU Jing-jing, JIANG Zhi-bing,et al. Distribution and diversity of net-collected phytoplankton in the waters nearby Zhoushan Islands[J]. Journal of Marine Sciences, 2016,34(1):76-83, doi:10.3969/j.issn.1001-909X.2016.01.010.
