吕泗渔场沿岸海域的虾类数量分布及其与环境的关系*
2016-04-20陈佳杰徐兆礼
徐 捷, 陈佳杰, 徐兆礼
(中国水产科学研究院东海水产研究所, 上海 200090)
吕泗渔场沿岸海域的虾类数量分布及其与环境的关系*
徐捷, 陈佳杰, 徐兆礼**
(中国水产科学研究院东海水产研究所, 上海 200090)
摘要:本文根据吕泗渔场及附近海域2010年春季(5月)和夏季(9月)的渔业资源调查资料,研究吕泗渔场沿岸海域虾类资源密度的时空分布,分析优势种、水团、水深和水文等环境因素对虾类数量分布的影响。研究表明:5和9月,虾类重量密度为24.21和2.19 kg·km(-2),尾数密度为12.68×103和1.30×103ind·km(-2)。2个季节虾类资源密度的平面分布特征呈相同的规律,都是外部水域高于沿岸水域。5月,虾类主要优势种由葛氏长臂虾(Palaemon gravieri)、鲜明鼓虾(Alpheus distinguendus)和日本鼓虾(Alpheus juponicus)构成,其中葛氏长臂虾对尾数和重量的贡献率较大,为5月最重要的优势种;9月,虾类优势种由哈氏仿对虾(Parapenaeopsis hardwickii)、中华管鞭虾(Solenocera crassicornis)和葛氏长臂虾(Palaemon gravieri)构成,其中哈氏仿对虾对尾数贡献率最大,对重量贡献率较大,为9月最重要的优势种。运用回归分析方法,结合虾类资源的密度、温度、盐度及水深等环境因子,对调查海域虾类资源时空分布特征进行了研究。结果表明:吕泗渔场沿岸海域虾类数量的变化主要来自优势种的贡献;虾类资源密度季节变化特征与调查海域底温呈显著负相关;特殊的沙棘潮沟地形与虾类密度平面分布关系明显,较深的潮沟往往是虾类资源集中分布的区域。本研究对于认识吕泗渔场沿海虾类资源动态变化有一定的科学意义,同时也为掌握该海域渔场渔汛及虾类资源的合理开发和保护提供依据。
关键词:吕泗渔场; 虾类; 优势种; 资源密度; 时空分布; 环境因子
XU Jie, CHEN Jia-Jie, XU Zhao-Li. Relationship of shrimp distribution with environment in the coastal water of Lvsi fishing ground in spring and summer [J]. Periodical of Ocean University of China, 2016, 46(3): 41-49.
吕泗渔场位于江苏省沿岸以东海域,为中国著名渔场。渔场近海苏北浅滩地处黄海南部,受苏北沿岸流、黄海暖流及长江径流等水团影响,因此该水域饵料生物繁生[1],不仅是鱼类,也是虾类索饵、产卵、繁育的良好场所。
中国早期对近海虾类的研究主要集中于分类方面,比如蔡秉及[2]、刘瑞玉等[3-4]研究了黄东海经济虾类的分类和区系特点,董聿茂等[5-7]对浙江沿海游泳虾类的分类进行了研究。近20年来,有关虾类种类组成、资源分布及群落结构的研究报道大量出现。比如徐兆礼等[8-10]研究了东海磷虾和糠虾类数量分布和生态适应,赵蒙蒙等[11-12]研究了海州湾南部海域虾类数量及群落特征,卢占晖等对东海大陆架虾类资源量进行了评估[13]。吕泗渔场近海为辐射沙脊浅滩地形海域,国内就浅滩水域虾类的报道不多,国外对于这类地形海域虾类的研究比较活跃,比如Gammelsrod[13]研究了莫桑比克索法拉浅滩沿海虾类丰度的变化与赞比西河径流的关系,Sabri等[14]研究了黑海南部斯诺半岛沿岸褐虾和长臂虾的时空变化,Charles R. Bronte等[15]研究了苏必利尔浅滩水深分布对虾类资源产卵场的影响。
本文利用2010年在江苏省如东县吕泗港附近海域开展的渔业资源调查资料,对吕泗渔场沿岸海域虾类资源密度和优势种的时空分布进行了研究,讨论吕泗渔场沿岸海域虾类资源分布与环境的关系,以期为该海域虾类资源的合理利用提供科学依据,对于认识中国沿海虾类资源动态变化有一定的科学意义。
1材料与方法
1.1 调查地点和采样方法
2010年在江苏省如东吕泗港附近海域(32°00′N~32°50′N、121°40′E~122°00′E)分别进行春季(5月8—12日)和夏季(9月4—10日)2个航次的渔业资源调查,2次调查分别设12个拖网站,站位设置见图1。调查船为苏如渔04416#,船体长度27.6m,宽度5.4m,主机功率198.53kW,采用单拖网(15m(宽)×5m(高)),网目范围2~5cm,拖网长度35m,网板规格1.1m长×0.7m宽,每网拖曳约0.5 h,平均拖速3.4节。
调查按《海洋水产资源调查手册》[16]对渔获物中虾类进行分物种的重量和尾数统计,记录网产量,并对每个物种进行生物学测定,记录体长、体重、成幼体比等数据。
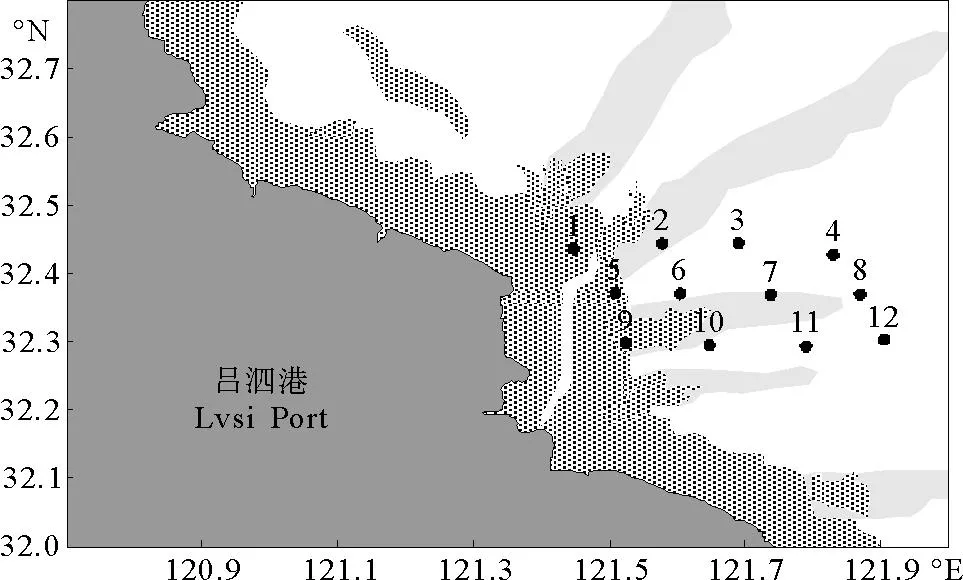
图1 调查站位分布
1.2 计算方法
虾类资源密度的估算可以采用扫海面积法[17]。在拖网统计结果基础上,计算各站位重量密度和尾数密度,公式如下:
ρi=Ci/aiq。
(1)
式中:ρi为第i站的虾类资源密度(重量:kg·km-2;尾数:103ind·km-2);Ci为第i站的每小时拖网渔获物中虾类数量(重量:kg·h-1;尾数:ind·h-1);ai为第i站的网具每小时扫海面积(km2·h-1)(网口水平扩张宽度(单位为km,本网具为0.0075km)×拖曳距离(km)),拖曳距离为拖网速度(km·h-1)和实际拖网时间(h)的乘积;q为网具捕获率(可捕系数,q=1-逃逸率),依据拖网网具,重量逃逸率取0.5,尾数逃逸率取0.5[18]。
Pinkas[19]的相对重要性指数IRI被采用来研究虾类优势种的优势度,计算公式如下:
IRI= (N% +W% )×F%。
(2)
式中:N%为某一种类的尾数占总尾数的百分比;W%为某一种类的重量占总重量的百分比;F%为某一种类出现的站数占调查总站数的百分比。
对虾类数量变化的动力学分析采用方差贡献法[20]。分析从两个方面进行,一是分析优势种对虾类数量变化的影响,在计算中以虾类总重量或尾数密度为因变量,各优势种重量或尾数密度为自变量;二是分析环境因子对虾类数量及主要优势种变化的影响,在计算中用同步观测到表层、底层水温(℃)和表层、底层盐度4个因子作自变量,以虾类及优势种的重量或尾数密度为因变量。上述分析具体的计算方法还可以参考Christensen[21]。
2结果
2.1 虾类资源密度季节变化
本调查水域2010年5和9月虾类重量密度均值分别为24.21和2.19kg·km-2,尾数密度均值分别为12.68×103和1.30×103ind/km-2(见图2)。
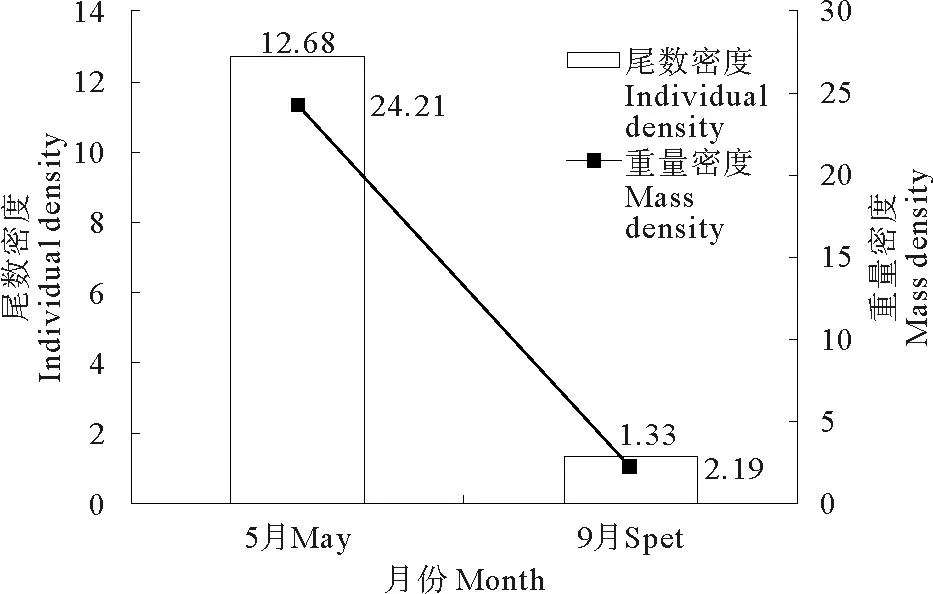
图2 虾类资源季节变化
2.2 虾类资源密度平面分布
2010年5月,虾类重量密度较高区域出现在调查水域东部的4号、7号和11号站,即32.30°N~45°N,121.70°E~90°E范围水域(见图3)。其中11号站虾类重量密度最高,为61.23kg·km-2,主要种为葛氏长臂虾(Palaemongravieri),该站位表层水温24.1℃,水深14m。4号站和7号站重量密度较高,水深分别15和10m,主要种类由葛氏长臂虾、鲜明鼓虾(Alpheusdistinguendus)和日本鼓虾(Alpheusjuponicus)构成。调查水域沿岸一侧的1号站和3号站虾类重量密度较低,其中1号站最低,重量密度仅为1.53kg·km-2。
9月,虾类重量密度最高水域位于调查水域东部的8号站,重量密度为9.13kg·km-2,表层水温15.0℃,水深18.0m,主要种为哈氏仿对虾(Parapenaeopsishardwickii)。调查海域东南部10号和12号站的重量密度相对较高(2.61和3.88kg·km-2),水深分别为13.5和19.0m,主要由哈氏仿对虾(Parapenaeopsishardwickii)和中华管鞭虾(Solenoceracrassicornis)构成。除此以外,其他站位分布较为平均,重量密度都不高。西北部5号站位鱼类重量密度仅为0.15kg·km-2,为所有调查站位中重量密度最少(见图2)。
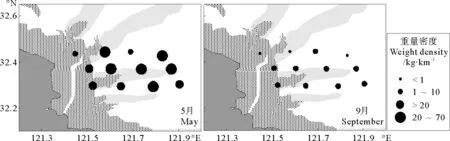
图3 吕泗渔场沿岸海域虾类重量密度(kg/km2)分布
2010年5月,尾数密度最大站位同样出现在东部水域的11号站位,其尾数密度为24.39×103ind·km-2,主要种类为日本鼓虾和葛氏长臂虾。另外调查水域西北部4号和7号站有较高的尾数密度,前者是鲜明鼓虾和葛氏长臂虾,后者仅为葛氏长臂虾(见图4)。其中,葛氏长臂虾、日本鼓虾和鲜明鼓虾几乎每个站位都出现,且有较大的数量比例。
9月,虾类尾数密度分布与重量密度分布基本一致,呈现出东部外海水域较高的特征(见图4)。尾数密度最大站位出现在东部外海水域8号站,尾数密度为3.84×103ind·km-2,主要由哈氏仿对虾和中华管鞭虾构成。另外西南部的10号站和12号站,虾类的尾数密度也较高,分别为1.87×103和1.93×103ind·km-2,主要种也为哈氏仿对虾和中华管鞭虾,这2个种每个站位都有出现,且数量较多。

图4 吕泗渔场沿岸海域虾类尾数密度(103ind/km2)分布
2.3 主要优势种平面分布及相对重要性指数
从虾类优势种的数量特征(见表1)可见:5月葛氏长臂虾是吕泗渔场沿岸海域最重要的种类,其重量占了虾类总重量的64.25%,其尾数占了虾类总尾数的55.43%,重要性指数IRI高达10 787.05,明显高于其它物种,出现率高达91.67%,广泛分布于调查水域(见图5和6)。另外,该月份鲜明鼓虾和日本鼓虾也有较高的资源密度,且出现率较高,也为重要的优势种。
9月,哈氏仿对虾大量出现,每个调查站位都有出现,占虾类重量(66.88%)和尾数(56.47%)比例都是最高,因而成为最重要的优势种,相对重要性指数IRI指数高达12 334.18,远高于其他种类,其分布特征与虾类总重量和尾数密度基本一致(见图5和6);其次是中华管鞭虾,IRI指数为3 150.47,5月的第一优势种葛氏长臂虾虽然也为9月的优势种,但优势度大幅下降。
2.4 优势种密度对虾类总密度的贡献
虾类优势种密度对总密度的贡献如表2所示。从表中可以发现,5月,除了细螯虾(Leptochelagracilis),其它优势种葛氏长臂虾、鲜明鼓虾、日本鼓虾和疣背宽额虾对虾类密度变化都有密切的贡献,其中葛氏长臂虾贡献率最大,重量贡献率β为0.77,尾数为0.54 (见表2),远高于其他优势种。其次是鲜明鼓虾和日本鼓虾,都有十分显著的重量和尾数贡献率。9月,哈氏仿对虾优势明显,无论对于重量贡献和尾数贡献均最大,分别达到0.96和0.90,远高于其他优势种。除了哈氏仿对虾,其它优势种诸如中华管鞭虾、细指长臂虾和脊尾白虾与虾类资源密度变化关系也十分显著(见表2)。
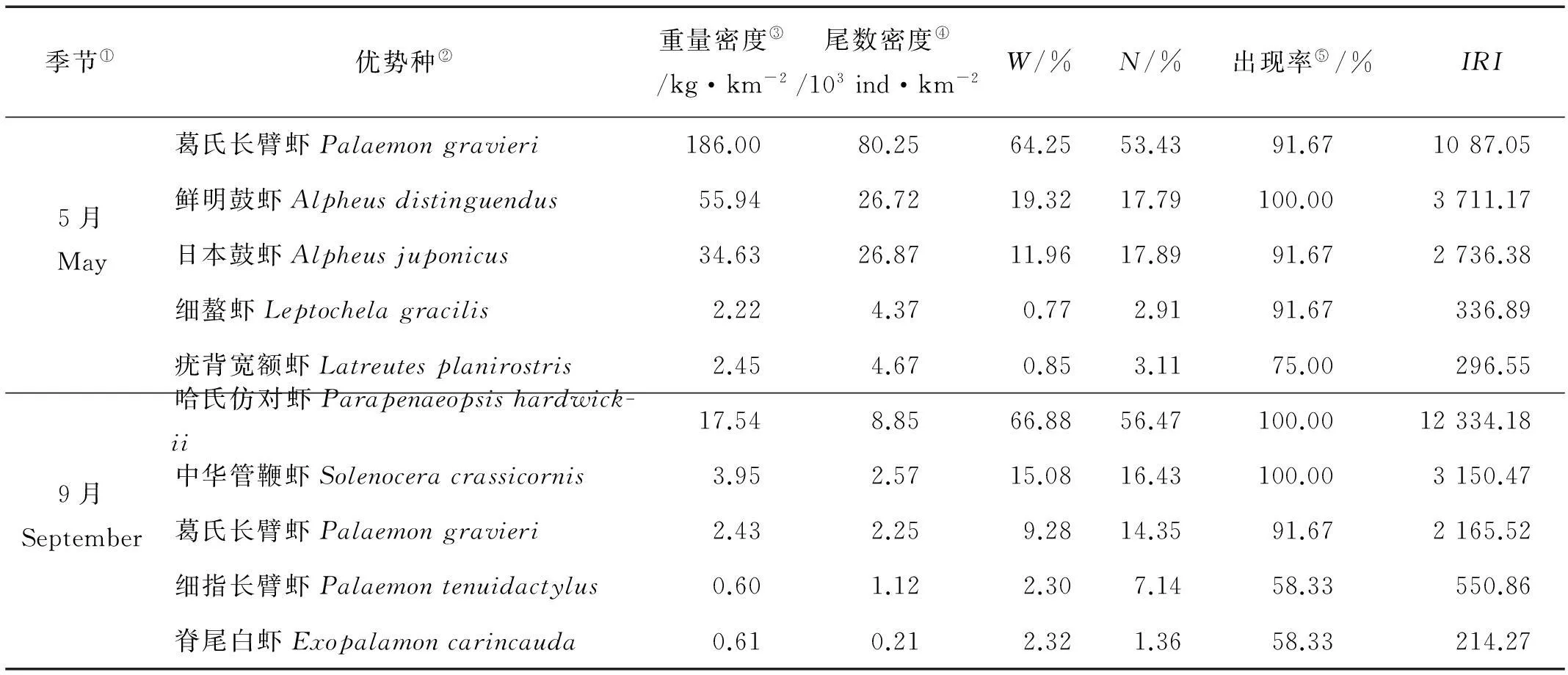
表1 虾类主要优势种生态特征
注:W为虾类重量百分比;N为虾类尾数百分比;IRI为相对重要性指数。Note:Wmeans weight percentage of dominant species in the shrimp,Nmeans individual percentage of dominant species in the shrimp;IRImeans dominance.
①Season; ②Dominant species; ③Weight density; ④Individual density; ⑤Occurrence rate
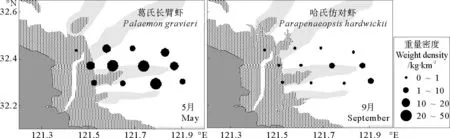
图5 吕泗渔场沿岸海域虾类主要优势种重量密度分布示意图
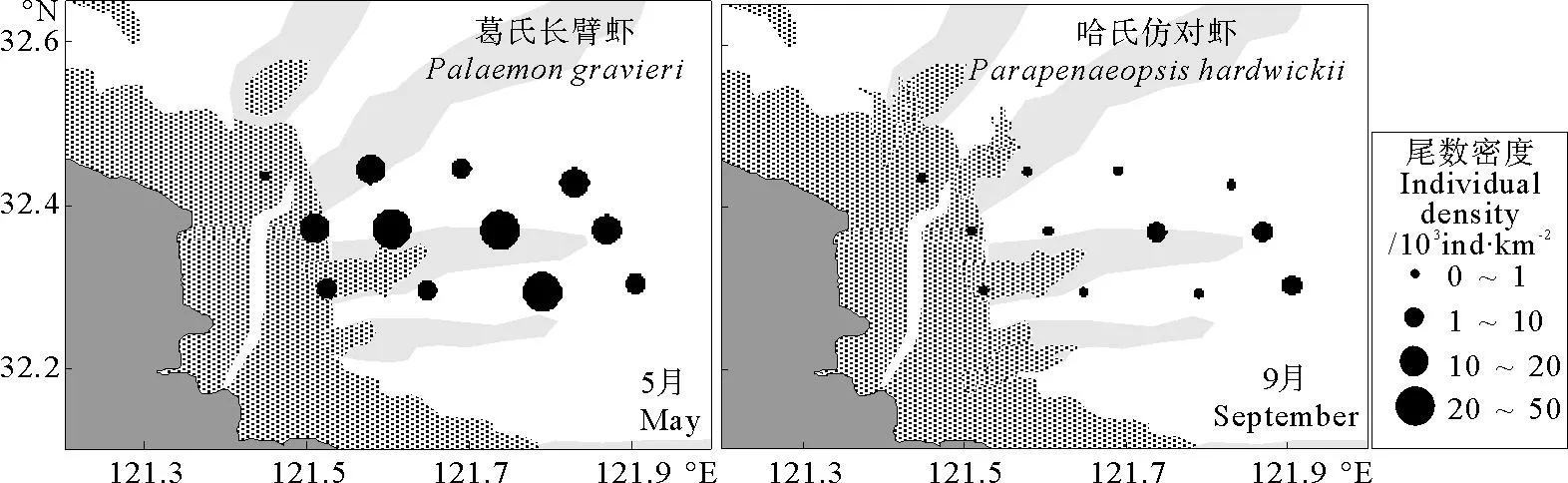
图6 吕泗渔场沿岸海域虾类主要优势种尾数密度分布示意图
2.5 温度和盐度对虾类数量分布的影响
虾类重量或尾数密度与表、底层水温(℃)和盐度4个自变量做逐步回归分析表明,底温与虾类重量密度和尾数密度有显著的负相关关系,方程分别为W=62.36-2.51t0(R=0.68,p=0.000)和N=32.11-1.29t0(R=0.78,p=0.000)。相关系数p=0.000,故回归方程自变量和因变量的相关性极显著。
表、底层水温和盐度4个自变量对2个月份虾类主要优势种葛氏长臂虾、哈氏仿对虾、鲜明鼓虾、日本鼓虾和中华管鞭虾的资源密度做逐步回归分析。结果发现:底温对葛氏长臂虾、鲜明鼓虾和日本鼓虾的重量和尾数密度均存在显著的负相关关系,葛氏长臂虾重量和尾数标准回归系数β值分别为0.66和0.84,鲜明鼓虾分别为0.57和0.55,日本鼓虾分别为0.43和0.44;底温对哈氏仿对虾和中华管鞭虾的重量和尾数密度均存在显著的正相关关系,哈氏仿对虾重量和尾数标准回归系数β值分别为0.43和0.52,中华管鞭虾分别为0.67和0.66。根据本次调查,5月调查水域平均底温为15.2℃,9月平均底温为24.0℃,底温差异影响了主要优势种资源季节变化,而虾类总体资源密度由主要优势种决定。其他环境因子对主要虾类优势种密度影响不显著。
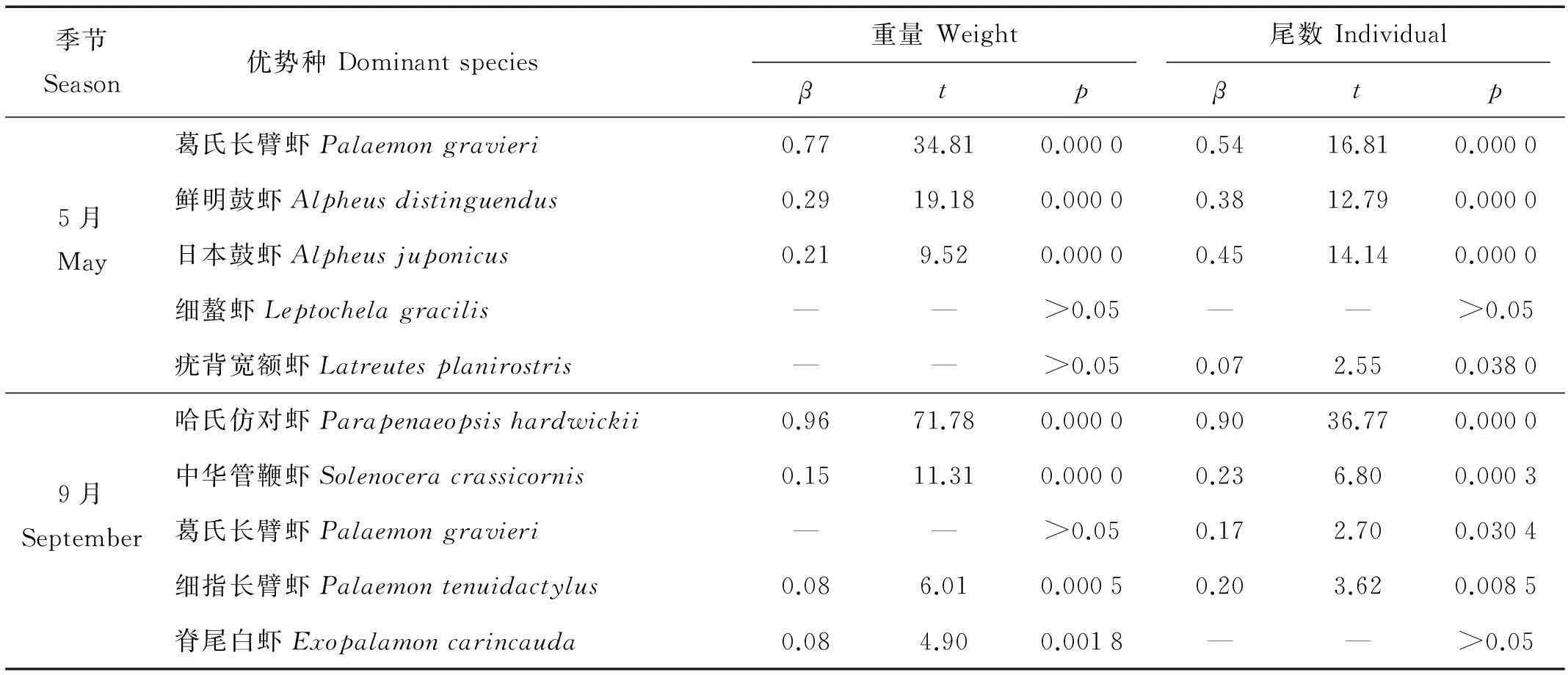
表2 优势种密度对虾类总密度的贡献
注:β:标准回归系数;t:t检验值;p:显著性;“—”代表无显著性。Note:β:Standard regression coefficient;t:tvalue;p:Significant;“—”Means no significance.
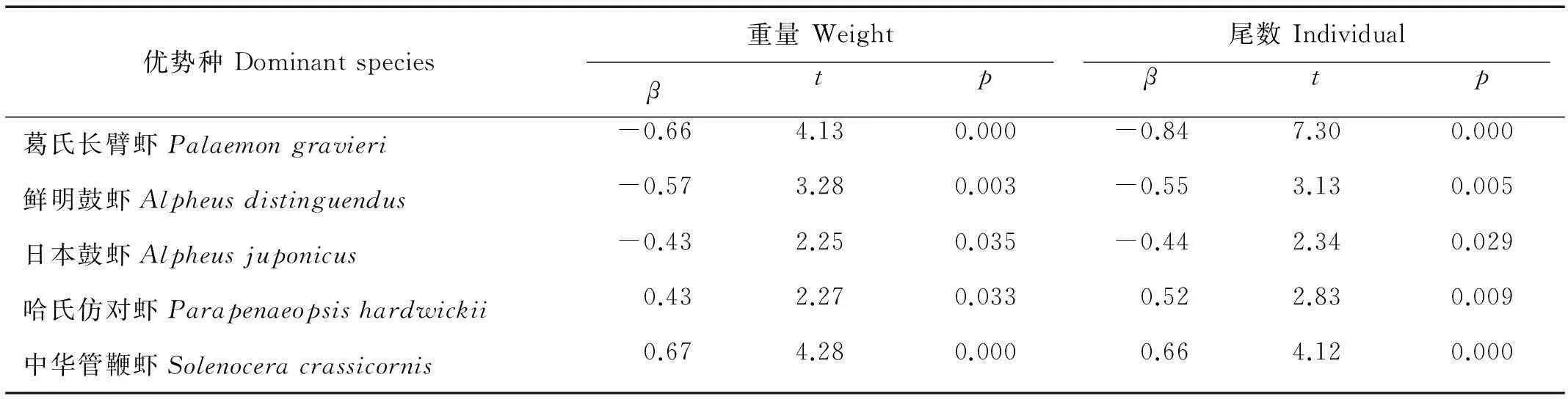
表3 底温对虾类主要优势种资源密度影响
注:β:标准回归系数;t:t检验值;p: 显著性。Note:β:Standard regression coefficient;t:tvalue;p:Significant.
2.6 水深对虾类数量的影响
1.1一般资料本组收治的宫颈癌患者40例,对40例患者随机分为两组每组各20例,其中对照组20例,年龄30~65岁,平均年龄42±2.5岁;按FIG0国际分期标准:Ib2期12例,IIa期8例。肿瘤形态:菜花型6例,结节型6例,溃疡型8例。病理诊断:鳞癌I级5例,II级9例,III级6例。治疗组20例,年龄32~64岁,平均年龄43±3.5岁;按FIG0国际分期标准:Ib2期13例,IIa期7例。肿瘤形态:菜花型5例,结节型7例,溃疡型8例。病理诊断:鳞癌I级6例,II级9例,III级5例。两组患者一般资料形态等无显著差异,具有统计学意义(P>0.05)。
回归分析表明,水深与重量和尾数密度相关性明显。5月水深与虾类的重量(p=0.056)和尾数(p=0.042)较为显著。9月水深与虾类重量(p=0.050)和尾数(p=0.026)相关性非常显著。
从表4可以进一步发现5和9月虾类资源密度随水深变化相同。春季,不同水深水域均为重量密度高于尾数密度;秋季,虾类资源密度大幅下降,无论是10m以内还是10~20m水域虾类重量密度均高于尾数密度。2个月份呈现另一相同规律是随着水深加深,虾类重量密度和尾数密度都会随之上升,资源密度集中分布于水深为10~20m水域,其中尤以5月较为明显。
3讨论
3.1 吕泗渔场沿岸海域虾类季节变化特征分析
在本调查中,2010年吕泗渔场沿岸海域不同季节的虾类尾数密度和重量密度变化趋势一致,5月虾类重量密度和尾数密度均高于9月,5月虾类尾数密度是9月的1.7倍,重量密度是9月的1.9倍,可以看出5月虾类资源量远大于9月。
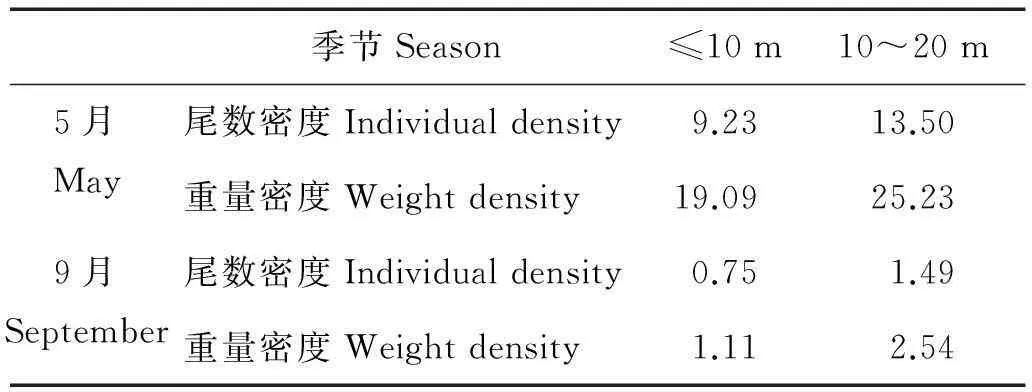
表4 不同水深虾类数量的变化
研究发现,春季是吕泗渔场虾类的渔期旺季[23-24],因此春季虾类群体数量高于夏季。根据本次调查,5月水温范围在14~18℃之间,适合虾类栖息生长。而这个季节虾类多为产卵群体,所以春季随着沿岸水温回升,虾类群体从外侧深水海域进入沿岸浅水海区产卵,夏秋季随着水温升高,虾类生活环境发生改变。所以,与5月相比,沿岸调查海域虾类无论在重量密度还是尾数密度都大幅下降(见图2和3)。
逐步回归分析结果表明,吕泗渔场沿岸海域虾类重量密度和尾数密度与底温关系非常显著。依据计算得到的回归方程,重量密度和尾数密度均与底层温度呈负相关。进一步证实了水温较低的春季更加适合虾类的聚集,而夏季虾类数量由于水温升高导致虾类资源减少。
3.2 吕泗渔场沿岸海域虾类数量空间分布变化特征分析
根据沿岸海域地形特点(见图1),吕泗渔场沿岸海域地貌为辐射状沙脊群,海底地形主要由潮沟和沙脊相间分布构成,虾类数量分布格局与沙脊潮沟地形特征密切相关。沿岸海域主要由沙脊构成,受地形阻挡的影响,虾类往往难以翻越到达沙脊水域,致使沿岸沙脊水域里的虾类密度总体上明显小于沙脊外侧潮沟处的虾类密度,造成虾类数量由外向内呈现逐渐降低的特征。从空间分布特征来看,沙脊外侧水域是虾类主要的肥育场,从资源保护出发,应加强外侧水域虾类资源保护。
3.3 优势种对虾类总数量变化的贡献
依据相对重要性指数(IRI)的计算结果(见表1),吕泗渔场沿岸海域5月虾类第一优势种为葛氏长臂虾,其对虾类总重量和尾数密度贡献均最大(见表2)。根据研究表明,葛氏长臂虾产卵高峰在春夏季,且产卵期很长,在4、5月达到顶峰[25]。所以每年5月,随着沿岸水温的回升,葛氏长臂虾大量聚集于吕泗渔场沿岸海域进行产卵,数量群体较大,且都为成体,所以本次调查葛氏长臂虾占该月份虾类总尾数和总重量百分比均为最高。春季调查还发现在调查水域外侧葛氏长臂虾资源密度较高,其分布特点与虾类总资源密度平面分布趋势基本相同(见图3~6),说明葛氏长臂虾对虾类密度平面分布有着极其重要的影响,虾类资源的总体变动情况往往由这些绝对优势种决定。除了葛氏长臂虾,其他优势种诸如鲜明鼓虾和日本鼓虾该季节也到沿岸进行产卵洄游,大量虾类群体聚集于此构成春汛虾群产卵旺季。
9月,哈氏仿对虾为第一优势种,IRI值远远高于其他优势种。研究发现,哈氏仿对虾繁殖期一般在5—9月,渔获捕捞高峰期出现在秋季[25],本次调查9月恰好是哈氏仿对虾的高峰期,所以哈氏仿对虾占9月虾类资源密度极高比例,其重量和尾数贡献率也最高(见表2)。除了哈氏仿对虾,该月份另两大主要优势种为中华管鞭虾和葛氏长臂虾。其中,中华管鞭虾繁殖期在6—8月,高峰期8—10月[26],所以秋季也是中华管鞭虾捕捞旺季。葛氏长臂虾为5月第一优势种,虽然9月也是主要优势种,但优势性大幅下降。
从上述的分析中可以发现,吕泗渔场沿岸海域虾类数量的变化动力主要来自这些经济价值极高优势种的回归贡献。所以,研究优势种环境适应特性能把握渔场资源汛期,对于资源的开发以及保护是很有必要的。
3.4 虾类优势种环境适应特征
吕泗渔场沿岸海域虾类资源密度季节变化十分明显,依据回归分析,虾类重量密度和尾数密度与底温关系非常显著,且均与底层水温呈负相关。说明水温改变是导致虾类资源发生变动重要因素。根据3.3分析,虾类数量的变化动力主要来自优势种的贡献。因为优势种是数量占比最多的关键种,因此2个月份虾类资源密度变化特征,在某种程度上可由优势种来决定。由此可见,吕泗渔场沿岸海域虾类重量密度和尾数密度与底温呈负相关的变化和可能与虾类优势种的适温性有关。
葛氏长臂虾为2个月仅有的共同优势种(见表1),5月为第一优势种,9月重量和尾数密度明显降低,IRI指数明显低于5月。这一点从优势种对总密度贡献的回归方差分析可以得到证实,春季葛氏长臂虾对总尾数和总重量的贡献分别为0.77和0.54,均为最大,而秋季仅尾数贡献率为0.17,重量上都没有贡献(见表2)。通过环境因子与葛氏长臂虾资源密度回归分析可以发现,底温对葛氏长臂虾重量和尾数密度均存在显著的负相关关系(见表3),说明调查水域底温变动是导致葛氏长臂虾资源骤减的主要原因,同时也进一步导致虾类整体资源发生变化。春季调查海域水温明显低于夏季,使得暖温性种葛氏长臂虾大量出现,推测是由于葛氏长臂虾大量聚集于吕泗渔场沿岸海域进行产卵所至。根据其他海域虾类资源的研究来看,水温相对较低的春季,整个东黄海沿岸海区葛氏长臂虾都有较高的数量群体出现,从北部海州湾、莱州湾[11-12,27],到江苏大丰、东台海域[23-24],甚至东海南部椒江口海域葛氏长臂虾都是主要优势种[26]。夏季,随着水温的进一步升高,葛氏长臂虾生活环境发生改变,虾群开始向外侧深水海域迁移,这与丁天明等[27]对葛氏长臂虾生物学特性的研究结果一致。所以,到了9月葛氏长臂虾数量大幅下降,优势性显著降低。5月虾类其他主要优势性暖温种诸如鲜明鼓虾和日本鼓虾的数量也因水温上升有所下降,回归分析结果显示均与底温有显著的负相关关系(见表3)。
夏季,哈氏仿对虾大量出现,优势性最高(见表2)。根据回归分析结果来看,发现底温对哈氏仿对虾的重量密度和尾数密度均存在显著的正相关关系(见表3)。哈氏仿对虾属广温、广盐性的暖水种,对温度的适应范围较高。研究发现,夏秋季高温季节,随着台湾暖流向北推进,哈氏仿对虾密集中心向北移动,主要分布在东海北部、黄海以南海域。冬春季随着水温下降,暖流向南退缩,哈氏仿对虾密集中心向东南部外侧海区移动[28]。这就解释了为什么春季调查哈氏仿对虾数量较少。到了夏季,水温开始升高,春季暖温性优势种资源大幅衰退,哈氏仿对虾的优势性逐渐显现出来。另外,该季节另一大优势种中华管鞭虾受底温影响也很明显,回归分析结果显示底温对中华管鞭虾的重量和尾数密度存在显著的正相关关系(见表3)。总之,随着夏季来临,水温升高,适应较低水温的暖温种数量减少,适应较高水温的暖水种,比如哈氏仿对虾,在数量不能补偿暖温种数量减少的部分,这是夏季虾类数量低于春季的主要原因。
所以,春夏季节交替,吕泗渔场沿岸海域虾类资源与调查水域底温变化密切相关,从上述分析可以发现虾类群落数量变动是由优势种的温度适应性决定。掌握虾类优势种环境适宜特性可以更好地开发保护资源。
3.5 虾类资源量与水深的关系
根据回归分析表明,虾类重量和尾数密度与水深相关关系较为显著。由于吕泗渔场沿岸海域地貌为辐射状沙脊群,海底地形主要由潮沟和沙脊相间分布构成,沙脊宽度由外海向中心逐渐变宽,潮沟则由外海向中心逐渐变狭变浅,呈辐射状向外伸。因此,海底地面并不平坦,地势较高的为沙脊,地势较低的是潮沟,导致调查海域不同站位水深差异十分明显。
5月,水深较大的区域往往是虾类分布的密集区。尾数密度较高的水域集中分布于10m水深以上的水域(见图5),所以春季10 ~ 20m水深海域尾数密度相对较高(见表3),其中4、7、8、11号站等水深较高站位,资源密度最高,与葛氏长臂虾所处的水深一致,且站位特点都位于调查水域外侧潮沟处,说明虾类由于索饵位置往往处于浅滩外侧水深且宽广的水域。而重量密度的分布规律与尾数密度一样,也是水深较高处虾类重量密度较高。这就直接导致10~20m水域鱼类的尾数密度和重量密度均高于10m内鱼类资源密度(见表3),说明虾群趋向于栖息于较深的水域。9月,虾类资源大幅下降,各层次水深虾类资源密度差距不大,但优势度前几位的哈氏仿对虾、中华管鞭虾主要集中于10~20m水深区域,其中,资源密度最高的8号和12号站,水深都超过10m,且都位于沙脊以东。所以总体上来看,还是所处10m以上水深虾类资源略高。和春季一样,显示出虾群还是喜欢栖息于外海较深的水环境,因此,可以认为调查海域的外侧,吕泗渔场中心位置是虾类主要渔场所在。
致谢:周进、叶金清等同志参加海上样品采集和标本鉴定,沈晓民先生在论文写作中给予很大的帮助,谨致谢忱。
参考文献:
[1]张海景, 徐兆礼. 小黄鱼育幼期吕泗渔场的饵料浮游动物特征 [J]. 生态学杂志, 2010, 29(10): 2072-2076.
ZHANG Hai-Jing , XU Zhao-Li. Distribution pattern of diet zooplanktons in Lǜsi fishing ground during nursing period of small yellow croaker [J]. Chinese Journal of Ecology, 2010, 29(10): 2072-2076.
[2]蔡秉及. 南黄海和东海磷虾分类的初步研究 [J]. 海洋通报, 1982, 20(4): 68-78.
Cai Bing-Ji. The preliminary study on the classification of krill in the south Yellow Sea and East China Sea[J]. Marine science bulltetin, 1982, 20(4): 68-78.
[3]刘瑞玉. 黄海及东海经济虾类区系的特点 [J]. 海洋与湖沼, 1959, 2(1): 35-42.
LIU Rui-Yu. Notes on the economic Macrurous crustacean fauna of the Yellow Sea and the East China sea [J]. Oceanologia et Limnologia Sinica, 1959, 2(1): 35-42.
[4]刘瑞玉. 黄、东海虾类动物地理学研究 [J]. 海洋与湖沼, 1963, 5(3): 230-244.
LIU Rui-Yu. Zoogeographical studies on the macrurous crustacean fauna of the Yellow Sea and the East China Sea [J]. Oceanologia et Limnologia Sinica, 1959, 2(1): 35-42.
[5]董聿茂, 胡萸英, 汪宝永. 浙江沿海游泳虾类报告Ⅲ [J]. 动物学杂志, 1986, 21(5): 4-6.
Dong Y M, Hu Y Y, Wang B Y. A report on shrimps of Zhejiang coast Ⅲ [J]. Chinese Journal of Zoology, 1986, 21(5): 4-6.
[6]董聿茂, 胡萸英. 浙江沿海游泳虾类报告Ⅱ [J]. 动物学杂志, 1980, 15(2): 20-40.
Dong Y M, Hu Y Y. A report on shrimps of Zhejiang coastⅡ [J]. Chinese Journal of Zoology, 1980, 15(2): 20-24.
[7]董聿茂, 虞研原, 胡萸英. 浙江沿海游泳虾类报告I [J]. 动物学杂志, 1959, 3(9): 389-394.
Dong Y M, Yu Y Y, Hu Y Y. A report on shrimps of Zhejiang coast Ⅰ [J]. Chinese Journal of Zoology, 1959, 3(9): 389-394.
[8]徐兆礼, 陈亚瞿. 东海磷虾类优势种的生态适应 [J]. 生态学报, 2005, 25(9): 2227-2233.
XU Zhao-Li , CHEN Ya-Qu . The dominant species of euphausi ids in the east China sea and their ecological adaptability [J]. Acta Ecologica Sinica, 2005, 25(9): 2227-2233.
[9]徐兆礼, 晁敏, 崔学森. 东海糠虾类数量分布 [J]. 水产学报, 2006, 30(3): 341-346.
XU Zhao-li , CHAO Min , CUI Xue-sen. The abundance distribution of mysids (Mysidacea)in the East China Sea [J]. Journal of Fisheries of China, 2006, 30(3): 341-346.
[10]徐兆礼. 东海浮游糠虾种类特征和多样性 [J]. 应用生态学报, 2006, 17(9): 1711-1714.
XU Zhao-Li . Species characteristics and diversity of pelagic Mysidacea in East China Sea [J]. Chinese Journal of Applied Ecology, 2006, 17(9): 1711-1714.
[11]赵蒙蒙, 徐兆礼. 海州湾南部海域不同季节虾类数量及其分布特征 [J]. 海洋通报, 2012, 31(1): 38-44.
Zhao Meng-Meng, Xu Zhao-Li. Abundance and distribution of shrimps in different seasons of in the southern Haizhou Bay of Jiangsu Province[J]. Marine Science Bulletin, 2012, 31(1): 38-44.
[12]赵蒙蒙, 徐兆礼. 海州湾南部海域虾类群落特征 [J]. 上海海洋大学学报, 2012, 21(6): 1038-1045.
Zhao Meng-Meng, Xu Zhao-Li. Community characteristics of shrimps in the southern Haizhou Bay [J]. Journal of Shanghai Ocean University, 2012, 21(6): 1038-1045.
[13]卢占晖, 薛利建, 张龙, 等. 东海大陆架虾类资源量评估 [J]. 水生生物学报, 2013(5): 855-862.
LU Zhan-Hui, XUE Li-Jian, ZHANG Long, et al. Assessment of shrimp stock biomass in continental shelf waters of the East China Sea [J]. Acta Hydrobiologica Sinica, 2013(5): 855-862.
[14]Gammelsrod T. Variation in shrimp abundance on the Sofala Bank, Mozambique, and its relation to the Zambezi river runoff [J]. Estuarine, Coastal and Shelf Science, 1992, 35: 91-103.
[15]Sabri B, Ozen O, Ates A S. Spatial and temporal variation ofPalaemonadspersus,PalaemonelegansandCrangoncrangon (Decapoda: Caridea) in the southern Black Sea [J]. Estuarine, Coastal and Shelf Science, 2008, 79: 671-678.
[16]Charles R. Bronte, James H Selgeby, J ames H Saylor, et al. Hatching, dispersal, and bathymetric distribution of age-0 wild lake trout at the Gull Island Shoal Complex Lake Superior [J]. Journal of Great Lakes Research, 1995, 21(1): 233-245.
[17]黄海水产研究所. 海洋水产资源调查手册(第二版) [M]. 上海: 上海科学技术出版社, 1981.
Yellow Sea Fisheries Research Institute. Marine Aquatic Resources Survey. 2nd Ed [M]. Shanghai: Shanghai Science and Technology Press, 1981.
[18]詹秉义. 渔业资源评估 [M]. 北京: 中国农业出版社, 1995: 18-46, 124-130.
ZHAN Bing-Yi. Fishery Resource Assessment [M]. Beijing: China Agriculture Press, 1995: 18-46, 124-130.
[19]杨吝, 张旭丰, 谭永光, 等. 南海区拖网网囊最小网目尺寸选择性研究 [J]. 中国水产科学, 2003, 10(4): 325-332.
YANG L, ZHANG Xu-Feng, TAN Yong-Guang, et al. Selectivity study on minimum mesh size of bottom trawl codend in South China Sea [J]. J Fish Sci China, 2003, 10 (4): 325-333.
[20]Pinkas L M, Oliphant S, Iverson I L K. Food habits of albacore, bluefin tuna and bonito in Californian waters [J]. Calif Shrimp Game, 1971, 152: 1-105.
[21]徐兆礼, 李春鞠. 东海浮游磷虾类数量分布 [J]. 水产学报, 2005, 29(3): 373-378.
XU Zhao-Li, LI Chun-Ju. Abundance distribution ofEaphausiidsin the East China Sea [J]. Journal of Fisheries of China, 2005, 29(3): 373-378.
[22]Christensen R. Analysis of variance, design and regression: applied statistical methods [M]. New York: Chapman and Hall, 1996: 75-98.
[23]阙江龙, 康伟, 徐兆礼, 等. 苏北浅滩中部海域春秋季口足目和十足目虾类分布特征 [J]. 海洋渔业, 2012, 34(3): 301-307.
QUE Jiang-Long, KANG Wei, XU Zhao-Li, et al. Distribution of Hoplocarida and Decapoda shrimps in spring and winter in the middle area of Subei Shoal[J]. Marine Fisheries, 2012, 34(3): 301-307.
[24]阙江龙, 柯昶, 徐兆礼, 等. 苏北浅滩沙脊潮沟地形和潮流对虾类分布的影响 [J]. 生态学杂志, 2013, 32(3): 661-667.
QUE Jiang-Long, KE Chang, XU Zhao-Li, et al. Effects of topography and tidal current of sand ridge and tidal creek system in North Jiangsu shoal of East China on the distribution of shrimps[J]. Chinese Journal of Ecology, 2013, 32(3) : 661-667.
[25]任中华, 郑亮, 李凡, 等. 莱州湾海域虾类群落结构及其多样性 [J]. 海洋渔业, 2014(3): 193-201.
REN Zhong-Hua, ZHENG Liang, LI Fan, et al. Community structure and diversity of shrimp in Laizhou Bay[J]. Marine Fisheries, 2014(3): 193-201.
[26]齐海明, 孙岳, 徐兆礼, 等. 椒江口海域春秋季虾类群落结构及其影响因素 [J]. 应用生态学报, 2013, 24(12): 3546-3552.
QI Hai-Ming, SUN Yue, XU Zhao-Li, et al. Shrimp community structure and its influential factors in the Jiaojiang River estuary during spring and autumn [J]. Chinese Journal of Applied Ecology, 2013, 24(12) : 3546-3552.
[27]丁天明, 宋海棠. 东海葛氏长臂虾Palaaemongravieri生物学特征研究 [J]. 浙江海洋学院学报(自然科学版), 2002, 21(1): 1-5.
Ding Tian-Ming, Song Hai-Tang. The Study on Biology ofPalaaemongravieriin East China Sea [J]. Journal of Zhejiang Ocean University(Natural Science) , 2002, 21(1): 1-5.
[28]宋海棠, 俞存根, 薛利建. 东海哈氏仿对虾的数量分布和生长特性研究 [J]. 水生生物学报, 2009, 33(1): 15-21.
Song H T, Yu C G, Xue L J. Study on the biomass distribution and growth property of Parapenaeopsis Hardwickii in the East China Sea [J]. Acta Hydrobiologica Sinica, 2009, 33(1): 15-21.
[29]凌建忠, 程家骅, 刘敏. 东海主要经济虾类繁殖期和分布区域及其资源保护的探讨 [J]. 海洋渔业, 2004, 26(1): 13-17.
Ling J Z, Cheng J H, Liu M. Discussion on the propagative periods, distribution areas and protection of the main economic shrimp in the East China Sea [J]. Marine Fisheries, 2004, 26(1): 13-17.
责任编辑朱宝象
Relationship of Shrimp Distribution with Environment in
the Coastal Water of Lvsi Fishing Ground in Spring and Summer
XU Jie, CHEN Jia-Jie, XU Zhao-Li
(East China Sea Fisheries Research Institute, Chinese Academy of Fisheries Sciences, Shanghai 200090, China)
Abstract:Lvsi fishing ground is located at the east sea area of Jiangsu Province, which is one of the most famous fishing grounds in China. The relationship between the distribution of shrimp resources and environment in the coastal waters of Lvsi fishing ground was determined in order to provide a scientific basis for the rational use of shrimp resources in this area. It has scientific significance to understand the dynamic change of shrimp resources in the Chinese coastal sea. Based on the surveying data of fish resources in the coastal area of Lvsi fishing ground in May and September, 2010, we studied the spatiotemporal distribution of shrimp density and analyzed the effect of fishery resources, such as dominant species, current flow, water depth and environmental factors, aimed to explore the attributes of the fishing ground in this sea area. In May and September, the average of shrimp weight density was 24.21 and 2.19 kg·km(-2), respectively, while the figure for individual density was 12.68×103 and 1.30×103 ind·km(-2), respectively. Both of the spatial distribution patterns were basically the same, being higher in the eastern than in the western water areas near the side of Rudong. Palaemon gravieri was the most dominant specie in May, whereas Parapenaeopsis hardwickii was the most dominant species in September. The stepwise regression analysis showed that dominant species had great contribution to the shrimp number density and weight density. Shrimp weight and individual density were significantly correlated with bottom water temperature, but less significantly with water salinity. Water depth was related to the distribution of shrimp density significantly, and the distribution of the density of regional resources was often focused on the high depth of the sea area. It was considered that sea area was an ideal feeding ground for economic fishes. Our findings will aid to finding out the change of fishery resource, and mastering the fishing season and providing reasonable basis of exploration and protection.
Key words:Lvsi fishing ground; shrimp; dominant species; resource density; spatiotemporal distribution; environmental factor
DOI:10.16441/j.cnki.hdxb. 20140427
中图法分类号:S931
文献标志码:A
文章编号:1672-5174(2016)03-041-09
作者简介:徐捷(1989-),男,研究实习员。E-mail:rihannaxu@163.com**通讯作者:E-mail:xiaomin@sh163.net
收稿日期:2014-12-23;
修订日期:2015-04-21
*基金项目:国家自然科学基金项目(41176131);海洋公益性行业科研专项经费项目(201305027-8); 浙江省科技厅项目 (2015C33246)资助
引用格式:徐捷, 陈佳杰, 徐兆礼. 吕泗渔场沿岸海域的虾类数量分布及其与环境的关系[J]. 中国海洋大学学报(自然科学版), 2016, 46(3): 41-49.
Supported by Project of National Natural Science Foundation of China (41176131);Marine Research Special Foundation for Public Welfare Projects(201305027-8); Project of Science and Technology Department of Zhejiang Province (2015C33246)
