甘南高寒草甸群落花期物候研究
2016-04-18张春花章志龙
张春花,章志龙,贾 鹏
(1.甘肃民族师范学院 甘肃 合作 747000; 2.草地农业生态系统国家重点实验室 兰州大学生命科学学院,甘肃 兰州 730020)
甘南高寒草甸群落花期物候研究
张春花1,章志龙1,贾 鹏2
(1.甘肃民族师范学院 甘肃 合作 747000; 2.草地农业生态系统国家重点实验室 兰州大学生命科学学院,甘肃 兰州 730020)
摘要:对草地群落物候的研究,不仅有助于对草地群落功能认知,更重要的是为生态系统评估提供背景资料。本研究从地理环境因素、功能性状、传粉者竞争以及系统发育4个方面讨论影响甘南高寒草甸植物花期物候的因素。结果表明,在群落水平上,开花期涵盖了整个生长季,但在整体上有一个高峰期,群落中各物种的始花期、开花峰值期及开花持续期不同,每一物种均有其独特的花期格局;植株高度(P<0.001)和种子大小(P=0.034)能够显著地预测开花,而比叶面积(SLA)则不显著(GLM,P>0.05);风媒物种开花期早于虫媒物种(P=0.008);花期物候重叠与系统发育距离之间存在着负相关关系。
关键词:高寒草甸;花期物候;功能性状;传粉;系统发育
物候学是一门古老的学科,历史上物候记录的起源很早,在中国可以追溯到公元前1000年,出于服务农业生产的需要已经按月记载全年的物候历,物候学是一门研究生物生命周期随时间变化规律的学科[1],研究在自然界中以年为周期重复发生的各种生命现象的出现时间及其与环境变化的关系。现代物候学已发展成为一门综合性学科,从形态学到物种进化及生态系统结构与功能均涉及物候学,物候是生物重要生活史性状之一[2-5]。在生态系统中,物候学是基于时间的角度对群落中不同物种的发展变化进行阐述的,是研究生态系统的一个重要学科,有助于认识群落结构及其功能在时间梯度上的变化规律。早期物候研究主要集中于单个植物种群和森林植物群落物候,而有关草地群落物候研究并不多见。在全球气候环境变化的背景下,草地群落物候的研究得到了快速的发展。青藏高原高寒草甸生态系统是陆地生态系统的重要组成部分,其重要性更加突出,其变化能够预示陆地生态系统的变化动态,对青藏高原草地群落物候的研究不仅有助于深入认识草地群落构建机制,并有助于对其功能属性的深入探究,更重要的是为青藏高原高寒草甸生态系统的预测和评估提供基础研究。基于此,本研究分析青藏高原东缘高寒草甸草本群落的花期物候,并就花期物候与环境因素,传粉模式及系统发育等的内在关联进行探讨,以期为提出更有利于植物繁殖的有效策略提供有用的参考资料。
1材料与方法
1.1试验区概况
试验点在兰州大学高寒草甸与湿地生态系统定位研究站(玛曲县阿孜分站)(33°40′ N、101 52′ E,海拔3 550 m),年均温1.2 ℃(1月均温-7.8 ℃,7月均温为11.9 ℃),无霜期大于270 d。降水主要集中于短暂的夏季,过去30年的年均降水量为620 mm。根据甘肃省56074号气象台提供的气象数据制作2008年月均温和月降水量(图1)。试验地属于典型的高寒草甸,主要物种有莎草科禾叶嵩草(Kobresiagraminifolia)、甘肃苔草(Carexkansuensis),毛茛科小花草玉梅(Anemonerivularis),禾本科垂穗披碱草(Elymusnutans)、波伐早熟禾(Poapoophagorum)、羊茅(Festucaovina)。地上部分的生产力为360~560 g·m-2。

图1 2008年试验地月均温和降雨量
1.2物候观测计算
试验地自2007年起季节性封育,每年4-11月禁牧,2008 年 4 月在季节性封育围栏内选择地势较平坦的区域设置50个0.5 m×0.5 m的固定样方,样方间隔约为3.5 m,进行物候观测。2008年从5月初到9月中旬对固定样方内出现的所有物种的物候进行观测记录,每6~7 d进行一次记录,共观测21次。将物候的观测时间转化为儒略历(Julian Calendar)即:1月1日为1年中第1天。
开花峰期通过加权日期来计算:
式中,dt是在第t次观测时的儒略日(Julianday),T是总的观测次数。nt是在t次观测时,该物种处于花期的样方数目。
花持续期通过Pianka生态位宽度指数[6]进行计算:

花期的重合通过Pianka重叠指数[6]计算:

式中,Ptj是物种j在第t次观测时开花的比例,Ptk是物种k在第t次观测时开花的比例。Ojk的大小介于0到1之间,分别表示两个物种没有重合与完全重合。
1.3功能性状的测定
选取了48种最为常见的物种进行功能性状的测定,这些物种可以控制群落85%~95%的盖度和地上生物量。在植物的花期进行采样,当某一个物种处于开花的比率超过50%时,在试验地采样区随机采集20~25株处于花期个体的地上部分进行该物种功能形状的测定。所测量的功能性状有植株自然高度、比叶面积(SLA)和种子大小。每株植物在采集后立即随机取下两片叶片(部分植物取1片)测量其叶面积,之后将其烘干并称重,样本在80 ℃下烘干48h后称重。当某物种的种子成熟后,采集种子从净种子中数取一定数量的种子称量其重量,种子的质量为百粒重,质量的称量在Sartorius电子天平 (1/10000精度)上进行。所测量的功能性状均进行了对数转化。对具有明显花结构的物种(如鲜艳的花冠),将其认定为昆虫传粉;不具有明显花结构的物种(莎草和禾草)认定为风媒传粉[7]。
1.4花期重合与系统发育
利用Mantel检验[8]来检测系统发育与花期重合的关系。试验期间共观测到70个物种,其中有55个种观测到开花。再将花期物候的数据组织成一个55行21列的矩阵:每一行代表一个物种而每一列代表一次观测。矩阵的每一个数据则表示当次观测中该物种所处于花期的样方数目。通过Phylomatic[9]以及公开数据[10]构建所观测到开花物种的系统发育关系。同科属之间以及同属物种之间的关系被假定为单源多岐(MonophyleticandPolytomous),属和物种水平的分枝长度通过Phylocom的BLADJ模块计算得出[11]:计算的方法为均匀放置属和种的结点以使得分枝长度的方差最小。两个物种的系统发育距离(PhylogeneticDistance)就是二者之间的分枝长度。计算所观测到的花期物候重叠指数与系统发育距离之间的相关系数,相关系数观测值会与999个随机化群落进行比较。对其显著性的检验水平定为P=0.05。如果观测值的秩(Rank)小于25,或者大于975,就认定观测值显著地偏离了零模型。
1.5数据分析
本研究所有的数据检验都在R2.10.1[12]下进行。
2结果与分析
2.1种间花期分布
2008年,在试验地样方中观测到了55个物种开花,对群落中物种的开花序列分布进行了研究(图2)。

图2 2008年55个物种开花序列分布图

系数CoefficientsEstimateSEtPIntercept140.07643.5973.2130.003** 传粉模式:风媒Pollination:Wind-15.4065.512-2.7950.008** 高度Height47.5067.86.09<0.001** 比叶面积SLA2.94520.1080.1460.884 种子大小Seedsize-9.3054.254-2.1880.034*
高寒草甸植物群落中各物种从5月初开始分别进入花期,至9月下旬结束,多数物种的花期处于5月末至生长季结束,群落水平的花期物候几乎遍及整个生长季。在群落水平上,开花期涵盖了整个生长季,但整体上仍然有一个高峰,群落中各物种的不同,每一物种均有其独特的花期格局。高寒草甸植物群落55种植物的始花期、开花峰值期和开花持续期见图2。
2.2开花峰值期与功能性状的一般线性模型分析
开花峰值期与功能性状:从GLM(表1),物种的传粉模式、高度、比叶面积和种子大小与开花峰期。GLM的结果表明,风媒物种开花峰期要早于虫媒物种(P=0.008);高度(P<0.001)和种子大小(P=0.034)能够显著地预测开花,而比叶面积(SLA)则不显著。从结果可以看出来,传粉模式对花期是有影响的,风媒植物的花期显著早于虫媒植物。花期物候也反映了植株高度和种子大小的权衡:晚期开花的物种往往具有较高的成体高度和较小的种子,而早期开花的物种则有较低的成体高度和较大的种子。
2.3系统发育
高寒草甸草本植物的花期物候在系统发育上比较保守,即亲缘关系越近的物种更加倾向于“同时开花”,有较高的重合度(图3)。
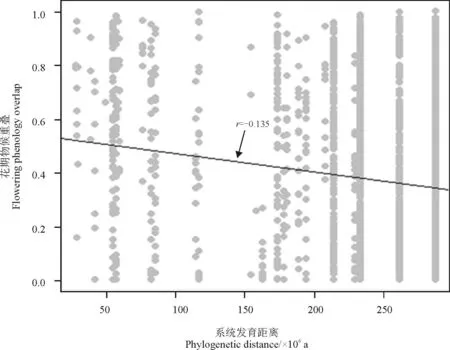
图3 花期物候重叠与系统发育距离之间的关系
注:每个点代表一个物种对。
Note: One species pair.
3讨论与结论
3.1环境因素对花期物候的影响
花期物候是植物繁殖状态的基本反映,环境因素将会对植物的繁殖产生直接的影响,环境决定不同植物间在复杂多变的生态环境中是否可以繁殖[13]。气候因素(比如温度、降水等)通常被认为是开花时间最好的判断指标[14-16]。尤其在高海拔、低温的区域,植物群落一般具备以下性状:生长季短,其花期会较早且种子的生长周期短,较易发育成熟,对环境变化敏感[17-20]。甘南高原草地受地理区域性和环境的影响大,群落中早期开花的物种一旦雪融即行开花,如报春花科的植物等就是这样的类型。有报道指出[21],一旦气温骤增,其花期提前,导致植物的生长周期提早,进而导致对资源利用分配的不平衡,影响到其它相关物种的生长节律。因此,高原草地这些特殊的地理区位下,物候会受到环境和气候要素显著的影响。
3.2花期物候与功能性状
除环境因素外,植物本身的性状对物候也具有重要的作用[22-24],而且这些性状也直接可以用来观察植物群落受环境因子的影响。植物在特定环境下的繁殖周期长短影响着植物内部的营养吸收以及生长情况,植物的生长周期内,开花期提前意味着植物在生长过程中光合作用不充分,植物内部的光合产物不充足,但是其优势在于开花期早的植物生长发育的时间增加。相反,开花期晚的植物其内部的光合产物累积充足,但是发育时间延缓,尤其是在生长季节短的高寒地区,有可能导致植物出现生长不成熟的现象。有研究已经表明,开花时间与植物个体的大小存在着正相关关系[25-27];而开花太晚的植物个体可能会无法完成结实[28-30]。根据一项对英国的115种昆虫类开展传粉活动时对其周边植物群落的影响关系研究[31]可知,植物的开花期越早,开花时间越短则植物的果实成熟度越低,果实形状越小,两者间是呈负相关的关系。在甘南高原高寒草甸草原地区,花期物候对于植物繁殖会产生较大的影响,甚至还会影响到群落的构建,所以,预测花期静候性状是关键。本研究结果也证实了上述观点:花期较早的物种高度较低,种子较大(表1)。
3.3传粉模式
植物花期物候不可避免地要受到其它生物的影响,其中传粉者的角色尤为重要[32]。一般而言,风媒植物倾向于早开花,甚至早在叶长出之前,而虫媒植物则倾向于在传粉者最活跃的时期开花[33]。开花期和传粉昆虫的关系时而互助时而竞争,由此植物界和昆虫界的相关学者对于两者间关系的研究众多。甚至出现了如下的假说:植物开花时间和传粉昆虫的竞争强度可以通过降低植物开花时间的重叠性来获得,而传粉竞争则会降低种子数量与质量。根据实证可知,共享传粉者会导致植物的繁殖成功率下降,对物种的生长起到一定的物候阻碍,导致种子产量降低[34-35]。这种情况下,选择压力会倾向于分化二者的开花时间以减少重叠[36]。然而也有研究表明,种群的开花期与传粉者多度并非同步,如有研究认为草地群落中多数植物为风媒花,因此传粉者访问对花期的影响有限[37]。本研究中风媒传粉的物种(图2),如甘肃苔草、矮藨草(Scirpuspumilus)、迷果芹(Sphallerocarpusgracilis)、羊茅、禾叶嵩草的花期要早于高山韭(Alliumsikkimense)、展毛翠雀花(Delphiniumkamaonense)、星状风毛菊(Saussureastella)、麻花艽(Gentianastraminea)、华丽龙胆(Gentianasino-ornata)这些虫媒传粉的物种。花期物候在风媒物种中同时具有很强的特异性:莎草科物种集中在群落中其它植物没有形成遮蔽之时开花,而禾本科物种,如草地早熟禾和垂穗披碱草等则在生长季的中期开花,尽管花期相对较迟,但这些禾本科植物能够保证传粉不受其它虫媒植物遮蔽的干扰。本研究的结果也证实风媒植物花期的确要早于虫媒植物(表1)。
3.4系统发育
在同一环境下,同种类别的物种生长特点和生长习性类似,所以对于传粉者或者物种间的竞争也会较为激烈,同属物种的物候特性也会相类似。在同一群落中的同属物种其变形出现差异是因为进化史中出现的改变或基因突变、遗传变化形成的,具体还是与进化史密切相关。有研究表明,物种的功能性状(包括繁殖性状)在演化上是保守的[38-39],因此,物种间的基因越相似,亲缘越近则物种的表型越类似。同一群落中,物候性状和其它的一些物种性状一样,物种的变化多是受到物候和环境的影响,群落内的物候对植物的影响关系重大,这是生态系统中所默认的进化原因。花期物候被认为是适应性的物种特性,认为会导致生态位分化,植物物候亦可被视作时间生态位[40]。普遍认为,环境筛选以及竞争格局是导致群落物种发生系统发育和进化演变的原因[41-42],竞争格局会对群落的生态系统进行分散,对于那些同属物种,其亲缘关系近,其表型则可能出现较大差别。相反,由于环境筛选而区别的同属物种则物种间的表型会类似,因此,群落物候的时间格局与其变化也可以反映环境的变化。许多研究发现,系统发育与群落内的物候有较强的相关性[43]。本研究结果表明,花期物候具有进化上的保守性,系统发育上的限制要比种间竞争作用更大。甘南高寒草甸草本植物群落的花期物候呈显著的系统发育保守格局(图3)。
参考文献(References)
[1]竺可桢.中国近五千年来气候变迁的初步研究.考古学报,1972(1):15-38.
[2]Blomberg S P,Garland T Jr.Tempo and mode in evolution:Phylogenetic inertia,adaptation and comparative methods.Journal of Evolutionary Biology,2002,15(6):899-910.
[3]Whitehead D R.Wind pollination in the angiosperms:Evolutionary and environmental considerations.Evolution,1969,23(1):28-35.
[4]Ashton P S,Givnish T J,Appanah S.Staggered flowering in the Dipterocarpaceae:New insights into floral induction and the evolution of mast fruiting in the aseasonal tropics.American Naturalist,1988,132(1):44-66.
[5]Franks S J,Sim S,Weis A E.Rapid evolution of flowering time by an annual plant in response to a climate fluctuation.Proceedings of the National Academy of Sciences,2007,104(4):1278.
[6]Pianka E R.The structure of lizard communities.Annual Review of Ecology and Systematics,1973,4(1):53-74.
[7]Rabinowitz D,Weaver J C.Phenological properties of wind- and insect-pollinated prairie plants.Ecology,1981,62(1):49-56.
[8]Mantel N.The detection of disease clustering and a generalized regression approach.Cancer Research,1967,27(1-2):209.
[9]Webb C O,Donoghue M J.Phylomatic:Tree assembly for applied phylogenetics.Molecular Ecology Notes,2005,5(1):181-183.
[10]Davies T J,Barraclough T G,Chase M W,Soltis P S,Soltis D E,Vincent S.Darwin’s abominable mystery:Insights from a supertree of the angiosperms.Proceedings of the National Academy of Sciences of the United States of America,2004,101(6):1904-1909.
[11]Webb C O.Ackerly D D.Kembel S W.Phylocom:Software for the analysis of phylogenetic community structure and trait evolution.Bioinformatics,2008,24(18):2098.
[12]R Development Core Team.R:A Language And Environment For Statistical Computing.Vienna,Austria:R Foundation for Statistical Computing,2009.
[13]Kudo G.Flowering phenologies of animal-pollinated plants:Reproductive strategies and agents of selection.In:Harder L D,Barrett S C H.(eds).Ecology and Evolution of Flowers.Oxford,UK:Oxford University Press,2006.
[14]Diekmann M.Relationship between flowering phenology of perennial herbs and meteorological data in deciduous forests of Sweden.Canadian Journal of Botany,1996,74(4):528-537.
[15]White L M.Predicting flowering of 130 plants at 8 locations with temperature and day length.Journal of Range Management,1995,48(2):108-114.
[16]Price M V,Waser N M.Effects of experimental warming on plant reproductive phenology in a subalpine meadow.Ecology,1998,79(4):1261-1271.
[17]Alatalo J M A,TotlandØ.Response to simulated climatic change in an alpine and subarctic pollen-risk strategist,Sileneacaulis.Global Change Biology,1997,3(S1):74-79.
[18]Henry G H R,Molau U.Tundra plants and climate change:The international tundra experiment (ITEX).Global Change Biology,1997,3(S1):1-9.
[19]Stenström M,Gugerli F,Henry G H R.Response ofSaxifragaoppositifoliaL. to simulated climate change at three contrasting latitudes.Global Change Biology,1997,3(S1):44-54.
[20]Suzuki S,Kudo G.Short-term effects of simulated environmental change on phenology,leaf traits,and shoot growth of alpine plants on a temperate mountain,northern Japan.Global Change Biology,1997,3(S1):108-115.
[21]Sherry R A,Sherry R A,Xuhui Z,Shiliang G.Divergence of reproductive phenology under climate warming.Proceedings of the National Academy of Sciences,2007,104(1):198.
[22]Sola A J,Ehrlén J.Vegetative phenology constrains the onset of flowering in the perennial herbLathyrusvernus.Journal of Ecology,2007,95(1):208-216.
[23]Bolmgren K.Adaptation and Constraint in the Plant Reproductive Phase.Botaniska Institutionen,2004.
[24]Bolmgren K,Eriksson O,Linder H P.Contrasting flowering phenology and species richness in abiotically and biotically pollinated angiosperms.Evolution,2003,57(9):2001-2011.
[25]Dahlgren J P,Eriksson O,Bolmgren K.Specific leaf area as a superior predictor of changes in field layer abundance during forest succession.Journal of Vegetation Science,2006,17(5):577-582.
[26]Kolb A,Barsch F,Diekmann M.Determinants of local abundance and range size in forest vascular plants.Global Ecology and Biogeography,2006,15(3):237-247.
[27]Vile D,Shipley B,Garnier E.A structural equation model to integrate changes in functional strategies during old-field succession.Ecology,2006,87(2):504-517.
[28]Helenurm K,Barrett S C H.The reproductive biology of boreal forest herbs.II.Phenology of flowering and fruiting.Canadian Journal of Botany,1987,65(10):2047-2056.
[29]Rathcke B.Flowering phenologies in a shrub community:Competition and constraints.The Journal of Ecology,1988,76(4):975-994.
[30]Kudo G.Relationships between flowering time and fruit set of the entomophilous alpine shrub,Rhododendronaureum(Ericaceae),inhabiting snow patches.American Journal of Botany,1993,80(11):1300-1304.
[31]Primack R B.Longevity of individual flowers.Annual Review of Ecology and Systematics,1985,16:15-37.
[32]Ollerton J,Lack A.Relationships between flowering phenology,plant size and reproductive success in shapeLotuscorniculatus(Fabaceae).Plant Ecology,1998,39(1):35-47.
[33]Elzinga J A,Anne A,Arjen B,Luc G,Arthur E W,Giorgina B.Time after time:Flowering phenology and biotic interactions.Trends in Ecology & Evolution,2007,22(8):432-439.
[34]Waser N M.Interspecific pollen transfer and competition between co-occurring plant species.Oecologia,1978,36(2):223-236.
[35]Galen C,Gregory T.Interspecific pollen transfer as a mechanism of competition:Consequences of foreign pollen contamination for seed set in the alpine wildflower,Polemoniumviscosum.Oecologia,1989,81(1):120-123.
[36]Poole R W,Rathcke B J.Regularity,randomness,and aggregation in flowering phenologies.Science,1979,203:470.
[37]Thomson J D.Patterns of visitation by animal pollinators.Oikos,1982,39(2):241-250.
[38]Ackerly D D,Reich P B.Convergence and correlations among leaf size and function in seed plants:A comparative test using independent contrasts.American Journal of Botany,1999,86(9):1272-1281.
[39]Fenner M.The phenology of growth and reproduction in plants.Perspectives in Plant Ecology,Evolution and Systematics,1998,1(1):78-91.
[40]Levin D A.Flowering phenology in relation to adaptive radiation.Systematic Botany,2006,31(2):239-246.
[41]Stiles F G.Coadapted competitors:The flowering seasons of hummingbird-pollinated plants in a tropical forest.Science,1977,198:1177-1178.
[42]Snow D W.A possible selective factor in the evolution of fruiting seasons in tropical forest.Oikos,1965,15(2):274-281.
[43]Oberrath R,Bohning-Gaese K.Phenological adaptation of ant-dispersed plants to seasonal variation in ant activity.Ecology,2002,3(5):1412-1420.
(责任编辑武艳培)
Plants flowering phenology in Gannan alpine meadow
Zhang Chun-hua1, Zhang Zhi-long1, Jia Peng2
(1.Gansu Normal University for Nationalities, Hezuo 747000, China;2.State Key Laboratory of Grassland Agro-ecosystems, School of Life Sciences,Lanzhou University, Lanzhou 730020, China)
Abstract:The research on the phenology of the grassland community could help cognize the function of the grassland community, and provide the background data for the ecosystem assessment. In this study, the factors that affect the flowering phenology of alpine meadow in Gannan were discussed in four aspects: geographical environment, functional traits, pollination modes and phylogeny. The results showed that, at the community level, the flowering period covered the whole growing season, but there was still a peak flowering period in the whole species. The first flowering date, peak flowering date and duration of flowering time were different. Each species has unique pattern in flowering phenology; Plants height(P<0.001) and seed size(P=0.034) could significantly predict flowering time; Wind-pollinated species flowered earlier than insect-pollinated species(P=0.008);There was a negative correlation between flowering phenology overlaps and phylogenetic distances.
Key words:alpine meadow; flowering phenology; function traits; pollination; phylogeny
Corresponding author:Zhang Chun-huaE-mail: zchjry@126.com
中图分类号:Q948.15+8;Q944.43
文献标识码:A
文章编号:1001-0629(2016)2-0283-07
通信作者:张春花(1968-),女,河南永城人,副教授,本科,主要从事区域资源环境与可持续发展研究。E-mail: zchjry@126.com
基金项目:国家自然科学基金重点资助项目(90202009)
*收稿日期:2015-10-10接受日期:2015-12-25
DOI:10.11829/j.issn.1001-0629.2015-0552
张春花,章志龙,贾鹏.甘南高寒草甸群落花期物候研究.草业科学,2016,33(2):283-289.
Zhang C H,Zhang Z L,Jia P.Plants flowering phenology in Gannan alpine meadow.Pratacultural Science,2016,33(2):283-289.
