种植密度和施氮水平对燕麦生物量分配的影响
2016-04-18赵宏魁张春辉雷占兰姚步青周华坤
赵宏魁,马 真,张春辉,雷占兰,3,姚步青,周华坤
(1.海南州少数民族科普和信息中心,青海 恰卜恰 813099; 2.中国科学院西北高原生物研究所,
青海 西宁 810001; 3.青海师范大学,青海 西宁 810008)
种植密度和施氮水平对燕麦生物量分配的影响
赵宏魁1,2,马 真2,张春辉2,雷占兰2,3,姚步青2,周华坤2
(1.海南州少数民族科普和信息中心,青海 恰卜恰 813099; 2.中国科学院西北高原生物研究所,
青海 西宁 810001; 3.青海师范大学,青海 西宁 810008)
摘要:本研究基于在中国科学院海北高寒草甸生态系统定位站进行的盆栽试验,分析了不同种植密度和施氮水平对燕麦(Avena sativa)(巴燕3号)的生长特性、生物量积累及生物量分配格局的影响。结果表明,不同密度处理下,燕麦的植株高度、叶的长度、穗的长度、分蘖个数、繁殖生物量(穗)、茎叶生物量及地下生物量(根)等各性状参数均呈现极显著差异(P<0.01);繁殖器官生物量分配比例随密度的增加呈上升趋势,根生物量分配随密度的增加逐渐减少;随种植密度增大,燕麦茎叶生物量分配呈现先减少后增加的趋势。氮素的施用有利于燕麦营养生长和繁殖生长生物量的积累。施氮肥对燕麦茎叶器官的生物量累积作用明显大于地下根器官。对照(F1)和低氮肥(0.6 g·盆(-1),F2)大致相等,中氮肥(1 g·盆(-1),F3)和高氮肥(2 g·盆(-1),F4)大致相等,且前两者大于后两者,即在不施氮肥条件下,燕麦增加了对茎叶、根器官生物量的分配比例。在不同处理下,燕麦地下生物量分配、茎叶生物量分配及繁殖生物量分配三者间存在权衡。总体来看,不同处理下,燕麦选择最佳资源配置策略,即增大与获取限制资源相关器官的资源分配,而减少与获取非限制资源相关器官的资源分配,以适应生物环境(例如密度)和非生物环境(例如氮素添加)的变化。
关键词:燕麦;种植密度;氮肥添加;繁殖分配;生物量分配;高寒草甸
生物量分配是生物量在植物不同器官或功能单位(根、茎、叶、地上或地下部分等) 中的分配方式。植物根茎叶生物量等比重的变化不但会影响植物对光、养分和水分的获取等功能[1-2],还会影响群落的结构和功能[3]。另外,繁殖分配是生物量分配研究中最为重要的内容之一。繁殖分配是植物在生长发育过程中,繁殖器官获得同化产物的比例,控制着植物繁殖与生存的平衡[4],体现了物种当前的繁殖力,还能推测出种群潜在的繁殖力,是植物繁殖生态学的研究热点之一[5-6]。在栽培草地的建植中,生物量分配(包括繁殖分配)会显著影响建植物种的产量、质量和稳定性。然而,生物量分配又常常受到生物因素(例如种植密度)和环境因素(例如土地肥力)的影响[7-8],因此,需要了解建植物种在不同种植密度和施肥水平等干扰背景下的生物量分配特征,以便能更好地建植和管理栽培草地。
大量实践证明,发展栽培草地是解决青藏高原高寒草地高效生产和持续发展矛盾的重要措施之一。燕麦(Avenasativa)是青藏高原高寒牧区栽培草地的主要栽培种,现已成为高寒地区优选牧草被大力推广种植[9-10]。对燕麦的相关研究已经有一些报道[11-13],主要集中在品比试验和杂交选育等方面。不同栽培措施对燕麦生物量分配会产生不同的效应,但这方面的研究主要为一些单因素的研究[14-15]。例如,朱志红和王刚[14]对不同种植密度燕麦的繁殖分配进行了研究;德科加等[15]对施氮条件下的有效分蘖数、小花数、小穗数、单序籽粒重等进行了研究。但种植密度和氮添加对燕麦生物量分配影响的研究尚未有相关报道。
密度依赖性死亡或正相互作用一般普遍存在于植物种群或群落[16-18],因而选择合适的种植密度是建植和管理栽培草地的关键之一。另外,施肥是提高栽培草地产量和稳定性的重要方式[19]。鉴于此,本研究探讨高寒地区特有的生境条件下,不同种植密度和施氮水平对燕麦繁殖分配及生长特性的影响,以此探讨上述两种因素对一年生栽培草地群落的结构功能及维持机制的影响,最为重要的是期望对高寒地区栽培草地的建植和管理在种植密度和施肥两个方面提出建设性的建议。
1材料与方法
1.1研究区概况
试验于中国科学院海北高寒草甸生态系统定位站进行(以后简称海北站,位于37°37′ N、101°19′ E,海拔3 200 m)。海北站地处青藏高原东北缘,祁连山北支冷龙岭东段南麓的大通河河谷。年均气温-1.7 ℃,最热月(7月)平均气温为9.8 ℃,最冷月(1月)平均气温为-15.2 ℃。年均降水量582 mm,主要分布于5-9月。无明显四季之分,只有冷暖两季之别,表现出冷季雨量较少,干燥;暖季雨量较高,湿润。
1.2研究方法
试验于2011年5月底在海北高寒草甸生态系统定位站野外进行,燕麦种子来自青海省牧草良种繁殖场。花盆内土壤来自于原生境的草毡寒冻雏形土,有机质含量丰富。选用29 cm×20 cm(直径×高度)聚乙烯花盆进行盆栽,随机区组设计。播种前将种子提前3天浸泡催芽,将催芽的种子按设定密度点播于盆内。设密度、氮肥2因素4水平,共8处理,每个处理24个重复,共192个花盆。4个密度处理分别为D1(5粒·盆-1)、D2(20粒·盆-1)、D3(35粒·盆-1)和D4(50粒·盆-1)[20]。4个施肥梯度分别为对照组F1(不施肥)、F2(0.6 g·盆-1)、F3(1 g·盆-1)和F4(2 g·盆-1)(表1),所施氮肥为尿素。4 个氮肥处理水平播种密度均为28粒·盆-1,播种深度2~3 cm。7月初燕麦苗出齐后,进行氮肥处理。人工除去花盆内杂草,一个月去除杂草一次。
9月初进行取样调查(此时燕麦处于果期)。取样前预先测量燕麦株高,记录分蘖数;测量叶长、穗长。将植株分为根、茎、叶和穗在65 ℃条件下烘48 h后称重(电子天平精确度0.001 g)。每个处理取6个重复。
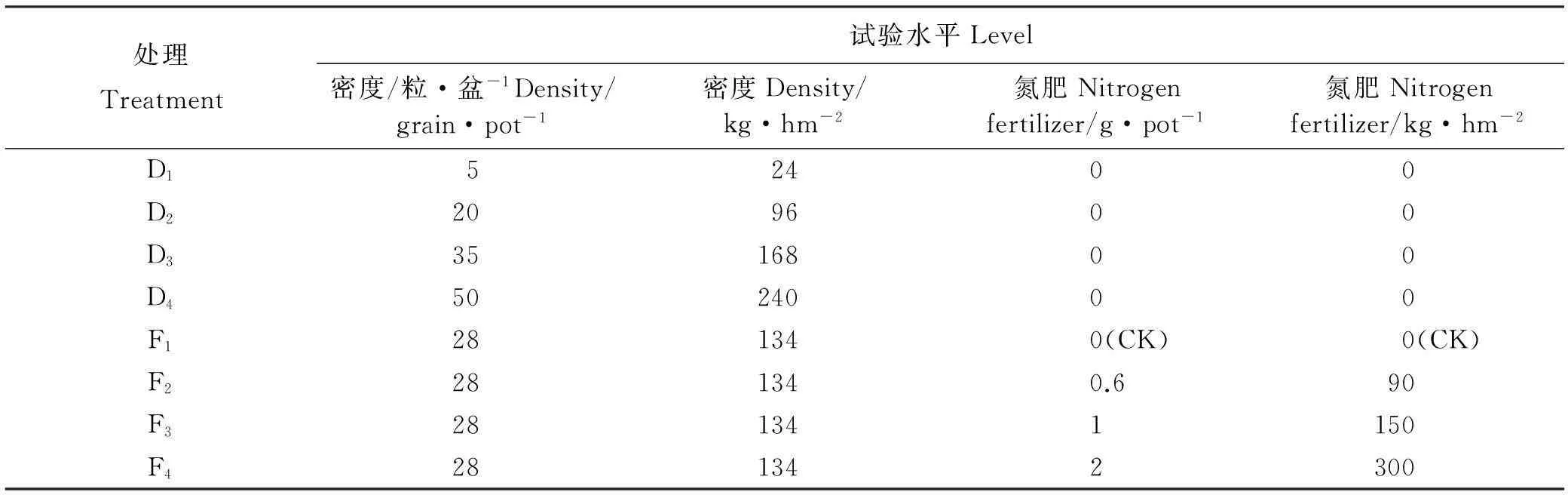
表1 试验设计
1.3数据处理
不同处理下,应用Pearson相关分析法分析燕麦各性状的相关性。不同处理下,应用一般线性回归分析繁殖生物量、茎叶生物量和地下生物量间的相互关系,从而分析三者之间的权衡。以植株不同器官的干重占植株总干重的比来评价这一器官的资源分配,例如繁殖生物量分配=穗生物量/总生物量。描述性统计分析使用Microsoft Excel;方差分析及LSD组间比较、相关分析和一般线性回归使用SPSS 17.0软件进行。
2结果
2.1种植密度和施肥水平对燕麦生长量的影响
密度对植物个体的外部性状指标均有极显著影响(P<0.01)。随着播种密度的增加,株高、叶长、穗长、分蘖数、地下生物量、茎叶生物量和繁殖生物量均呈现显著降低趋势(P<0.05,表2),且所有性状都在D1处理达最大值。氮肥处理对株高、叶长、穗长、分蘖数、地下生物量(根)、茎叶生物量及繁殖生物量(穗)均有极显著影响(P<0.01,表3),且所有性状在高氮肥水平(F4)达到最大值,而且施氮肥的3个处理都大于对照组(F1)。株高、叶长、穗长、分蘖数及各器官生物量随着施氮量的增加有增加的趋势。各氮肥处理中,F2、F3处理下各器官生物量的积累很接近,说明生物量积累在这两个氮肥处理下生长状况一致。F1处理下的各器官生物量和F2、F3及F4处理之间差异均显著(P<0.05),且F3和F4处理差异也显著(P<0.05)。
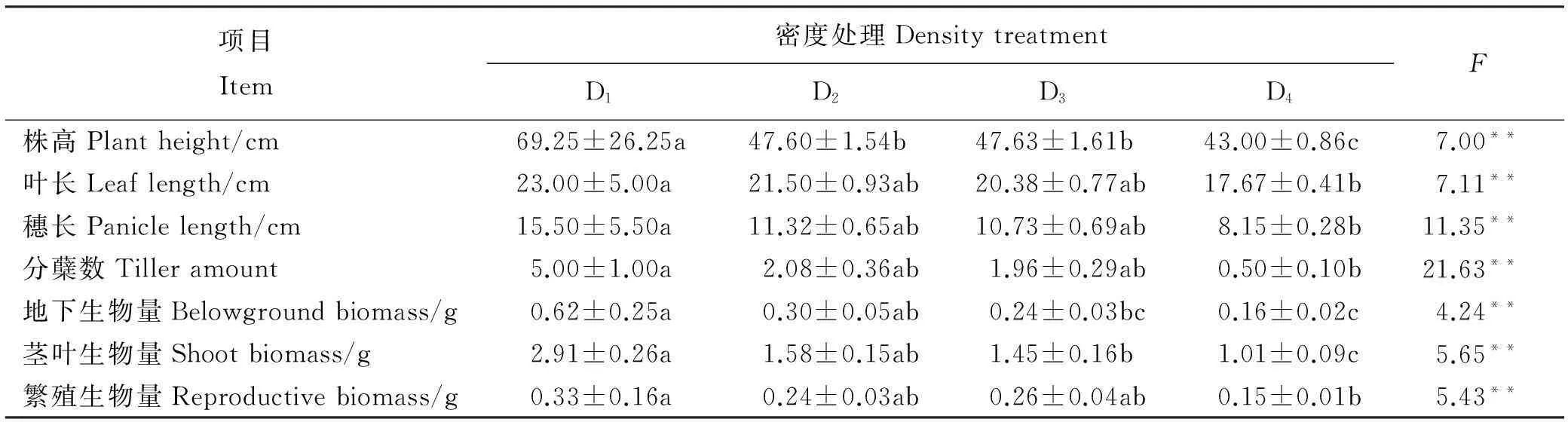
表2 不同播种密度处理下燕麦的生长指标差异
注: 数据表示平均值±标准误。同行不同的小写字母表示不同处理间存在显著差异(P<0.05)。*,P<0.05;**,P<0.01。表3、表4、表5同。
Note: Values are means±SE. Different lower case letters within the same row indicate significant difference among different treatments at 0.05 level.*,P<0.05; **,P<0.01. The same in Table 3,Table 4 and Table 5.
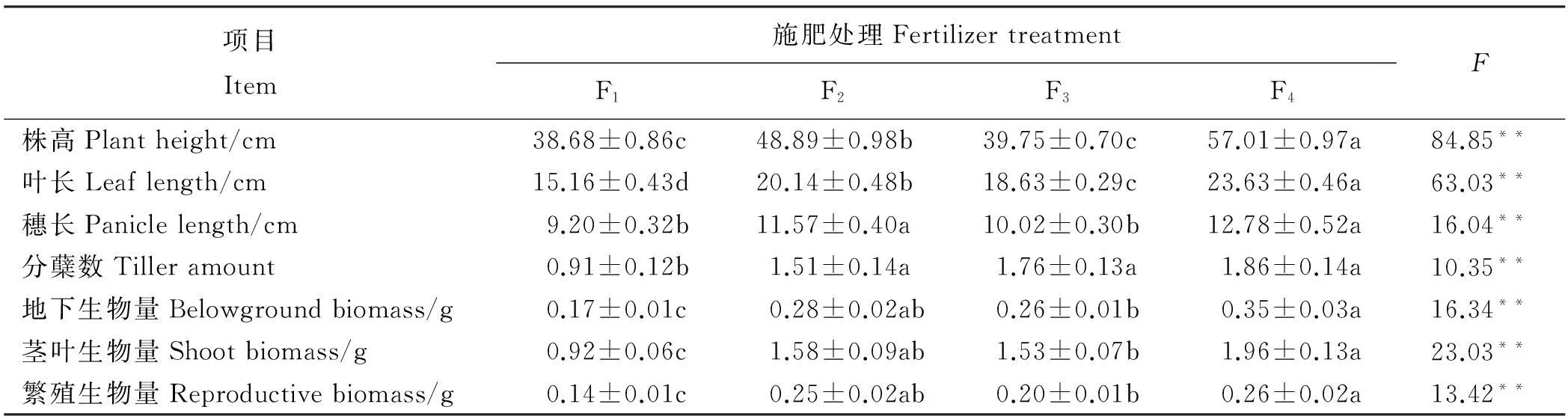
表3 不同氮肥施加水平下燕麦的生长指标差异
2.2不同处理下燕麦生长量之间的相关性
在密度、氮肥单因子作用下,各性状之间呈极显著正相关(P<0.01)。地下生物量与茎叶生物量的相关系数在密度处理下最高(r=0.906);穗长和分蘖数、地下生物量之间的相关系数较小(r<0.400)(表4)。氮肥处理下,所有性状间都极显著正相关,其中地下生物量和茎叶生物量相关系数最大(r=0.887),同时繁殖生物量和穗长的相关性也很高(r=0.882)(表5)。
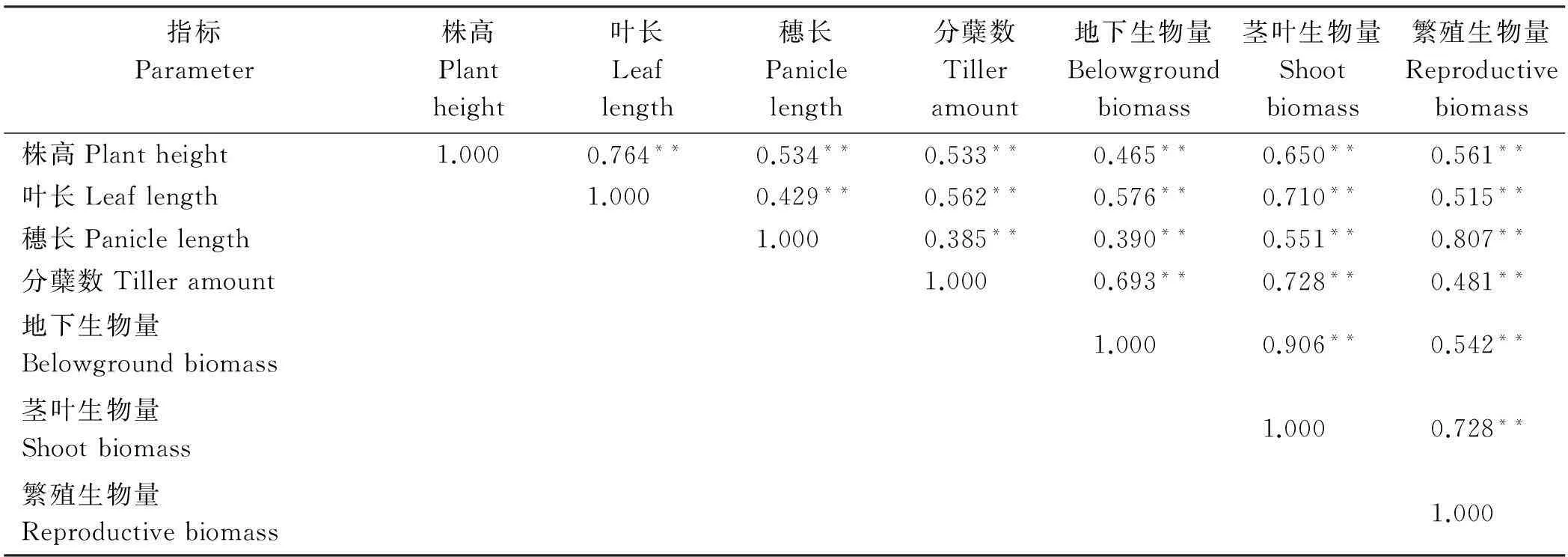
表4 不同播种密度下燕麦各性状间的相关性
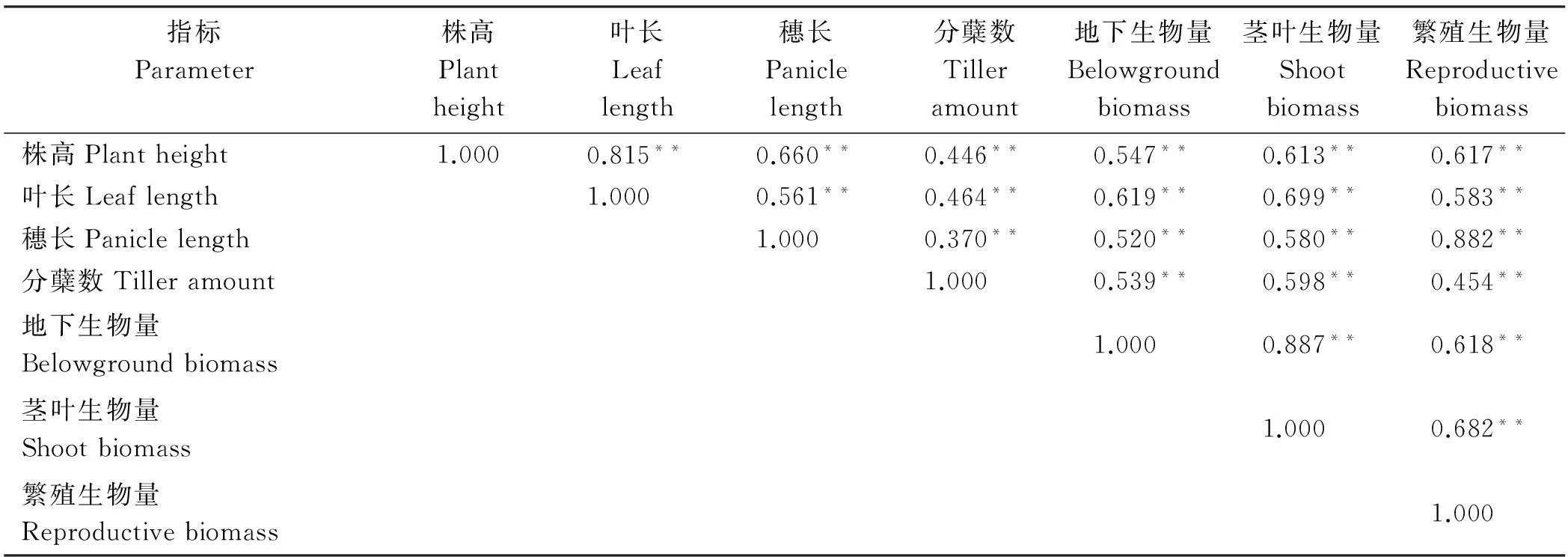
表5 氮肥处理下燕麦各生长性状之间的相关性
2.3不同处理对燕麦生物量分配的影响
地下生物量分配随密度的增加逐渐减少(表6),且在D4处理下,地下生物量分配显著低于D1和D2处理下的(P<0.05);茎叶生物量分配变化则呈先减少后增加的趋势,在D3处理下达到最低,且其显著低于D4处理下的。从繁殖分配来看,在D3处理下最高,达到13.72%,但处理间都未达到显著差异(P>0.05)。
地下生物量分配比例随施氮量的增加先减少后增加,在F3处理下最小,但处理间都未达到显著差异(P>0.05);茎叶生物量分配变化与地下根器官呈相反趋势,在F3处理下达到最大值(表6),且其显著高于F1和F2处理下的。繁殖器官的分配表现为F2>F1>F4>F3,但只有F2和F3处理间差异显著(P<0.05)。
2.4不同种植密度下燕麦生物量分配的权衡
2.4.1繁殖生物量分配与茎叶生物量分配间的权衡D2、D3和D4处理中,燕麦的繁殖生物量分配与茎叶生物量分配均表现为显著负相关(P<0.01,表7),但燕麦的繁殖生物量分配与茎叶生物量分配在D2、D3、D4密度的生境中存在着资源分配权衡。在这3个密度的生境中,随着密度的增加,燕麦的繁殖生物量分配呈先增加后降低的趋势,茎叶生物量分配呈现先降低后升高的趋势(表6),二者之间发生了竞争。繁殖生物量分配在D3密度处理下最大,茎叶生物量分配在D4密度处理下最大。
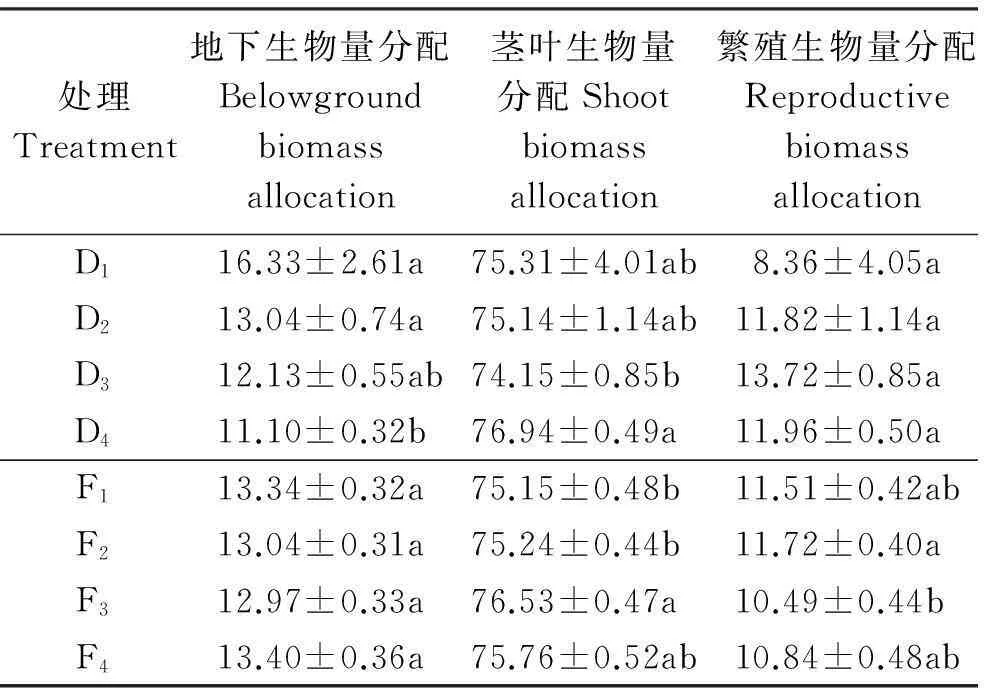
表6 不同处理下燕麦各器官生物量分配
注:同列不同小写字母表示密度或肥料处理间差异显著(P<0.05)。
Note: Different lower case letters within the same column indicate significant difference among different density or fertilizer treatments at 0.05 level.
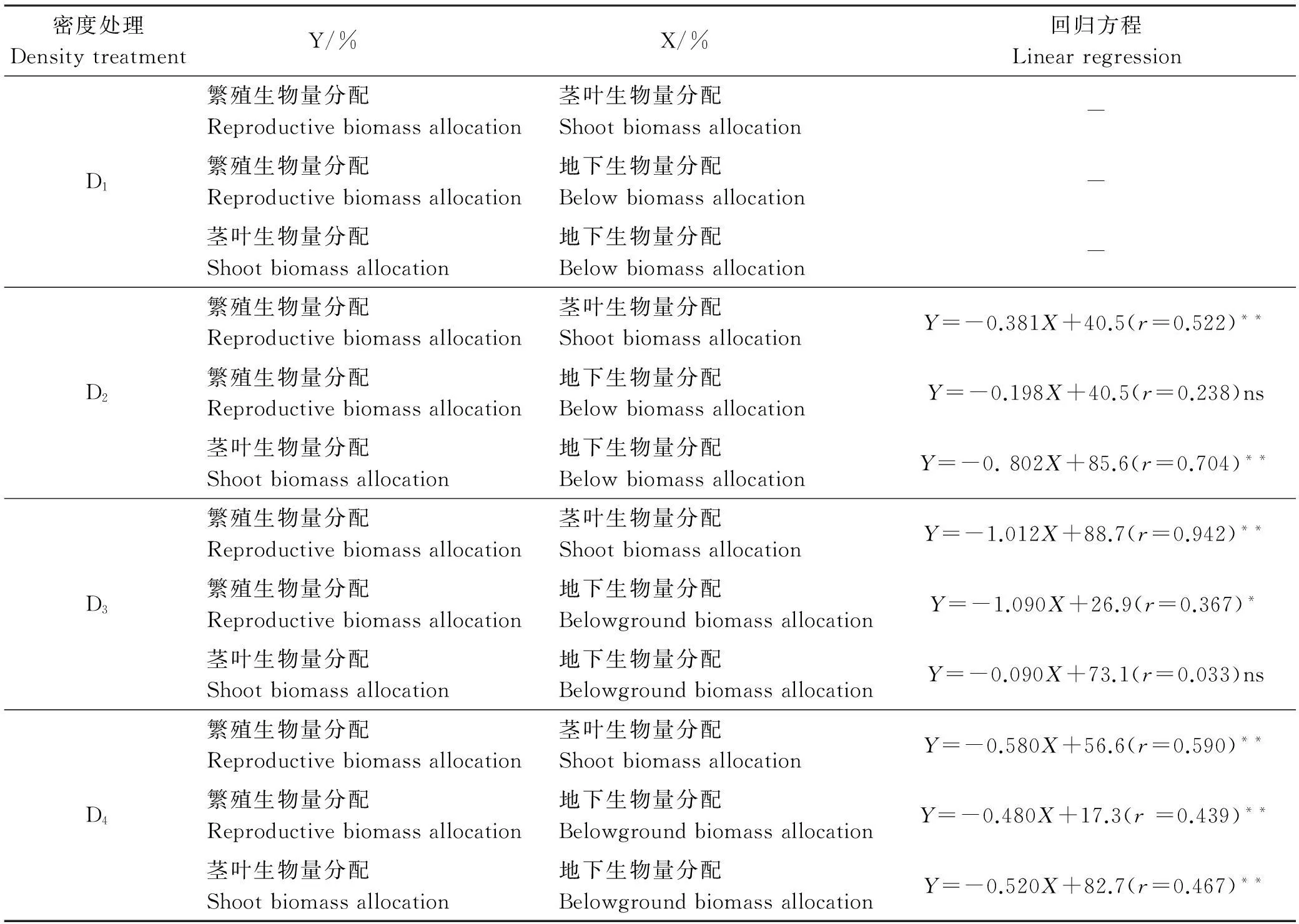
表7 不同播种密度下燕麦各器官生物量分配的相互关系
注:*,P<0.05;**,P<0.01;ns,P>0.05。“-”表示样本量不足(n<5)未做回归。表8同。
Note: *,P<0.05; **,P<0.01; ns,P>0.05. “-” mean no regression (n<5). The same in Table 8.
2.4.2繁殖生物量分配与地下生物量分配的权衡在较高密度(D4和D3)时,燕麦的繁殖生物量分配和地下生物量分配存在极显著(P<0.01)或者显著的负相关 (P<0.05,表7),而在D2密度处理中为非显著性负相关(P>0.05),所以总体上来说二者存在资源分配的权衡问题。地下生物量分配随着密度的增加呈减少的趋势(表6);在D3密度处理下其繁殖生物量分配达到了最大,在D4密度处理下降低。
2.4.3茎叶生物量分配与地下生物量分配间的权衡在D2和D4密度处理下,燕麦的茎叶生物量分配和地下生物量分配都为极显著负相关(P<0.01,表7),即这一结果说明茎叶生物量分配与地下生物量分配间存在权衡。而在D3密度处理下无显著性相关(P>0.05)。
2.5不同施氮水平下燕麦生物量分配之间的权衡
2.5.1繁殖生物量分配与茎叶生物量分配之间的权衡在氮肥的4个水平中,燕麦的繁殖生物量分配与茎叶生物量分配均为极显著负相关(P<0.01,表8)。在F1、F2、F3、F4的生境中,燕麦的繁殖生物量分配和茎叶生物量分配存在着资源分配权衡。繁殖生物量分配在F2氮肥处理下最大(表6),说明F2氮肥处理更有利于繁殖器官的生长,茎叶生物量分配在F3氮肥处理下最大,说明氮肥量的增加促进了燕麦茎叶等营养器官生物量的增长,但F3处理下繁殖生物量分配最小。
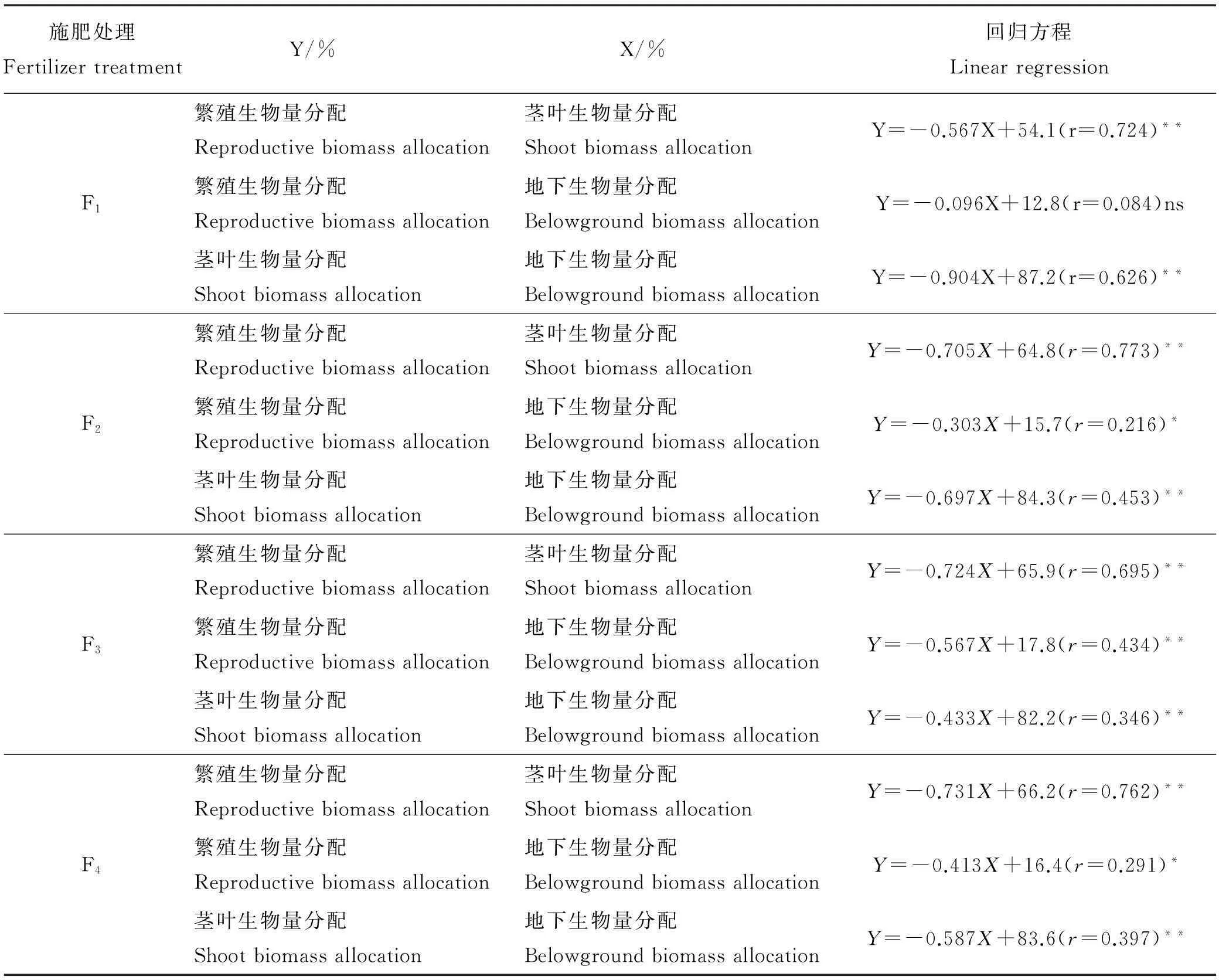
表8 不同氮肥量施加水平下燕麦各器官生物量分配间的相互关系
2.5.2繁殖生物量分配与地下生物量分配间的权衡在中氮肥(F3)时,燕麦的繁殖生物量分配和地下生物量分配呈极显著负相关 (P<0.01,表8),在F2和F4氮肥处理中为显著的负相关(P<0.05),在F1氮肥处理下相关性不显著(P>0.05)。随着氮肥量的增加,地下生物量分配呈先减少后增加的趋势,而繁殖生物量分配呈现先增加后减少的趋势(表6),所以二者之间存在权衡。
2.5.3茎叶生物量分配与地下生物量分配之间的权衡在氮肥处理的4个水平下,燕麦的茎叶生物量分配和地下生物量分配都为极显著负相关(P<0.01,表8)。随着氮肥量的增加,茎叶生物量分配先增加后减少,地下生物量分配先减少后增加。在氮肥F3处理下茎叶生物量分配最高,而地下生物量分配最少,所以二者之间存在权衡。
3讨论
3.1种植密度和氮肥添加对燕麦生长特性的影响
由于植物固着性的生活方式,不能像动物那样自由移动选择适宜的生境,因而各种养分胁迫和逆境是无法避免的[10]。但作为构件生物的植物却可以通过改变构件组成特征来适应外界环境,以此来影响植物对资源的利用[21]。在本研究中,燕麦各性状间在密度因子和氮肥因子作用下都极显著正相关(P<0.01)。
密度制约在植物种群中普遍存在,是种群极其重要的选择压力[17]。密度影响着种群内个体获得资源的量,由此引起邻近植株的光资源、水分和营养竞争[22]。种群密度可以显著影响植物种群内个体的特征[23-24]。本研究也同样表明,密度对株高、分蘖数、叶长、穗长、地下生物量、茎叶生物量和繁殖生物量各性状参数均影响极显著(P<0.01,表2),所有的性状都在D1处理下达到最大值,表明密度制约效应极其明显。本研究结果与杨允菲和张宝田[25]对松嫩平原碱化草甸天然虎尾草(Chlorisvirgata)种群密度制约特征的研究结果一致。随着密度的增加,植株间的竞争变得更加激烈,在资源有限的情况下,种群内个体可获得的资源量减少,从而引起各性状参数的变化。
氮元素是植物必不可少的营养元素之一,它同时影响着植物对其它营养元素的吸收[26-27]。植物的生长规律及繁殖分配格局因为土壤不同的含氮量而发生着变化[28]。氮肥处理对植株高度、叶的长度、穗的长度、分蘖个数、繁殖生物量、茎叶生物量及地下生物量等性状均有极显著影响(P<0.01,表3),所有性状在F4处理下达到最大值,说明不同的氮肥量显著影响了燕麦的生长特征及繁殖分配格局。
3.2不同处理对燕麦生物量分配的影响
生物量的累积和分配体现了密度对植物种群的影响[7]。植物通过地上和地下生物量的分配模式来适应养分和光照等资源的变化,从而表现出对某些资源的不同的竞争能力[10]。一般来说,较高的地上(茎叶)生物量分配表明植物对光照的需求及高的光竞争能力[29],而较高的地下生物量表明植物对土壤养分有着较高的需求及强的地下竞争力[30]。本研究表明,随着播种密度的增加,燕麦繁殖器官生物量分配比例总体上呈上升趋势,但分配比例均低于20%。这与已有的结论(即一年生植物繁殖分配比例高于20%)[31]不一致,这可能与本研究所取的农田熟土有关。地下生物量分配随播种密度的增加逐渐减少,茎叶生物量分配变化则先减少后增加的趋势,这表明不同播种密度对燕麦地上和地下竞争均有一定的影响。
研究表明,植物的生长规律及繁殖分配格局会因氮肥量的不同而异[7]。在一定范围内,氮素的增加可以明显提高种群的产量,但若超过一定限度,产量不再持续增加[32-33],即产量存在一个阈值。本研究也证实了上述结论。增加氮素有利于植株叶面积的增加,从而提高光合产量[34]。随着施氮量的增加,本研究发现,燕麦地下生物量分配先减少后增加(表6),而茎叶生物量分配变化与地下根器官呈相反的趋势。
3.3燕麦根、茎叶及穗生物量分配三者的权衡
植物生活史理论认为繁殖生长与营养生长两者之间存在权衡[31]。关于植物繁殖分配模式与密度的关系,以前的研究并未得出一致的结论。这些观点主要可以概括为3类:1)高密度下植物将更多的资源分配给繁殖器官;2)高密度下植物将更多的资源分配给营养器官;3)植物分配到繁殖器官和营养器官的资源比例不受密度的影响,即繁殖分配比例为一定值[35-36]。本研究认为,燕麦属于第3类观点,即繁殖分配比例为定值。燕麦的地下生物量分配、茎叶生物量分配及繁殖生物量分配在不同密度处理下三者间存在权衡关系。在不同种植密度下,不论三者之间如何权衡分配,繁殖分配的比例始终在8.36%~13.72%变化,且未表现出显著差异(表6、表7)。
植物繁殖的成功可以通过控制分配用于营养生长和繁殖生长的氮元素实现。繁殖分配理论表明,在资源有限条件下,用于营养生长和繁殖生长的资源表现为负相关,资源可调节现象使植物各器官权衡生长[7-8,10]。在不同氮肥添加水平下,本研究也同样发现燕麦的资源分配在三者间(地下生物量分配、茎叶生物量分配和繁殖生物量分配)存在权衡关系(表6、表8)。
4结论
不同密度处理下,燕麦植株各性状参数均有极显著差异(P<0.01),且所有性状都在5粒·盆-1处理下达到最大值;繁殖器官生物量分配比例总体上呈上升趋势;地下(根)生物量分配随密度的增加逐渐减少;燕麦茎叶生物量分配呈现先减少后增加的趋势。综上所述,如以获取燕麦茎叶为目的,播种密度以240 kg·hm-2左右为益,而如果以收获燕麦种子为目的,种植密度以168 kg·hm-2左右为最佳。氮肥处理下,所有性状在高氮肥水平2 g·盆-1下显著大于其它处理(P<0.05);随施氮量的增加,地下生物量分配比例先减少后增加;茎叶生物量分配变化与地下根器官呈相反趋势。如以获得种子高产,建议施氮量为90 kg·hm-2最佳。总之,燕麦的繁殖分配比例较为恒定,且在不同处理下地下生物量分配、茎叶生物量分配及繁殖生物量分配三者间存在权衡,以适应生物环境(例如密度)和非生物环境(例如氮素添加)的变化。
参考文献(References)
[1]Norby R J,Todd D E,Fults J,Johnson D W.Allometric determination of tree growth in a CO2-enriched sweetgum stand.New Phytologist,2001,15:477-487.
[2]Hautier Y,Niklaus P A,Hector A.Competition for light causes plant biodiversity loss after eutrophication.Science,2009,324:636-638.
[3]Keddy P,Fraser L H,Wisheu I C.A comparative approach to examine competitive response of 48 wetland plant species.Journal of Vegetation Science,1998(9):777-786.
[4]钟章成.植物种群的繁殖对策.生态学杂志,1995,14(1):37-42.
Zhong C Z.Reproduetive strategies of plant populations.Chinese Journal of Ecology,1995,14(1):37-42.(in Chinese)
[5]郭伟,邓巍,燕雪飞,潘星极,陈红霞.植物生殖分配影响因素研究进展.东北农业大学学报,2010,41(9):150-155.
Guo W,Deng W,Yan X F,Pan X J,Chen H X.Research advances on impact factors of plant reproductive allocation.Journal of Northeast Agricultural University,2010,41(9):150-155.(in Chinese)
[6]张大勇.植物生活史进化与繁殖生态学.北京:科学出版社,2004.
Zhang D Y.Plant Life History Evolution and Reproductive Ecology.Beijing:Sicience Press,2004.(in Chinese)
[7]牛建伟,雷占兰,周华坤,刘泽华,黄瑞灵,姚步青,王文颖,赵新全.种植密度和施氮水平对垂穗披碱草生物量分配的影响.草业科学,2014,31(7):1343-1351.
Niu J W,Lei Z L,Zhou H K,Liu Z H,Huang R L,Yao B Q,Wang W Y,Zhao X Q.Reproductive allocation ofElymusnutansunder different planting densities and nitrogen fertilizing treatments.Pratacultural Science,2014,31(7):1343-1351.(in Chinese)
[8]雷占兰,周华坤,刘泽华,黄瑞灵,王文颖.密度氮肥交互处理下垂穗披碱草生长特性与生殖分配.中国草地学报,2014,36(2):12-19.
Lei Z L,Zhou H K,Liu Z H,Huang R L,Wang W Y.The growth and reproductive characteristics ofElymusnutansunder the interaction between different planting density and nitrogen application.Chinese Journal of Grassland,2014,36(2):12-19.(in Chinese)
[9]马晓刚,任有成,王显萍.发展燕麦生产在青海经济和生态建设中的作用.作物杂志,2004(5):9-11.
Ma X G,Ren Y C,Wang X P.The role of oat production development in economic and ecological construction of Qinhai.Grops,2004(5):9-11.(in Chinese)
[10]雷占兰,周华坤,刘泽华,黄瑞灵,姚步青,王文颖,赵新全.密度氮肥交互处理下高寒地区燕麦的生长特性与生殖分配.草业科学,2014,31(6):1110-1119.
Lei Z L,Zhou H K,Liu Z H,Huang R L,Yao B Q,Wang W Y,Zhao X Q.The growth performance and reproductive allocation ofAvenasativaunder density and nitrogen interaction.Pratacultural Science,2014,31(6):1110-1119.(in Chinese)
[11]施建军,马玉寿,李青云,董全民.高寒牧区燕麦高产栽培技术的研究.草原与草坪,2003(4):39-41.
Shi J J,Ma Y S,Li Q Y,Dong Q M.Study on culture technology of oat for high quality and high yield in alpine region.Grassland and Turf,2003(4):39-41.(in Chinese)
[12]董世魁,蒲小朋,马金星,石振田.甘肃天祝高寒地区燕麦品种生产性能评价.草地学报,2001,9(1):44-49.
Dong S K,Pu X P,Ma J X,Shi Z T.Productive evaluation on different oat(Avenasativa) varieties in alpine region of Tianzhu.Acta Agrestia sinica,2001,9(1):44-49.(in Chinese)
[13]于福同,张爱民.植物营养性状遗传研究的进展.作物杂志,1998(1):6-9.
Yu F T,Zhang A M.Current research situation on genetics of plant vegetative traits.Grops,1998(1):6-9.(in Chinese)
[14]朱志红,王刚.燕麦表型可塑性与繁殖分配的研究.兰州大学学报(自然科学版),2002,38(1):76-83.
Zhu Z H,Wang G.Studies on the phenotypic plasticity and reproductive allocation inAvenasativaL. Journal of Lanzhou University(Natural Sciences),2002,38(1):76-83.(in Chinese)
[15]德科加,周青平,刘文辉,徐成体,王德利.施氮量对青藏高原燕麦产量和品质的影响.中国草地学报,2007,29(5):43-48.
De K J,Zhou Q P,Liu W H,Xu C T,Wang D L.Effects of nitrogen application on the yield and quality of oat in Qinghai-Tibet plateau.Chinese Journal of Grassland,2007,29(5):43-48.(in Chinese)
[16]Chu C J,Weiner J,Maestre F,Xiao S,Wang Y S,Li Q,Yuan J L,Zhao L Q,Ren Z W,Wang G.Positive interactions can increase size inequality in plant populations.Journal of Ecology,2009,94:1401-1407.
[17]Japhet W,Zhou D W,Zhang H X.Evidence of phenotypic plasticity in the response ofFagopyrumesculentumto population density and sowing date.Journal of Plant Biology,2009,52:303-311.
[18]张炜平,潘莎,贾昕,储诚进,肖洒,林玥,白燕远,王根轩.植物间正相互作用对种群动态和群落结构的影响:基于个体模型的研究进展.植物生态学报,2013,37(6):571-582.
Zhang W P,Pan S,Jia X,Chu C J,Xiao S,Lin Y,Bai Y Y,Wang G X.Effects of positive plant interactions on population dynamics and community structures:A review based on individual-based simulation models.Chinese Journal of Plant Ecology,2013,37(6):571-582.(in Chinese)
[19]田育红.干扰我国人工草地生态系统的主要影响因素研究.安徽农业科学,2011,39(25):15600-15602.
Tian Y H.Research on the main factors on the ecosystem of artificial grassland in China.Journal of Anhui Agriculture Science,2011,39(25):15600-15602.(in Chinese)
[20]乔有明.不同播种密度对燕麦几个数量性状的影响.草业科学,2002,19(1):31-32.
Qiao Y M.Effects of seeding density on quantitative characters of oat.Pratacultural Science,2002,19(1):31-32.(in Chinese)
[21]贾程,何飞,樊华,蔡蕾,尤海舟,李贤伟,秦嘉励,刘兴良.植物种群构件研究进展及其展望.四川林业科技,2010,31(3):43-50.
Jia C,He F,Fan H,Cai L,You H Z,Li X W,Qin J L,Liu X L.Researches on plant population modules.Journal of Sichuan Forestry Science and Technology,2010,31(3):43-50.(in Chinese)
[22]黎磊,周道玮,盛连喜.密度制约决定的植物生物量分配格局.生态学杂志,2011,30(8):1579-1589.
Li L,Zhou D W,Sheng L X.Density dependence-determined plant biomass allocation pattern.Chinese Journal of Ecology,2011,30(8):1579-1589.(in Chinese)
[23]李雪林,张爱峰,吴忠祥,赵燕良.老芒麦种群密度制约特性初探.青海草业,2001,10(2):9-12.
Li X L,Zhang A F,Wu Z X,Zhao Y L.Characteristic investigation on community density restrain ofElymussibiricus.Qinghai Prataculture,2001,10(2):9-12.(in Chinese)
[24]赵相健,王孝安.太白红杉分枝格局的可塑性研究.西北植物学报,2005,25(1):113-117.
Zhao X J,Wang X A.Study on branching pattern plasticity ofLarixchinensis.Acta Botanica Boreali-Occidentalia Sinica,2005,25(1):113-117.(in Chinese)
[25]杨允菲,张宝田.松嫩平原碱化草甸天然虎尾草种群密度制约特征的研究.生态学报,1992,12(3):266-272.
Yang Y F,Zhang B T.Study on specificities of the density dependence to naturalChlorisvirgatapopulation of alkalization meadow in the Songnen plain of China.Acta Ecologica Sinica,1992,12(3):266-272.(in Chinese)
[26]马立祥,赵甍,毛子军,刘林馨,赵溪竹.不同氮素水平下增温及CO2升高综合作用对蒙古栎幼苗生物量及其分配的影响.植物生态学报,2010,34(3):279-288.
Ma L X,Zhao M,Mao Z J,Liu L X,Zhao X Z.Effects of elevated temperature and CO2under different nitrogen regimes on biomass and its allocation inQuercusmongolicaseedlings.Chinese Journal of Plant Ecology,2010,34(3):279-288.(in Chinese)
[27]陈永春.不同施肥处理对款冬花生物量分配和产量的影响.南方农业,2009(1):55-56.
Cheng Y C.Effects of different fertilizing treatments on biomass allocation and yield ofTussilagofarfara.South China Agriculture,2009(1):55-56. (in Chinese)
[28]王满莲,韦霄,蒋运生,柴胜丰,李锋,漆小雪.氮对黄花蒿生长、光合特性和青蒿素含量的影响.广西植物,2009,29(2):260-263.
Wang M L,Wei X,Jiang Y S,Chai S F,Li F,Qi X X.Effect of soil nitrogen levels on the growth,photosynthetic properties and artemisinin content ofArtemisiaannuaseedling.Guihaia,2009,29(2):260-263. (in Chinese)
[29]Niu K C,Choler P,Zhao B B, Du G Z.The allometry of reproductive biomass in response to land use in Tibetan alpine grasslands.Functional Ecology,2009,23:274-283.
[30]王军邦,王政权,胡秉民,牛铮,王长耀.不同栽植方式下紫椴幼苗生物量分配及资源利用分析.植物生态学报,2002,26(6):677-683.
Wang J B,Wang Z Q,Hu B M,Niu Z,Wang C Y.Biomass allocation and resource use of TiliaAmurensisjuvenileunder different planting treatments.Acta Phytoecologica Sinica,2002,26(6):677-683.(in Chinese)
[31]李金花,潘浩文,王刚.草地植物种群繁殖对策研究.西北植物学报,2004,24(2):352-355.
Li J H,Pan H W,Wang G.Reproductive strategy of grassland plant.Acta Botanica Boreali-Occidentalia Sinica,2004,24(2):352-355. (in Chinese)
[32]王立秋.冀西北春小麦高产优质高效栽培研究——氮磷肥对春小麦产量和品质的影响及效益分析.干旱地区农业研究,1994,12(3):8-13.
Wang L Q.High-yielding,good-quality and high-efficiency cultivation of spring wheat in northwest of Hebei Province——Effects of N and P fertilizer upon grain yields and quality of spring wheat.Agricultural Research in the Arid Aress,1994,12(3):8-13. (in Chinese)
[33]秦亚洲,王利立,柴强,殷文.大麦间作豌豆的种间竞争力及产量对施氮量的响应.农业现代化研究,2015,36(3):482-487.
Qin Y Z,Wang L L,Chai Q,Yin W.Responses of interspecific competition and crop yield to nitrogen applications in a barley-field pea intercropping system.Research of Agricultural Modernization,2015,36(3):482-487.(in Chinese)
[34]Antonopoulos V Z.Modeling of water and nitrogen balances in the ponded water and soil profile of rice fields in Northern Greece.Agricultural Water Management,2010,12(2):321-330.
[35]杜国祯,王刚.垂穗披碱草个体大小与种群密度的关系.植物生态学报,1998,22(6):552-558.
Du G Z,Wang G.Relationship between individual size and density ofElymusnutanspopulation.Acta Phytoecologica Sinica,1998,22(6):552-558.(in Chinese)
[36]Weiner J.The influence of competition on plant reproduction.In:Doust J L,Doust L L.(eds).Plant Reproductive Ecology:Patterns and Strategies.Oxford:Oxford University Press,1988:228-245.
(责任编辑武艳培)
The reproductive allocation ofAvenasativaunder different planting densities and nitrogen addition treatments
Zhao Hong-kui1,2, Ma Zhen2, Zhang Chun-hui2, Lei Zhan-lan2,3,Yao Bu-qing2, Zhou Hua-kun2
(1.Hainan State Ethnic Minority Science and Information Center, Qiabuqia 813099, China;2.Northwest Plateau Institute of Biology, Chinese Academy of Sciences, Xining 810001, China;3.Qinghai Normal University, Xining 810008, China)
Abstract:To study the response of the growth characteristics, biomass accumulation and biomass allocation pattern of Avena sativa to planting density and nutrient addition, a pot experiment was conducted with different planting densities and different levels of nitrogen addition. The results showed that the plant height, leaf length, panicle length, tiller number, root biomass, shoot biomass and reproductive biomass of A. sativa differed significantly (P<0.01) in the different density levels; as plant density increasing, reproductive biomass allocation increased, while belowground biomass (root) allocation decreased; shoot (leaf and stem) biomass allocation decreased along with plant density and then increased afterwards. Nutrient addition increased both vegetative and reproductive biomass. Nutrient addition had more effect on shoot biomass than root biomass. Reproductive biomass allocation: F1 (no nutrient addition) ≈ F2 (low nutrient addition) >F3 (medium nutrient addition) ≈ F4 (high nutrient addition), i.e. A. sativa exhibited high root biomass allocation and shoot biomass allocation in F1. In general, to obtain limited resources and optimal biomass allocation and to adapt to biotic (e.g. plant density) and physical (e.g. nitrogen addition) environment changes, there’s a trade-off between the below-ground biomass allocation and between vegetative-reproductive biomass allocation of A. sativa in different treatments.
Key words:Avena sativa; planting density; nitrogen addition; reproductive allocation; biomass allocation; alpine meadow
Corresponding author:Ma ZhenE-mail:qinfenma@126.com
中图分类号:S512.606;Q945.79
文献标识码:A
文章编号:1001-0629(2016)2-0249-10
通信作者:马真(1982-),女,新疆石河子人,助理研究员,博士,主要从事草地恢复研究。E-mail:qinfenma@126.com
基金项目:青海省海南州科技合作项目(2015-HZ-01);国家自然科学基金项目(31172247、31201836、31472135);青海省自然科学基金青年项目(2015-ZJ-919Q、2015-ZJ-918Q、2015-ZJ-711);中国博士后科学基金会(2015M582736);青海省自然科学基金项目(2013-Z-916);国家科技支撑课题专题(2014BAC05B03);青海省重点实验室发展专项资金计划(2014-Z-Y01)
*收稿日期:2015-06-25接受日期:2015-09-09
DOI:10.11829/j.issn.1001-0629.2015-0361
赵宏魁,马真,张春辉,雷占兰,姚步青,周华坤.种植密度和施氮水平对燕麦生物量分配的影响.草业科学,2016,33(2):249-258.
Zhao H K,Ma Z,Zhang C H,Lei Z L,Yao B Q,Zhou H K.The reproductive allocation ofAvenasativaunder different planting densities and nitrogen addition treatments.Pratacultural Science,2016,33(2):249-258.
第一作者:赵宏魁(1964-),男,青海大通人,高级工程师,本科,主要从事草地恢复及保护方面的工作。 E-mail:1156382012@qq.com
