干旱胁迫对3种园林绿化树种幼苗生理指标的影响
2016-04-17列志旸黄威龙许建新
李 洁 列志旸 薛 立 黄威龙 许建新,2
(1.华南农业大学林学与风景园林学院,广东广州510642;2.深圳市铁汉生态环境股份有限公司,广东深圳518040)
干旱胁迫对3种园林绿化树种幼苗生理指标的影响
李 洁1列志旸1薛 立1黄威龙1许建新1,2
(1.华南农业大学林学与风景园林学院,广东广州510642;2.深圳市铁汉生态环境股份有限公司,广东深圳518040)
以小叶榕、海南蒲桃、红桂木3种公路绿化树种幼苗为试材,人工模拟干旱胁迫环境,通过测定各树种幼苗叶片的生理指标,探讨干旱胁迫对3种幼苗生长的影响。研究表明:叶片组织含水量呈下降趋势,而可溶性糖含量呈上升趋势;小叶榕和红桂木幼苗叶片叶绿素含量先升后降,海南蒲桃逐渐上升;小叶榕和红桂木幼苗的蛋白质含量显著增加后保持稳定,海南蒲桃幼苗先升后降;小叶榕和红桂木幼苗的脯氨酸含量上升,海南蒲桃缓慢下降;小叶榕幼苗叶片的SOD活性先降后升,海南蒲桃幼苗逐渐上升,红桂木幼苗先升后降;小叶榕幼苗的MDA含量稳定,海南蒲桃和红桂木幼苗上升。主成分分析结果显示,3种幼苗的抗旱性为红桂木>海南蒲桃>小叶榕。
小叶榕;海南蒲桃;红桂木;幼苗;干旱胁迫;生理特性;主成分分析
随着我国绿化建设的发展,城镇绿化树种的栽培面积迅速扩大。由于城市建设过程中大量的原有土表被破坏,取而代之的新土有机质含量低,土壤结构差,水分蒸发快,随之引起的土壤干旱影响植物生长和生态恢复[1],也影响植物的生理代谢过程[2],因而可以通过生理指标来研究树木的抗旱性[3]。关于植物的抗旱生理研究已有较多报道,如干旱胁迫时,细胞内可溶性糖含量提高,蛋白质含量增加,脯氨酸含量呈上升趋势[3-4],细胞渗透能力增强以抵御干旱的影响[5];过氧岐化酶(SOD)上升,以减少脂质过氧化,稳定膜的透性[6],丙二醛(MDA)随干旱胁迫时间上升,表明细胞膜可能受损[7]。
海南蒲桃(Syzygium hainanense)、小叶榕(Ficus microcarpa)、红桂木(Artocarpus nitidus ssp.lingnanensis)是热带、亚热带地区重要的园林绿化树种。本试验将这3种幼苗置于温室中,模拟干旱环境,测定其生理生化指标,并利用主成分分析法对其抗旱性进行综合评价,得出了3种幼苗抗旱能力的排序,对于筛选择抗旱性强的绿化树种有一定的参考价值。
1 材料与方法
1.1 试验材料
试验地在华南农业大学林学院院楼温室内。处理所用幼苗为1年生实生苗,原装营养袋直径9 cm,深9 cm,后转入直径16 cm,深17 cm的白色无纺布营养袋,基质用荷兰土与黄心土,体积比为1∶1。转入新营养袋培养2周后开始试验,幼苗生长基本情况见表1。

表1 3种幼苗的基本情况Tab.1 Basic information of the three seedlings
1.2 试验方法
将试验苗木置于温室内,采用自然干旱胁迫方法,对待测苗木进行干旱处理。分别于停水当天,4、8、12 d及复水后4 d测定幼苗的生理指标。在测定当日8:30每种幼苗选择5株生长良好的个体,采取其第3至第8片大小均一的功能叶,用自来水冲洗除去表面污物,再用蒸馏水冲洗2~3次后,用吸水纸吸干叶片表面水分。将从5株采集的叶片混合后测定生理指标,每个指标作3次重复测定。
1.3 生理指标测定
组织含水量用烘干称重法测定[8]。叶绿素含量用分光光度法测定、脯氨酸含量用酸性茚三酮法测定、超氧化物歧化酶(SOD)活性用氯化硝基四氮唑蓝(NBT)光化还原法、丙二醛(MDA)含量用硫代巴比妥酸显色(TBA)法[9]。可溶性糖含量参照蒽酮比色法测定[8]。可溶性蛋白质含量参照考马斯亮蓝法[10]。
1.4 数据分析
数据统计分析和作图由Microsoft Excel和IBM SPSS Statistics 19.0软件系统[11]完成,用Duncan多重比较检验同一幼苗在不同处理时间后的差异显著性(显著性水平为0.05)。用主成分分析计算各个抗旱指标的得分,按得分多少评定各树种的抗旱能力[11-12]。进行主成分分析前,按极差法对各指标数据进行标准化:

式中:X为标准化后所得数据;xi为某项指标的原始数据;xmax为某项指标的最大值;xmin为某项指标的最小值。对标准化后的数据进行主成分分析。以累积贡献率达85%以上为参考值,确定主成分个数。因主成分之间相互独立且有着不同的贡献率,所以每个主成分得分是其加权值,其中权重是主成分对应的贡献率,由主成分的特征向量计算出每种幼苗的主成分得分。
2 结果与分析
2.1 叶片含水量
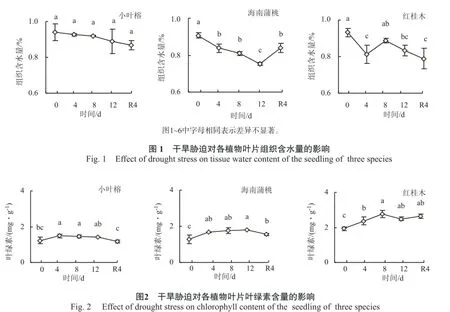
在干旱条件下,抗旱力强的植物可维持较多的水分,组织含水量比较高[13]。干旱胁迫处理期间,小叶榕幼苗的叶片组织含水量缓慢下降,与对照差异不显著;海南蒲桃幼苗的叶片组织含水量逐渐下降,干旱胁迫12 d时显著小于对照(P<0.05);红桂木幼苗叶片的组织含水量呈现波动(图1)。
2.2 叶绿素含量
叶绿素作为集光色素可以捕获光能用于光合生产,其含量的高低表明植物光合能力的强弱[14]。干旱胁迫处理期间,小叶榕和红桂木幼苗叶片叶绿素含量先升后降,复水处理4 d后,红桂木幼苗的叶片的叶绿素含量显著大于对照(P<0.05)(图2);海南蒲桃的叶绿素含量逐渐上升,干旱胁迫4 d、8 d和12 d时显著大于对照(P<0.05)。复水处理4 d后,红桂木和海南蒲桃幼苗的叶片叶绿素含量显著大于对照(P<0.05)。
2.3 可溶性糖含量
可溶性糖积累的越多,植物的渗透调节能力就越强。植物吸水和保水能力强,增强了抗旱性[7,15-16]。干旱胁迫处理期间,小叶榕和红桂木幼苗的叶片可溶性糖含量逐渐上升,并在第12天的时候,达到最大值(图3)。海南蒲桃干旱胁迫处理4 d和8 d时与对照无显著差异,12 d时显著大于对照(P<0.05)。复水处理4 d后,3种幼苗的叶片可溶性糖含量都下降到对照水平。

2.4 蛋白质含量
植物体内的可溶性蛋白大多是参与各种代谢的酶类,在受到干旱胁迫时,植物体内会产生大量的可溶性蛋白,高含量的可溶性蛋白可帮助植物细胞维持较低的渗透势,保持水分平衡,抵抗干旱胁迫带来的伤害[17]。干旱胁迫处理期间,小叶榕和红桂木幼苗的叶片蛋白质含量显著增加后保持稳定,复水处理4 d后显著大于对照(P<0.05)(图4)。海南蒲桃幼苗的叶片蛋白质含量先升后降,4 d时达到最大水平,然后缓慢下降,复水处理4 d后与对照无显著差异。
2.5 脯氨酸含量
脯氨酸是一种具有较强水合能力的渗透调节物质,在植物遭受抗水分胁迫时发挥作用,既可以帮助细胞维持相对稳定的水环境[18-19],同时又可以消除氨积累对植物体所造成的毒害[8,20]。干旱胁迫处理期间,小叶榕幼苗的脯氨酸含量显著上升后缓慢增加,海南蒲桃缓慢下降,红桂木幼苗显著上升(图5)。经过复水处理4 d后,3种幼苗的叶片脯氨酸含量均下降到对照水平(P<0.05)。

2.6 超氧化物歧化酶活性
植物体有清除活性氧的酶系统和非酶促清除剂,可防御活性氧对细胞的伤害[21]。干旱胁迫期间,小叶榕幼苗叶片的SOD活性先降后升,12 d和复水处理4 d时与对照无显著差异(图6)。海南蒲桃幼苗叶片的SOD活性逐渐上升,12 d时显著大于对照(P<0.05),经过4 d复水处理后,下降到对照水平。干旱胁迫处理期间,红桂木幼苗的SOD活性先升后降,复水处理4 d后下降到对照水平。

2.7 丙二醛含量
植物在逆境下遭受伤害,往往发生膜脂过氧化作用,丙二醛(MDA)是膜脂过氧化的最终产物,它的积累会对植物细胞原生质膜造成不同程度的破坏,其含量的变化反映细胞膜损伤的程度和植物抗旱能力的大小[22]。干旱胁迫期间,小叶榕幼苗叶片MDA含量与对照无显著差异,海南蒲桃幼苗缓慢的上升后急剧上升,红桂木幼苗显著上升,后二者12 d时显著大于对照(P<0.05)(图7)。4 d复水处理后,3种幼苗的MDA含量与对照无显著差异。
2.8 3种幼苗抗旱性的综合评价
对幼苗的生理指标进行主成分分析,由主成分的特征向量计算出3种幼苗的主成分得分顺序为红桂木、海南蒲桃、小叶榕[11-12](表2)。因此,在热带、亚热带地区,3种重要的园林绿化树种幼苗抗旱性大小由大到小的顺序为红桂木、小叶榕、海南蒲桃。


表2 干旱胁迫下3种幼苗的生理生化指标得分Tab.2 Drought resistance scores of the three seedlings under drought stress
3 结论与讨论
1)叶片含水量和叶绿素。干旱胁迫处理期间,3种幼苗的叶片含水量均下降,其中小叶榕幼苗与对照差异不显著,说明其在干旱胁迫下可以维持自身含水量的稳定,抗旱性强。经干旱胁迫处理后,3种幼苗叶片的叶绿素含量均有不同程度的增加,原因可能是苗木在干旱胁迫处理后,体内水分降低,叶片变轻,每克叶干质量、细胞数增加,叶绿素呈相对浓缩状态,所以单位叶片鲜质量中叶绿素含量就会相对升高[3,7]。
2)渗透调节物质。在干旱胁迫期间,海南蒲桃叶片可溶性糖含量有所增加,但增长幅度小,不利于降低植物水势,表明其抗旱性较弱。小叶榕和红桂木幼苗的叶片可溶性糖含量逐渐上升,并在第12天时达到最大值,有利于抗旱。经过复水处理4 d后,3种幼苗的叶片可溶性糖含量均下降到对照水平,说明复水后土壤干旱胁迫得以缓解,3种植物体内调节物质逐渐向正常水平逆转,渗透调节能力逐步下降,引起可溶性糖含量减少,这与孙宗玖等[23]的研究结果一致。干旱胁迫处理期间,小叶榕和红桂木的幼苗叶片蛋白质显著增加后保持较高水平,有利于维持植物细胞较低的渗透势,保持水分平衡,抵抗干旱胁迫带来的伤害,具有较强的抵抗干旱胁迫的能力。海南蒲桃先增加后略降,可能是在胁迫处理初期,海南蒲桃幼苗通过增加可溶性蛋白含量来增强渗透调节能力,维持较低的渗透势,以抵抗水分胁迫带来的伤害[24]。而在胁迫处理后期,干旱胁迫超过植物所能耐受的阈值,胁迫使RNA转录和翻译受到抑制,造成蛋白质的合成量减少[25],或者加速了其降解。本研究中,小叶榕和红桂木幼苗的脯氨酸含量显著大于对照水平,有利于抵御干旱。
3)SOD活性和MDA含量。干旱胁迫处理期间,海南蒲桃幼苗叶片SOD活性逐渐上升,SOD保护酶增强了植物抗氧化能力,避免了活性氧等各种自由基的大量积累,所以抗旱性强[8,20]。红桂木幼苗叶片的SOD活性先升后降,与李云飞等[26]对干旱胁迫下紫叶矮樱(Prunus×cistenena)和刘娟等[21]对白羊草(Bothriochloa ischaemum)的研究结果相似。干旱胁迫初期,SOD保护酶增强避免了活性氧等各种自由基的大量积累。随着干旱胁迫过程的加剧,植物体内保护酶系统的活力和平衡受到破坏,酶降解的增加和合成的减慢可能是导致SOD活性下降原因之一[21]。小叶榕幼苗叶片的SOD活性先降后升,与蒋明义等[27]对水稻(Oryza sativa)幼苗的研究结果相似。可能是水分胁迫造成的膜伤害包含着一个自由基伤害过程,抗旱物种在严重水分胁迫时仍有较活跃的SOD合成调节。SOD只是自由基清除酶庞大系统中的一员,细胞内的其他非酶系统也具有清除自由基的作用。干旱胁迫处理期间,海南蒲桃和红桂木幼苗的叶片丙二醛含量上升,12 d时2种幼苗的叶片丙二醛含量均显著大于对照。小叶榕幼苗的叶片丙二醛含量保持稳定,表明其可能有较强的防止膜脂过氧化作用的能力,因而具有较强的抗旱性。
4)幼苗抗旱性综合评价。植物的抗旱性是多种指标综合作用的结果[28-29]。对树木抗旱性的评价,当涉及到多种植物、多个指标的情况时,因各指标所表达的树木抗旱性顺序往往不一致,利用一般的简单方法很难得出确切结果。本研究运用主成分分析对3个树种的生理指标进行了综合评定,根据加权后的主成分得分可知,其抗旱性由大到小的顺序为红桂木、海南蒲桃、小叶榕。
[1] 赵明坤,刘秀峰,唐成斌.高等级公路边坡绿化草被植物筛选及其生物学特性[J].贵州农业科学,2004,32(2):18-21.
[2] 任磊,赵夏陆,许靖,等.4种茶菊对干旱胁迫的形态和生理响应[J].生态学报,2015,16(15):5-10.
[3] 刘琴,孙辉,何道文.干旱和高温对植物胁迫效应的研究进展[J].西华师范大学学报(自然科学版),2005,18(4):364-368.
[4] 史小玲,薛立,任向荣,等.华南地区4种阔叶幼苗水分胁迫条件下的抗旱性初探[J].林业科学研究,2011,13(6):760-767.
[5] 谢小玉,马仲炼,白鹏,等.辣椒开花结果期对干旱胁迫的形态与生理响应[J].生态学报,2014,34(13):3797-3805.
[6] 邵怡若,许建新,薛立,等.5种绿化树种幼苗对干旱胁迫和复水的生理响应[J].生态科学,2013,32(4):420-428.
[7] 冯慧芳,薛立,任向荣,等.4种阔叶幼苗对PEG模拟干旱的生理响应[J].生态学报,2011,31(2):371-382.
[8] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版,2000.
[9] 陈建勋,王晓峰.植物生理学实验指导[M].广州:华南理工大学出版社,2002.
[10] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[11] 邵怡若,许建新,薛立,等.低温胁迫时间对4种幼苗生理生化及光合特性的影响[J].生态学报,2013,33(14):4237-4247.
[12] 卢广超,许建新,薛立,等.干旱胁迫下4种常用植物幼苗的光合和荧光特性综合评价[J].生态学报,2013,33(24):7872-7881.
[13] 董明,苏德荣,刘泽良,等.干旱胁迫对阿诺红鞑靼忍冬生理指标的影响[J].西北林学院学报,2008,23(4):8-13.
[14] 金不换.干旱胁迫对不同品种早熟禾形态和生理特性影响的研究[D].哈尔滨:东北农业大学,2009.
[15] 刘芳,左照江,许改平,等.迷迭香对干旱胁迫的生理响应及其诱导挥发性有机化合物的释放[J].植物生态学报,2013,38(5):454-463.
[16] 张诚诚,曹志华,胡娟娟,等.水分胁迫对油茶容器苗生理生化特性的影响[J].安徽农业大学学报,2013,40(4):623-626.
[17] 宫丽丹,田耀华,龙云峰,等.持续干旱胁迫及复水对橡胶树渗透调节能力的影响[J].中国农学通报,2012,24(1):35-38.
[18] 刘涛,李柱,安沙舟,等.干旱胁迫对木地肤幼苗生理生化特性的影响[J].干旱区研究,2008,23(2):231-235.
[19] 李力,刘立强,周光益,等.南岭常绿阔叶林林冠受损对穿透雨和树干流水化学的影响[J].水土保持学报,2014,19(2):45-50.
[20] 李燕,孙明高,孔艳菊,等.皂角苗木对干旱胁迫的生理生化反应[J].华南农业大学学报,2006,24(3):66-69.
[21] 刘娟,董宽虎.干旱胁迫及复水处理对白羊草抗旱生理特性的影响[J].草原与草坪,2011,34(2):74-78.
[22] 谷艳蓉,张国芳,孟林.4种牧草幼苗对水分胁迫的响应及其抗旱性[J].四川草原,2005,15(4):4-7.
[23] 孙宗玖,李培英,阿不来提,等.干旱复水后4份偃麦草渗透调节物质的响应[J].草业学报,2009,18(5):52-57.
[24] 白向历,齐华,刘明,等.玉米抗旱性与生理生化指标关系的研究[J].玉米科学,2007,23(5):79-83.
[25] 史玉炜,王燕凌,李文兵,等.水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响[J].新疆农业大学学报,2007,30(2):5-8.
[26] 李云飞,李彦慧,王中华,等.土壤干旱胁迫及复水对紫叶矮樱生理特性的影响[J].河北农业大学学报,2008,22(6):78-82.
[27] 蒋明义,荆家海,王韶唐.渗透胁迫对水稻幼苗膜脂过氧化及体内保护系统的影响[J].植物生理学报,1991,22(1):80-84.
[28] 季孔庶,孙志勇,方彦.林木抗旱性研究进展[J].南京林业大学学报(自然科学版),2006,30(6):123-128.
[29] 张朋飞,武军艳,孙万仓,等.干旱胁迫对白菜型冬油菜苗期生理特性的影响[J].西北农业学报,2015,13(2):84-90.
(责任编辑 张 坤)
Physiological Changes of Three Greening Seedling Species under Drought Stress
Li Jie1,Lie Zhiyang1,Xue Li1,Huang Weilong1,Xu Jianxin1,2
(1.College of Forestry and Landscape Architecture,South China Agricultural University,Guangzhou Guangdong 510642,China;2.Shenzhen Techand Ecology&Environment Co.Ltd.,Shenzhen Guangdong 518040,China)
Studies were conducted on seedlings of Ficusmicrocarpa,Syzygium hainanense and Artocarpus nitidus ssp.lingnanensis under simulated droughtenvironment to study their physiological changes and then explore the effects of stress on the growth of seedlings.The result showed that,under drought stress,tissue water contents of the three seedlings tended to decreased,whereas thesoluble sugar content tended to increased;chlorophyll content of F.microcarpa and A.nitidus ssp.lingnanensis increased first and decreased afterwards and S.hainanense increased gradually;protein contentof F.microcarpa and A.nitidus ssp.lingnanensis increased and then keptsteady,and S.hainanense increased first and decreased afterwards;the contentof free proline of F.microcarpa and A. nitidus ssp.lingnanensis increased,whereas S.hainanense decreased slowly;the activity of SOD of F.microcarpa first decreased and increased afterwards,S.hainanense increased gradually,and A.nitidus ssp.lingnanensis increased first and decreased afterwards.The contents of MDA of F.microcarpa kept steady,whereas S.hainanense and A.nitidus ssp.lingnanensis increaseds.Principal component analysis indicated that drought resistance of the three seedling types was in the order of A.nitidus ssp.lingnanensis>S.hainanense>F.microcarpa.
Ficusmicrocarpa;Syzygium hainanense;Artocarpusnitidus ssp.lingnanensis;seedling;drought stress;physiological characteristics;principal component analysis

S718.43
A
2095-1914(2016)02-0056-06
10.11929/j.issn.2095-1914.2016.02.009
2015-08-15
深圳市战略性新兴产业专项资金项目(GCZX20120618100801416)资助。
第1作者:李洁(1992—),女,硕士生。研究方向:风景园林和植物生理。Email:houis_56028923@qq.com。
薛立(1958—),男,博士,教授。研究方向:森林培育和生态学。Email:forxue@scau.edu.cn。
