丙酮醛降解酶基因的克隆与高效表达
2016-04-11王平赵巧巧黄鹰
王平++赵巧巧 +黄鹰
摘要:由于丙酮醛降解酶(methylglyoxal degradation enzyme,MD)是生物体内代谢丙酮醛的一种关键酶,实现MD的高效表达对调控丙酮醛的代谢途径具有重要意义。所以根据Schizosaccharomyces pombe_GeneDB中MD的基因序列(SPAC22E12.03C)设计引物,以粟酒裂殖酵母(Schizosaccharomyces pombe )cDNA文库为模板通过PCR扩增技术得到目标基因,目的片段全长576 bp。将SPAC22E12.03C连接到pET-28a(+)上,得到重组质粒pET-28a(+)-SPAC22E12.03C,并在Escherichia coli BL2l(DE3)中实现表达。同时,对其目的蛋白的表达条件进行了优化,获得最佳表达条件:诱导温度37 ℃,诱导起始菌体D600 nm为0.5~0.6,诱导剂IPTG浓度为0.8 mmol/L。这为研究粟酒裂殖酵母体内丙酮醛的降解途径打下了坚实基础。
关键词:粟酒裂殖酵母;丙酮醛降解酶;丙酮醛;基因克隆;基因表达
中图分类号: Q785文献标志码: A
文章编号:1002-1302(2016)02-0062-04
[HJ1.4mm]
收稿日期:2015-03-24
基金项目:国家自然科学基金(编号:31070703) 。
作者简介:王平(1990—),女,河南南阳人,硕士研究生,从事粟酒裂殖酵母分子机理研究。E-mail:350479708@qq.com。
通信作者:黄鹰,教授,从事微生物生物化学与分子生物学研究。E-mail:paula_wong@163.com。[HJ]
丙酮醛(methylglyoxal)是在糖酵解、脂质和氨基酸代谢过程中产生的一种对生物体有毒害作用的代谢物[1-2],性质活跃,属于α-酮基醛类,由糖酵解的中间产物丙糖磷酸化裂解生成,或由丙酮和氨基丙酮代谢生成[3]。其醛基可与蛋白质的氨基基团之间发生非酶性糖基化反应(又称Maillard反应)[4]形成一系列具有高度活性和高度异质性的终产物,从而导致蛋白功能的修改和缺陷,Ward等研究证实,丙酮醛能通过细胞内颗粒释放而加强应激反应[5],并进一步诱导细胞的应激反应,包括胞吞、细胞因子释放和细胞凋亡[6]等,丙酮醛在生物体内过度累积会导致细胞死亡;此外,丙酮醛与生物体的很多病变有关,如慢性糖尿病[7-8]、结肠癌和乳腺癌等。因此,丙酮醛在生物体内的累积量必须受到严格的控制,丙酮醛降解酶是代谢丙酮醛的一种关键酶,于是对丙酮醛降解酶的研究显得尤为重要。
近年来,各种慢性病患病率的增加,使得丙酮醛在体内的代谢途径受到越来越多的关注,丙酮醛代谢相关酶的研究也成为国际热点。研究报道在细菌体内丙酮醛的降解主要通过3条途径:(1)依赖于谷胱甘肽的醛酮变位酶Ⅰ和醛酮变位酶Ⅱ[9];(2)不依赖于谷胱甘肽的醛酮变位酶Ⅲ;(3)依赖于NADPH的醛还原酶和依赖于NADH的乙醇脱氢酶[10]。人体内的DJ-1也具有分解丙酮醛的作用,我们以粟酒裂殖酵母(Schizosaccharomyces pombe)为模式生物,通过构建系统进化树和基因比对,找到了粟酒裂殖酵母体内的能够降解丙酮醛的酶,即SPAC22E12.03C的产物。
本试验根据SPAC22E12.03C的基因序列设计引物,以粟酒裂殖酵母的cDNA为模板通过PCR扩增技术得到了目标酶的基因,构建T7强启动子表达载体和重组工程菌株,研究丙酮醛降解酶的高效表达,希望以后能够通过高效表达丙酮醛降解酶来调控和优化丙酮醛的代谢途径,从而为解决因丙酮醛在体内累积过多引起的各种慢性疾病提供研究基础。
1材料与方法
1.1菌株与质粒
本试验中所用的菌株名为Escherichia coli BL2l (DE3)、Escherichia coli top10,所用的质粒为pET-28a(+),这些材料均为笔者所在实验室保存。粟酒裂殖酵母cDNA文库来自美国,重组克隆pET-28a(+)-SPAC22E12.03C为笔者所在实验室构建。
1.2试剂及其来源
限制性内切酶NcoⅠ和XhoⅠ、solutionⅠ[kite code.6022]、rTaq DNA 聚合酶、PrimeSTAR DNA聚合酶、蛋白marker、异丙基硫代-β-D-半乳糖苷(IPTG),均购自TaKaRa 公司;DNA 割胶回收试剂盒,来源于BioSpin Gel Extraction Kit;PCR过柱纯化,来源于BioFLUX BioSpin PCR Purification Kit试剂盒;DNA marker 来源于北京全式金生物技术有限公司;PCR引物由南京思普金生物科技有限公司合成;卡那霉素(Kan)来源于Sigma 公司,其他常规试剂均为国产分析纯。
1.3粟酒裂殖酵母中目的片段SPAC22E12.03C的获得
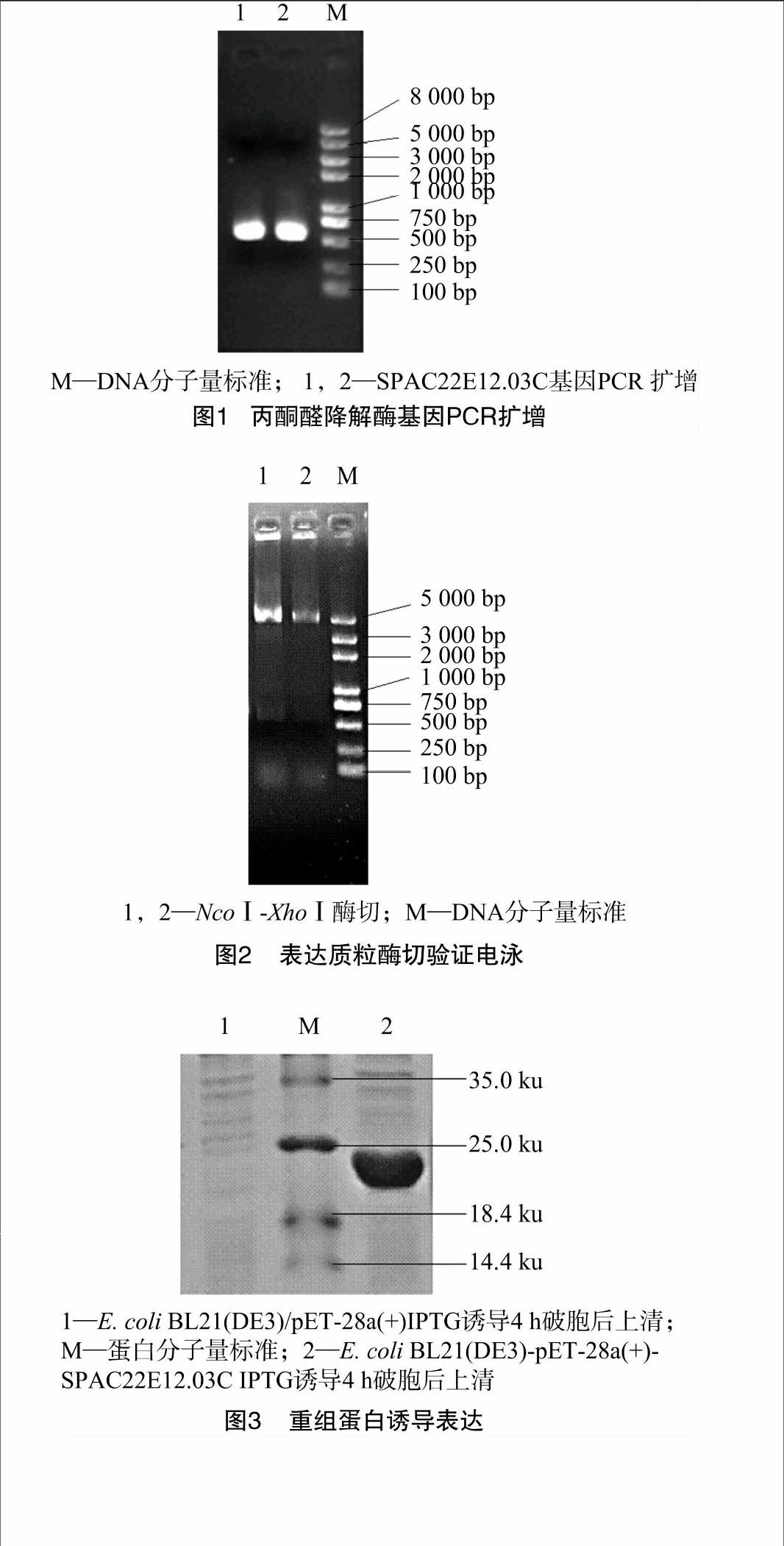
根据S.pombe_GeneDB数据库中的丙酮醛降解酶基因(SPAC22E12.03C)序列设计上、下游引物,分别为SPAC22E12.03C-NcoⅠ-up:5′-CATG[ZZ(Z]CCATGG[ZZ)]TAAAGGTTTGCCTATTTGTTG-3′,SPAC22E12.03C-XhoⅠ-down:5′-CCG[ZZ(Z]CTCGAG[ZZ)]GGGCATACTAAGAGATTTATAGACA-3′ (下划线分别为NcoⅠ和XhoⅠ酶切识别位点)。以粟酒裂殖酵母cDNA为模板扩增丙酮醛降解酶基因,PCR反应体系为25 μL:ddH2O 14.7 μL、5×PS buffer 5 μL、dNTP(2.5 mmol/L) 2 μL、cDNA模板1 μL、SPAC22E12.03C-NcoⅠ-up 1 μL、SPAC22E12.03C-XhoⅠ-down 1 μL、高保真DNA聚合酶 03 μL。[JP3]PCR反应条件为:95 ℃预变性 2 min;95 ℃ 变性 30 s,50 ℃退火 30 s,72 ℃ 延伸1 min,共30个循环;最后 72 ℃ 延伸10 min。用1%的琼脂糖凝胶电泳检测PCR扩增结果。
1.4重组表达质粒的构建和鉴定
将上述PCR扩增片段SPAC22E12.03C过柱纯化后与质粒pET-28a (+)双酶切验证。酶切位点为NcoⅠ和XhoⅠ。将酶切产物回收得到目的DNA片段与质粒pET-28a (+)酶切产物。接下来建立酶连体系为16.6 μL:SPAC22E12.03C目的片段8 μL、pET-28a (+)酶切产物0.3 μL、连接酶(SolutionⅠ) 8.3 μL。16 ℃ 连接4 h,将酶连产物导入到E. coli Top10感受态细胞中,涂布到含Kan抗性的LB培养基上。随机挑取转化子提其质粒进行酶切验证,将阳性质粒送到南京斯普金科技有限公司测序。将测序正确的重组质粒和pET-28a(+)空载质粒分别通过转化导入E. coli BL21(DE3)感受态细胞,得到重组表达菌和带有空载质粒的对照菌。
1.5目的蛋白的表达
将得到的重组表达菌和带有空载质粒的对照菌分别接种于5 mL LB培养基中,其中含有50 mg/L Kan。37 ℃ 220 r/min 恒温摇床过夜培养之后将重组表达菌和对照菌分别转接至15 mL 同样含50 mg/L Kan的LB培养基中,此时菌液D600 nm≈0.2,继续培养,当D600 nm达到0.6时,加入1 mmol/L IPTG,37 ℃ 220 r/min 培养过夜,次日,4 ℃ 8 000 r/min离心5 min,收集菌体,用100 mmol/L 磷酸钠缓冲液(pH值为7.0)重悬菌体进行洗涤,重复洗涤3遍后加入适量的缓冲液(包含0.1 mmol/L PMSF和14.3 mmol/L的β-巯基乙醇)置于冰浴中进行超声波破碎,破碎10 s,冷却10 s,待菌体破碎完全后,4 ℃ 12 000 r/min 离心20 min,收集细胞上清,取适量进行SDS-PAGE,分析目标产物表达情况。
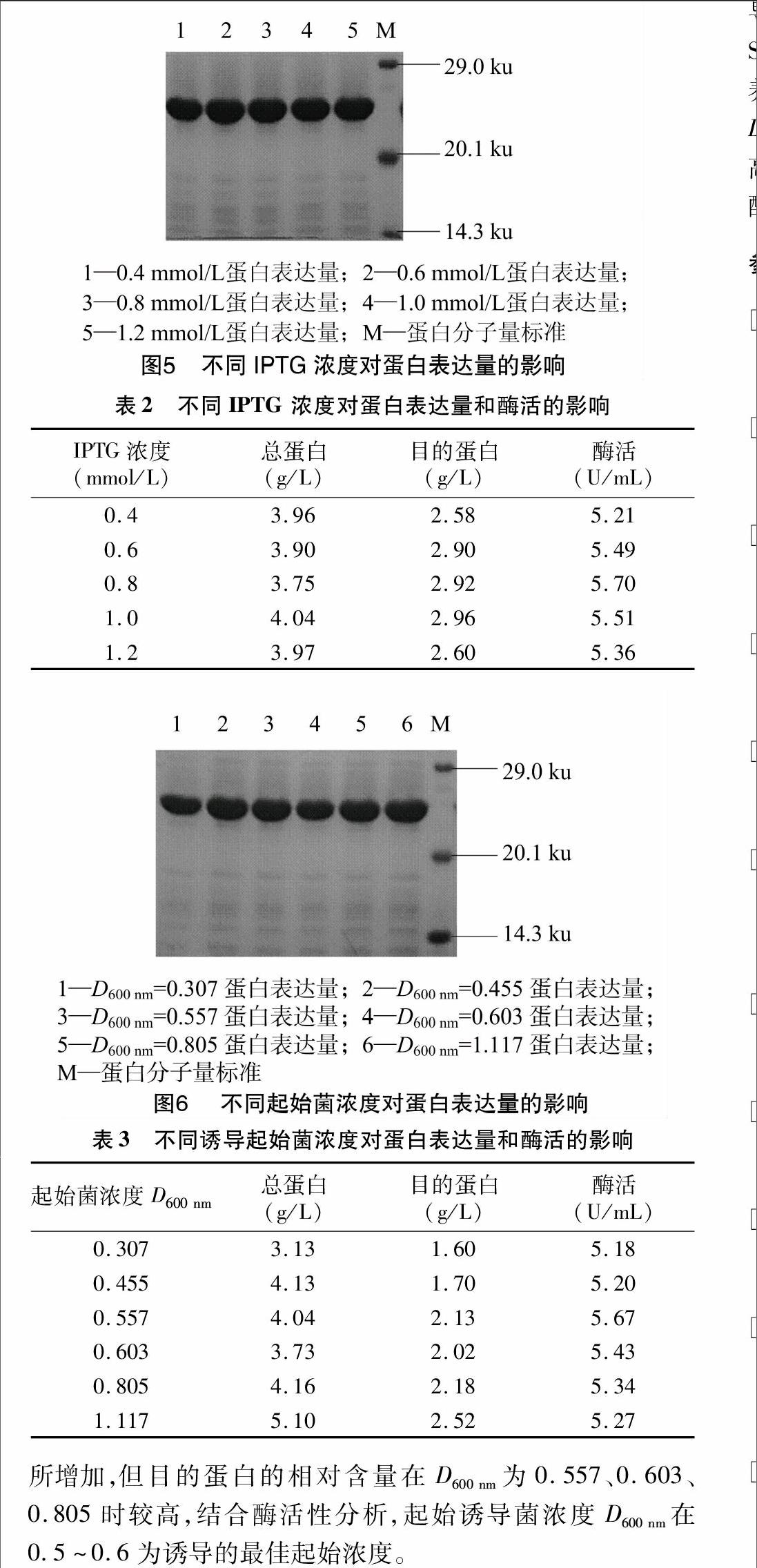
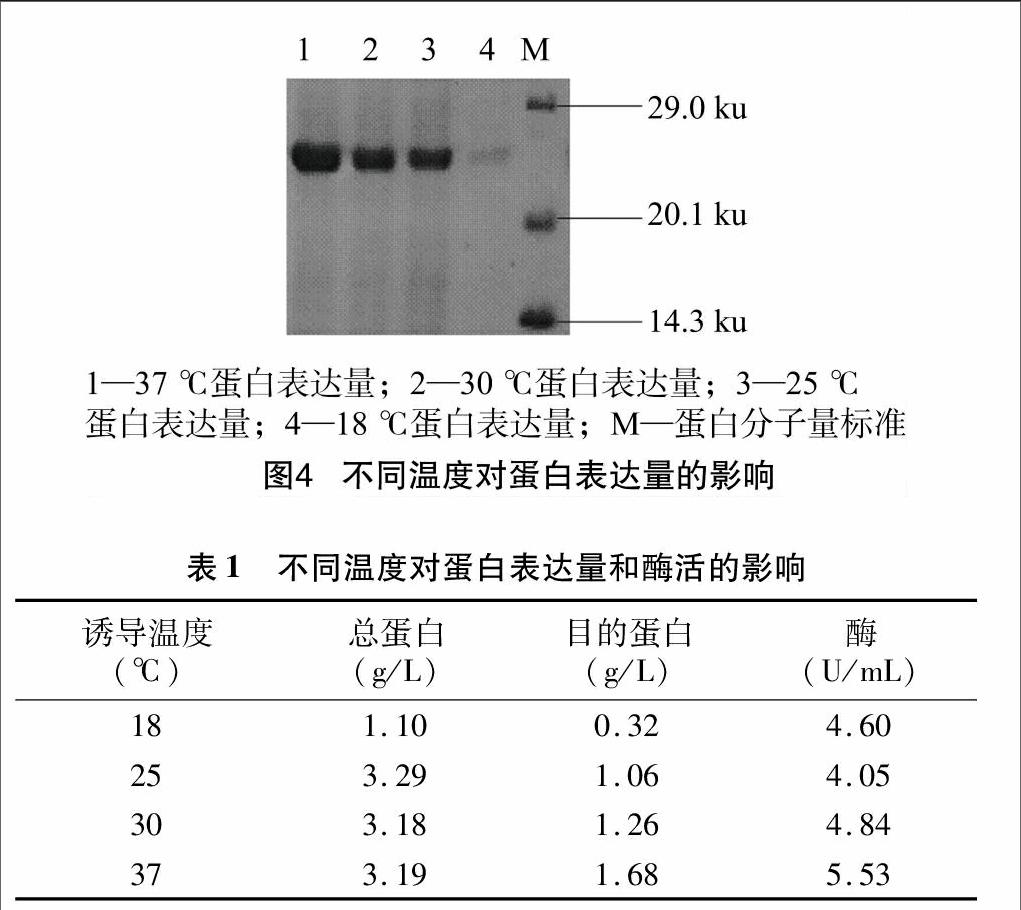
1.6目标蛋白表达条件的优化
优化表达条件时分别选择不同起始诱导菌浓度(D600 nm=0.307,D600 nm=0.455,D600 nm=0.557,D600 nm=0.603,D600 nm=0.805,D600 nm=1.117)与不同温度(37、30、25、18 ℃)培养;培养基中添加不同终浓度的IPTG(0.4、0.6、0.8、1.0、1.2 mmol/L)进行诱导培养,每个试验重复3次。通过SDS-PAGE 、总蛋白含量测定和酶活测定考察培养条件对目的蛋白表达的影响。
1.7酶活测定
酶活测定根据丙酮醛降解酶在一定温度条件下能降解丙酮醛,而丙酮醛可以与2,4-二硝基苯肼反应生成黄橙色的2,4-二硝基苯腙,苯腙在碱性条件下显紫红色,可在540 nm处检测[11]。反应体系是用100 mmol/L 磷酸钠缓冲液(pH 值为7.0)配制成不同丙酮醛浓度的反应液,总体积为200 μL,加入适量的丙酮醛降解酶,在45 ℃反应一定时间,加入2,4-二硝基苯肼终止反应,室温下静置15 min,再加入10%的NaOH,室温放置15 min后用分光光度计在540 nm处测定吸光度。酶活单位(U)定义为在该反应条件下,1 min内催化降解0.1 μmol丙酮醛所需的酶量。
2结果与分析
2.1丙酮醛降解酶基因的克隆
以粟酒裂殖酵母cDNA为模板,以SPAC22E12.03C-NcoⅠ-up、 SPAC22E12.03C-XhoⅠ-down为引物进行PCR,扩增获得1条大于500 bp的DNA产物(图1),与目的基因576 bp相吻合,过柱纯化PCR产物,用NcoⅠ和XhoⅠ对纯化后的PCR产物进行双酶切。
2.2表达载体pET-28a(+)-SPAC22E12.03C的构建及验证
[CM(24]获得丙酮醛降解酶基因并纯化后,将其连接到pET-28a
(+)(NcoⅠ和XhoⅠ双酶切)得到pET-28a(+)-SPAC22E12.03C重组质粒并转化E. coli Top10感受态细胞,挑选菌落,PCR验证有条带的阳性重组子继续培养,并提取重组质粒进行双酶切验证,双酶切后得到大小分别为500 bp和5 400 bp的条带,基因片段大小正确。重组质粒经测序比对与S.pombe_Gene DB上公布的基因序列一致,证明丙酮醛降解酶基因已插入到pET-28a(+)载体中(图2),成功构建了pET-28a(+)-SPAC22E12.03C表达载体。将构建成功的pET-28a(+)-SPAC22E12.03C 转入到E. coli BL21(DE3)感受态细胞中,得到重组菌。
[FK(W14][TPWP2.tif][FK)]
2.3重组菌的诱导表达
重组菌诱导表达经SDS-PAGE验证(图3),得到分子量大小约为20 ku的重组酶,与目的蛋白S.pombe_GeneDB上公布的21.08 ku相符,证明丙酮醛降解酶得到了表达。
[FK(W10][TPWP3.tif][FK)]
2.4丙酮醛降解酶的优化表达
2.4.1温度对丙酮醛降解酶可溶性表达的影响据文献报道,温度对目的蛋白的可溶性表达有很大的影响[12-13]。选定的条件为:在37、30、25、18 ℃,220 r/min 培养至D600 nm为0.8时,加入1 mmol/L IPTG,诱导4 h。由SDS-PAGE(图4和表1)分析可得,丙酮醛降解酶在37 ℃培养后上清中可溶蛋白的含量最多,相对沉淀中目的蛋白含量最少,酶活最高。可见37 ℃不仅是E. coli BL21的最适生长温度,并且在这个温度下有活性的菌和带外源基因的菌更多,从而表达的目的蛋白也多,且大部分以可溶形式存在,酶的活性也最高。
2.4.2IPTG对丙酮醛降解酶可溶性表达的影响在一定范围内,IPTG 浓度对重组蛋白诱导表达有一定影响[14],降低表达水平可能提高某些目的蛋白的产量。因此,本试验选取了5个IPTG浓度,分别为0.4、0.6、0.8、1.0、1.2 mmol/L。在 37 ℃ 220 r/min 培养D600 nm=0.6时,加入上述不同浓度的IPTG,37 ℃诱导4 h,分析IPTG浓度对丙酮醛降解酶可溶性表达的影响。表达载体pET28a(+)-SPAC22E12.03C含有T7/Lac 启动子,受IPTG诱导启动表达蛋白。当IPTG浓度过低时,外源基因不能被完全表达,而高浓度的IPTG将对细胞的生长产生毒害作用,由图5和表2分析可知,IPTG浓度在0.6~1.0 mmol/L 时,总蛋白量略微升高,此范围内酶的活性也相对较高,但就三者目的蛋白的相对含量分析:0.6 mmol/L时相对含量为74.4%,0.8 mmol/L 时目的蛋白相对含量为77.9%,1.0 mmol/L时相对含量为73.3%,由蛋白相对含量和酶活大小分析可得,0.8 mmol/L时蛋白相对表达含量最高,酶的活性最高,因此选择0.8 mmol/L作为诱导剂IPTG的最适浓度。
2.4.3诱导起始菌浓度不同对丙酮醛降解酶可溶性表达的影响据文献报道,诱导起始菌浓度不同,菌体的生长速率不同,外源蛋白表达水平会受到影响[15]。本试验选取了6个不同起始菌浓度:D600 nm=0.307,D600 nm=0.455,D600 nm=0.557,D600 nm=0.603,D600 nm=0.805,D600 nm=1.117,加入0.8 mmol/L的IPTG,37 ℃诱导4 h。由图6和表3分析可知,随着诱导起始菌浓度的增加,总蛋白水平在增加,目的蛋白的表达量也有
3结论
在前期研究基础上,本试验开展了对丙酮醛降解酶基因的克隆及其目的蛋白表达的研究。在对目的基因SPAC22E12.03C进行克隆时,选取的验证酶切位点为NcoⅠ和XhoⅠ。接下来经过酶联与转化,得到重组质粒,经测序,确定其为阳性质粒。再次经过转化,成功得到重组表达菌和带有空载质粒的对照菌。
接下来本试验采取单因素控制变量法对目的蛋白最佳表达条件的进行了探索。试验采用的3个变量分别为温度、诱导剂IPTG、起始诱导菌浓度。检测最佳条件的3个指标为SDS-PAGE、总蛋白含量测定和酶活测定。结果表明,在培养温度为37 ℃,IPTG浓度为0.8 mmol/L,诱导起始菌浓度的D600 nm在0.5~0.6时,结合酶活性分析,目的蛋白相对含量最高。通过对目的蛋白诱导表达条件的优化,为更好地揭示丙酮醛降解酶的作用机理提供了很好的平台。
[HS2][HT8.5H]参考文献:[HT8.SS]
[1][ZK(#]Thornalley P J,Langborg A,Minhas H S. Formation of glyoxal,methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose[J]. Biochemical Journal,1999,344(1):109-116.
[2]刘学伟,杨克,张杰. 丙酮醛对人牙周膜成纤维细胞的毒性作用[J]. 云南大学学报:自然科学版,2006,28(增刊1):393-395.
[3]Murata-Kamiya N,Methylglyoxal K H,Aldehyde A E. Crosslinks DNA polymerase and the substrate DNA[J]. Nucleic Acids Research,2001,29(16):3433-3438.
[4]Ferguson G P,Ttemeyer S,Maclean M J,et al. Methylglyoxal production in bacteria:suicide or survival?[J]. Archives of Microbiology,1998,170(4):209-218.
[5]Ward R A,Mcleish K R. Methylglyoxal:a stimulus to neutrophil Oxygen radical production in chronic renal failure?[J]. Nephrology Dialysis Transplantation,2004,19(7):1702-1707.
[6]Okado A,Kawasaki Y,Hasuike Y,et al. Induction of apoptotic cell death by methylglyoxal and 3-deoxyglucosone in macrophage-derived cell lines[J]. Biochemical and Biophysical Research Communications,1996,225(1):219-224.
[7]Mclellan A C,Thornalley P J,Benn J,et al. Glyoxalase system in clinical diabetes mellitus and correlation with diabetic complications[J]. Clinical Science,1994,87(1):21-29.
[8]Ahmed N,Babaei-Jadidi R,Howell S K,et al. Degradation products of proteins damaged by glycation,oxidation and nitration in clinical type 1 diabetes[J]. Diabetologia,2005,48(8):1590-1603.
[9]Thornalley P J. The glyoxalase system:new developments towards functional characterization of a metabolic pathway fundamental to biological Life[J]. Biochemical Journal,1990,269(1):1-11.
[10][ZK(#]van der Jagt D L,Robinson B,Taylor K K,et al. Reduction of trioses by NADPH-dependent aldo-keto reductases. Aldose reductase,methylglyoxal,and diabetic complications[J]. The Journal of Biological Chemistry,1992,267(7):4364-4369.
[11]Lee J Y,Song J,Kwon K,et al. Human DJ-1 and its homologs are novel glyoxalases[J]. Human Molecular Genetics,2012,21(14):3215-3225.
[12]Sorensen H P,Mortensen K K. Soluble expression of recombinant proteins in the cytoplasm of Escherichia coli[J].Microbial Cell Factories,2005,4(1):1.
[13]Williams R E,Bruce N C. ‘New uses for an old enzyme—the old yellow enzyme family of flavoenzymes[J]. Microbiology,2002,148(Pt 6):1607-1614.
[14]冯志国,刘慧娟,陆婕,等. 一新富含甘氨酸果蝇抗菌肽在大肠杆菌中的优化表达[J]. 生物技术,2008,18(4):18-20.
[15]Xu Z,Peng L,Zhong Z,et al. High-level expression of a soluble functional antimicrobial peptide,human beta-defensin 2,in Escherichia coli[J]. Biotechnology Progress,2006,22(2):382-386.
