洋葱T型细胞质雄性不育与花蕾ATP含量的关系
2016-04-06马蓉丽成妍吴海涛焦彦生乔
马蓉丽成 妍吴海涛焦彦生乔 宁
(1山西省农业科学院蔬菜研究所,山西太原 030031;2温州科技职业技术学院,浙江温州325000)
洋葱T型细胞质雄性不育与花蕾ATP含量的关系
马蓉丽1成 妍1吴海涛1,2*焦彦生1乔 宁1
(1山西省农业科学院蔬菜研究所,山西太原 030031;2温州科技职业技术学院,浙江温州325000)
摘 要:以洋葱T型细胞质雄性不育系8-43A、保持系8-43B和恢复系8-43-57C为试材,采用萤火虫荧光素酶检测技术,分析花蕾不同发育时期三磷酸腺苷(ATP)的含量变化,研究洋葱T型细胞质雄性不育与花蕾ATP含量的关系。结果表明:在整个花药发育过程中的各个阶段,保持系和恢复系的花蕾之间ATP含量均无显著差异,不育系花蕾ATP含量均极显著低于保持系和恢复系,尤其在花粉粒成熟早期,仅为保持系和恢复系的1%;花粉粒成熟早期,不育系花蕾ATP含量下降到不足四分体时期的1/10,说明花药发育通过单核期后耗费了组织内大量的ATP,更重要的是ATP的产生可能出现了障碍;洋葱T型细胞质雄性不育系花蕾ATP含量极显著低于各时期的可育株,可能是导致其雄性不育的原因。
关键词:洋葱;T型细胞质雄性不育;花蕾;ATP含量
马蓉丽,女,研究员,主要从事蔬菜遗传育种方面的研究,E-mail:marongli1957@163.com
细胞质雄性不育(cytoplasmic male sterile,CMS)是植物中广泛存在的现象,已经在逾320个种中发现存在细胞质雄性不育(孙新菊,2015)。细胞质雄性不育的不育性状不但能被恢复系恢复育性,还可以用保持系维持其后代的不育性,因此在植物杂交育种以及核质互作研究中占有非常重要的地位。
洋葱(Allium cepa L.)是世界上栽培时间最早、栽培面积最大的蔬菜作物之一,也是我国出口创汇最多的蔬菜作物之一(梁毅 等,2009)。但由于洋葱每朵小花仅产生6粒种子,人工去雄授粉成本很高,应用雄性不育系制种省时省工,因此洋葱CMS的研究非常重要(王建军 等,2003)。
三磷酸腺苷(ATP)是以次黄嘌呤核苷酸为底物,经生物发酵技术制得的高能化合物,是体内组织细胞一切生命活动所需能量的直接来源,被誉为细胞内能量的“分子货币”,储存和传递化学能,蛋白质、脂肪、糖和核苷酸的合成都需它参与,可促使机体各种细胞的修复和再生,增强细胞代谢活性(吕俊恒和邓明华,2014)。
根据细胞核中控制育性的基因不同,洋葱CMS被分成S型和T型两种(吴海涛 等,2009)。S型不育由1对隐性基因(msms)控制,最早在洋葱品种意大利红中发现(Jones & Emsweller,1936);T型不育受3对基因控制:1对相对独立的基因(A)和2对互补基因(B和C),在法国洋葱品种Jaunepaille des venus中首先发现(Berninger,1965)。
吴海涛(2010)采用分子标记方法鉴定出洋葱品种沙沟红皮的不育系8A为T型CMS(T-CMS)。为了初步判断洋葱T-CMS是否在小孢子发育能量需求旺盛阶段通过减少ATP的合成量诱导花粉败育,本试验对该材料的小孢子败育前后的ATP含量进行测定分析,旨在为洋葱T型细胞质雄性不育系的进一步研究与利用奠定基础。
1 材料与方法
1.1试验材料
供试洋葱T型细胞质雄性不育系、保持系和恢复系均从山西地方品种沙沟红皮中选出,回交10代,为近等位基因系。不育系材料为8-43A,保持系材料为8-43B,恢复系材料为8-43-57C,均由山西省农业科学院蔬菜研究所提供。
1.2试验方法
2012年8月下旬播种不育系、保持系和恢复系洋葱种子于山西省农业科学院东阳试验基地,11月中旬囤苗越冬,翌年3月中旬定植,7月中旬收获,贮藏越夏,10月中旬定植种球,2014年5月下旬抽薹现蕾。
1.2.1不同时期花蕾的分级和取样 在开花初期于上午8:00~10:00在田间摘取可育系发育程度不同的新鲜花球,撕开外苞片,测量不同大小花蕾的横径、纵径,剥取花药,在载玻片上挤压出小孢子,用0.5%的醋酸洋红染色,光学显微镜下观察并统计小孢子发育时期。
根据显微观察结果,分别选取不育系、保持系和恢复系小孢子处于四分体时期、单核花粉粒时期和花粉粒成熟早期的花蕾,置于液氮中保存备用。
1.2.2 ATP含量测定ATP含量测定基于在ATP存在的条件下,萤火虫荧光素酶可催化底物D-荧光素氧化并发射荧光(约560 nm,pH= 7.8),并且在除ATP外其他底物处于过量的情况下,光子数量与ATP含量呈线性关系这一原理。取保存的花蕾样品各0.5 g,用ENLITEN®ATP Assay System Bioluminescence Detection kit(Promega)提取小孢子发育不同时期花蕾中的ATP,在GloMax®20/20 Luminometer(Promega)仪器上进行荧光强度测定。根据标准曲线计算单位质量花蕾中ATP的含量,设3次重复,取平均值。
1.3数据处理
采用Microsoft Excel 2010软件进行数据整理,采用SPSS 19.0软件进行各指标的差异显著性分析。
2 结果与分析
2.1洋葱花球外部形态与花粉发育时期的对应关系
观察发现,当花球外苞充实约1/3时(图1-a),花蕾横径为0.21~0.23cm、纵径为0.25~0.32cm(图1-b),花药呈浅绿色、半透明状、多汁液,其中大部分花粉处于四分体时期(图1-c);当花球外苞充实约2/3时(图1-d),花蕾横径为0.23~0.27cm、纵径为0.31~0.35cm(图1-e),花药呈浅绿色、不透明、多汁液,其中大部分花粉处于单核花粉粒期(图1-f);当花球外苞即将或刚开裂时,花蕾横径为0.27~0.33cm、纵径为0.35~0.39cm(图1-g),花药呈绿色、不透明、少汁液,其中大部分花粉处于花粉粒成熟早期(图1-h)。

图1 洋葱可育系不同发育时期花球外部形态和花粉发育时期
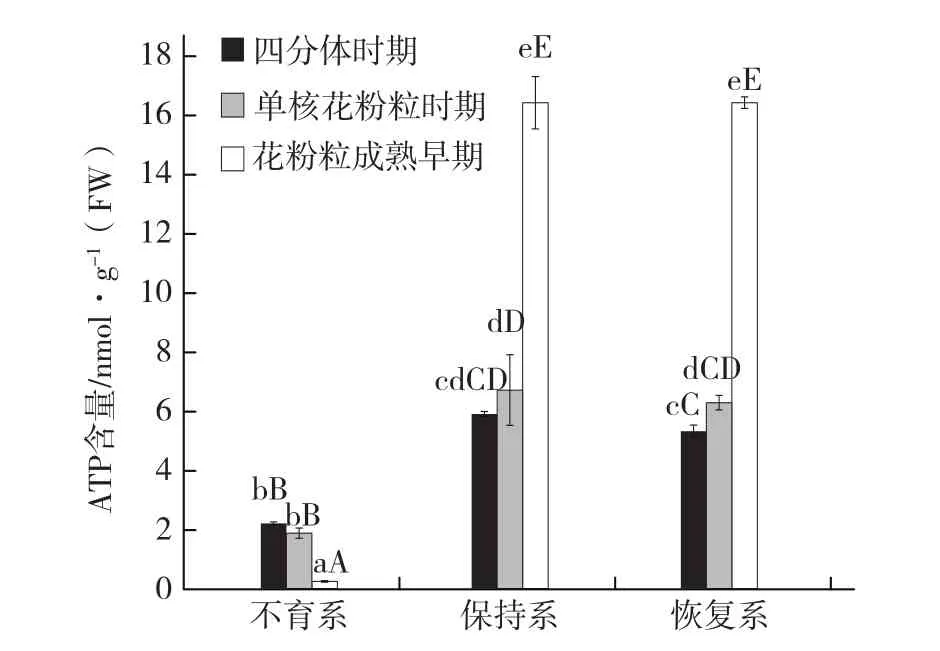
2.2洋葱T型细胞质雄性不育与花蕾ATP含量的关系
四分体时期18 16 14 12 10 eE eE单核花粉粒时期)花粉粒成熟早期FW1(mo1·g-86420 量/ndD cdCD dCD cC 含ATP bBbB aA不育系 保持系 恢复系
由图2可知,从洋葱花药四分体时期到单核花粉粒时期,T-CMS不育系和保持系花蕾ATP含量的变化均不显著。到花粉粒成熟早期,不育系花蕾ATP含量下降到不足四分体时期的1/10,而保持系和恢复系则增加到四分体时期的3倍左右。不育系花蕾ATP含量在各时期均极显著低于保持系和恢复系,尤其在花粉粒成熟早期,仅为可育系的1%。在整个花药发育过程中,保持系和恢复系的花蕾ATP含量均无显著差异。
3 结论与讨论
本试验采用萤火虫荧光素酶检测技术测定了洋葱T-CMS不育系、保持系和恢复系在花蕾不同发育时期的ATP含量,发现不育系花蕾ATP含量在各时期均极显著低于保持系和恢复系,尤其在花粉粒成熟早期,仅为可育系的1%。多种作物的研究中都发现,植物花粉败育过程中ATP的含量呈普遍降低趋势,玉米和高粱的CMS的形成与ATP含量变化有着密切关系,可育系花药中的ATP含量均高于不育系(王秀珍 等,1986);水稻从单核早期开始,不育系花药的ATP含量一直低于可育系,可育系花药的ATP含量比不育系高7倍以上(陈贤丰和梁承邺,1991);而在不育系中由于ATP含量较低,不足以提供花粉萌发或者花粉管伸长所需的能量,最终使得小孢子发育被迫停止,从而导致不育(邓继新 等,1990)。此外,在玉米(夏涛和刘纪麟,1994)、萝卜(魏毓棠 等,2001)雄性不育的研究中也发现了能量亏损现象。夏涛和刘纪麟(1994)在玉米三种(T、C、S)CMS不育系中均发现,随着小孢子发育的进行,花药组织ATP含量显著降低。萝卜雄性不育小孢子发育过程中,向花蕾中运输淀粉等物质会导致能量积累不足而败育(张丽 等,2002)。本试验结果表明,洋葱T-CMS不育系花粉粒成熟早期的花蕾ATP含量下降到不足四分体时期的1/10,说明花药发育通过单核期后耗费了组织内的大量ATP,更重要的是ATP的产生可能出现了障碍。这些现象表明,伴随着小孢子发生的开始和小孢子败育的发生,不育系花药组织中ATP含量及其变化出现了异常。这种异常必然会影响花药组织及小孢子物质合成和能量代谢以及与之有关的生命活动,导致小孢子发育的畸形和败育。下一步可以从影响ATP合成的因素出发,应用细胞学、遗传学、生理生化、分子生物学等手段进行研究,阐明洋葱T型细胞质雄性不育的机理。
参考文献
陈贤丰,梁承邺.1991.不育花药能量代谢的积累与雄性不育的关系.植物生理学通讯,27(1):21-24.
邓继新,刘文芳,肖翊华.1990.HPGMR花粉发育期花药ATP含量及核酸与蛋白质的合成研究.武汉大学学报:自然科学版,(3):85-88.
梁毅,王永勤,于春霞,关绚丽,张智,田保华.2009.中国洋葱产业的回顾与展望.中国农学通报,25(24):308-312.
吕俊恒,邓明华.2014.植物雄性不育与能量代谢的关系.辣椒杂志,(1):1-6.
孙新菊.2015.植物细胞质雄性不育的研究进展.现代园艺,(1):16-17.
王建军,侯喜林,宋慧,王学文.2003.洋葱育种研究进展.中国蔬菜,(4):57-59.
王秀珍,滕晓月,阎隆飞,周人纲.1986.玉米及高粱花药中三磷酸腺苷(ATP)含量与细胞质雄性不育的关系.作物学报,12(3):177-181.
魏毓棠,张丽,张大昕,梁守连.2001.萝卜雄性不育小孢子发育过程中物质代谢的研究.辽宁农业科学,(4):8-10.
吴海涛,马蓉丽,刘洪炯,焦彦生.2009.洋葱细胞质雄性不育系选育研究进展.园艺学报,36(2):297-302.
吴海涛.2010.洋葱细胞质雄性不育相关基因的结构与表达模式研究〔博士论文〕.南京:南京农业大学.
夏涛,刘纪麟.1994.玉米雄性不育细胞质细胞色素氧化酶活性及ATP含量的研究.华北农学报,9(4):33-37.
张丽,李霄燕,魏毓棠,田爱民.2002.萝卜雄性不育小孢子发育过程中物质代谢的研究.安徽农业科学,30(3):326-327.
Berninger E.1965.Contribution a L’etude de la sterilite-male de L’oignon(Allium cepa L.).Annual Amelior Plantes,15:183-199.
Jones H A,Emsweller S L.1936.A male-sterile onion.Proceeding of the American Society for Horticultural Science,34:582-585.
Relationship between ATP Contents of buds and T-type Cytoplasmic Male Sterility in Onion
MA Rong-li1,CHENG Yan1,WU Hai-tao1,2*,JIAO Yan-sheng1,QIAO Ning1
(1Institute of Vegetables,Shanxi Academy of Agricultural Sciences,Taiyuan 030031,Shanxi,China;2Wenzhou Professional College of Science and Technology,Wenzhou 325000,Zhejiang,China)
Abstract:Taking onion T-type cytoplasmic male sterile line ‘8-43A’,maintainer line ‘8-43B’and restorer line ‘8-43-57C ’as material,using firefly luciferase enzyme testing technology,this paper analyzed the changes in ATP contents of buds at different growing stages,and studied the relationships between ATP contents of buds and T-type cytoplasmic male sterility in onion.The results showed that there were no significant differences between the ATP contents of maintainer and restorer lines throughout the entire development process of bud.While,the ATP content ofcmS bud was very significantly lower than that of the maintainer and restorer lines,especially at early grain stage,which only took 1% of that in maintainer and restorer lines.ATP content of thecmS line at early grain stage decreased to less than 1/10 of those at tetrad stage,illustrating a large quantity of ATP was consumed after mononuclear stage.And an even important thing was there might be barriers in ATP generation.The ATP content of T-typecmS in onion was very significantly lower than that of the fertile lines might be the reason for male sterility.
Key words:Onion;T-type cytoplasmic male sterility;Bud;ATP content
基金项目:国家自然科学基金青年基金项目(31201638)
收稿日期:2015-08-12;接受日期:2015-12-09
*通讯作者(
Corresponding author):吴海涛,男,博士,助理研究员,主要从事蔬菜遗传育种方面的研究,E-mail:cucum2008@126.com
