东太湖水生植物季节演替及抑藻效应初探
2016-03-31李灵慧吴建斌陆雪林崔恒钊杨晨驰鲍婷婷李建华
李灵慧,吴建斌,陆雪林,崔恒钊,杨晨驰,鲍婷婷,李建华*
(1.同济大学环境科学与工程学院,上海 200092; 2.苏州市吴江区水利局,江苏苏州 215222; 3.上海市向明中学,上海 200092)
东太湖水生植物季节演替及抑藻效应初探
李灵慧1,吴建斌2,陆雪林2,崔恒钊1,杨晨驰1,鲍婷婷3,李建华1*
(1.同济大学环境科学与工程学院,上海 200092; 2.苏州市吴江区水利局,江苏苏州 215222; 3.上海市向明中学,上海 200092)
摘 要:为了研究东太湖水生植物植物时空分布特征及水环境因子变化趋势,同时探究东太湖水生植物对藻类的影响,于2013年对东太湖水生植物和水体环境状况进行调查。调查发现,采样区内水生植物共14种,其中,沉水植物6种,浮叶植物6种,挺水植物2种,并且东太湖水生植物不同季节优势种不尽相同,并表现出明显的区域特征。从环境因子对水生植物的影响来看,东太湖水生植物与温度呈显著正相关关系,与TP无显著相关关系。就东太湖水生植物抑藻效果而言,水体中Chl-a浓度随着水生植物生物量增加而减少,水生植物的抑藻效果明显。
关键词:东太湖;水生植物;水体环境;抑藻效应
DOI 10.16178/j.issn.0528-9017.20160247
水生植物作为湖泊生态系统中的初级生产者,具有显著的环境生态功能,是良性湖泊生态系统的重要组成部分,为维持湖泊生态系统平衡做出重要贡献。一方面,水生植物为水体中的鱼类提供栖息场所、食物来源,为浮游生物和周丛生物提供避难与附着场所[1]。另一方面,水生植物通过对生境资源的争夺以及分泌感化物种抑制水中藻类的过度生长[2-3]。与此同时,水生植物对水环境亦有着密切的相互作用,既有提高水体透明度,吸收同化营养盐的环境正效应[4-5],亦有因残体腐败污染水体,消耗溶解氧[6],过量生长促淤化等环境负效应[7-8]。
东太湖位于太湖东南隅(30°58′—31°07′N,120°25′—120°35′E),与西太湖之间以狭窄的湖面相通,总长度27.5 km,最大宽度9.0 km,总面积131 km2,平均水深1.7 m,湖区内生长着繁茂的水生植物,是长江中下游典型的草型浅水湖泊[9]。随着近年来当地经济持续快速发展,东太湖周围地区供水安全不断面临着挑战,研究表明,东太湖水体总体上处于中富营养状态,部分湖区已达到富营养状态[10],存在着蓝藻水华爆发的风险[11],但近几年,东太湖并未出现蓝藻水华的相关报道,这可能因为东太湖属于草型湖泊,水生植物对东太湖藻类有一定的抑制作用。
为了研究东太湖水生植物季节演替特点,以及水生植物与藻类的相互关系,对东太湖水源地的水生植物及水环境因子进行调查和分析,研究水生植物植物时空分布特征及水环境因子变化趋势,同时探究东太湖水生植物生物量对藻类的影响,为草型湖泊的科学管理提供理论依据。
1 研究方法
1.1采样点布置
根据东太湖功能及不同利用方式的区域,布设10个采样点,分布见图1。其中C3,C4,C7和C9点设在敞水航道区,C5,C8,C10,C11,C12 和C13点设在围栏养殖区内。
1.2样品采集与分析
于2013年4,6,8,10和12月对东太湖进行5次采样。溶解氧、水温和pH采用便携式分析仪在现场直接测定,透明度采用直径25 cm的透明度盘进行测定。现场用1 L聚乙烯塑料瓶取0.5 m处表层水,带回实验室进行营养盐测定,营养盐测定参考《水和废水监测分析方法(第四版)》。
1.3植物样本采集与测定
使用自制的水草采集刀,将长为0.12 m的刀头绑在3 m竹竿上,靠近底泥旋转一周,采集0.045 m2面积的水生植物地上部分。同一采样点采集2次以保证样品采集完全。样品采集后立即用湖水清洗去泥,去除枯枝、败叶和其他杂质,装入聚乙烯袋中运回实验室,去除植物体表多余的水分,按种类称其鲜重,求出2次采集样品的重量平均值,计算得出样点植物生物量。受采样条件限制,生物量估算不涉及挺水植物的调查。
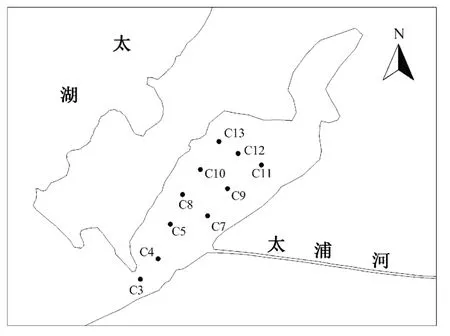
图1 采样点示意图
2 结果与分析
2.1水生植物种类组成及时空分布
根据2013年4月至12月5次调查区域采样结果看,共有水生植物14种。其中,沉水植物6种,分别为金鱼藻(Ceratophyllumdemersum),穗花狐尾藻(Myriophyllumspicatum),苦草(Vallisneriaspiralis),马来眼子菜(Potamogetonmalaianus),微齿眼子菜(Potamogetonmacckianus ),伊乐藻(Elodea nuttalli );浮叶植物6种,分别为荇菜(Nymphoidespeltatum),野菱(Trapamaximowiczii),金银莲花(Nymphoidesindica ),空心莲子草(Alternantheraphiloxriides ),凤眼莲(Eichhorniacrassipes),无根萍(Wolffiaarrhiza);挺水植物2种,分别为菰草(Zizaniacaduuciflora),莲(Nelumbonucifera) (表1)。沉水植物的优势种为:伊乐藻、穗花狐尾藻、金鱼藻,浮叶植物优势种为:荇菜、野菱和金银莲花。

表1 东太湖水生植物空间分布
东太湖水生植物生物量季节变化见图2。4月时,东太湖调查区域大部分水生植物还未萌发,小部分水生植物,主要是浮叶和部分沉水植物刚刚萌发,此时水生植物整体覆盖率很低。荇菜是春季唯一的优势种,金鱼藻、苦草、伊乐藻、金银莲花等水生植物在东太湖各采样点偶见,其均处在萌发初期,盖度不大。6月,沉水植物金鱼藻、穗花狐尾藻、苦草和马来眼子菜萌发生长,在水中可采集到。浮叶植物6月开始大量生长,成为优势种,人为引种的沉水植物伊乐藻也在此季节大量发生,进入生长旺盛期,也成为优势种。8月,荇菜进入结实衰亡期,沉水植物大量发生,东太湖8月水生植物最为繁茂的时期,此时水生植物生物量最大,平均值为1 831 g·m-2。10月,大部分植物进入结实衰亡期,穗花狐尾藻、苦草、空心眼子菜以及伊乐藻等沉水植物开始枯萎,荇菜和金银莲花大量枯萎,漂浮在水面上,而孔欣欣莲子草因空白生态位的出现,此时大量发生,成为优势种。12月,进入冬季后,绝对大部分水生植物已经消失,仅有处在越冬期的沉水植物金鱼藻和苦草,以及残余的浮叶植物金银莲花。
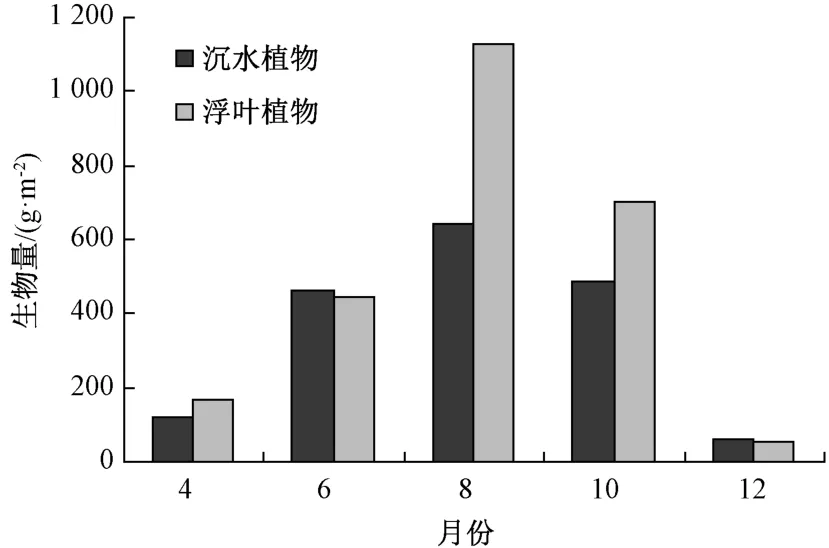
图2 水生植物生物量时间分布
东太湖水生植物生物量时空分布见图3。研究表明,围网养殖区(C5,C8,C10,C11,C12,C13)的生物量与敞水航道区(C3,C4,C7,C9)并未出现显著性变化,这与养殖区内渔民大量收割伊乐藻从而整体生物量下降有关。在研究区域内,采样点C7的年平均生物量最高,为1 217.2 g·m-2; C11的年平均生物量最小,仅为406 g·m-2。C7水生植物主要以浮叶植物金银莲花和野菱为主,沉水植物生物量较少,而C11主要以浮叶植物金银莲花和沉水植物穗花狐尾藻为主。此外,荇菜、野菱和金银莲花在敞水航道区的年平均生物量显著高于围网养殖区,主要因为航道区的水文条件适合浮叶植物的生长[12]。
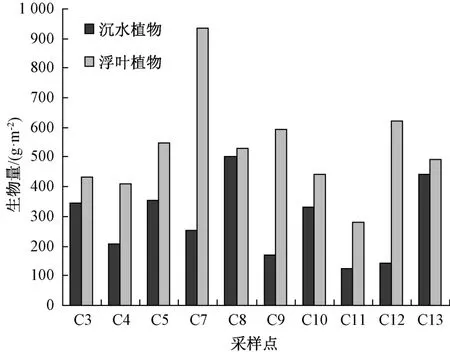
图3 水生植物生物量空间分布
2.2水质指标变化分析
表2显示,东太湖平均水温最高出现在6月,为(29.34±0.64)℃;最低出现在12月,为(7.47±0.20)℃。各采样点间的水温在季节内差异并不明显。

表2 东太湖水体环境因子指标
透明度随季节变化明显。夏秋两季透明度显著高于春冬两季,这与夏秋两季温度升高,水生植物的大量生长及沉降水体悬浮固体颗粒的作用有关[13]。空间变化上,围网养殖区内样点透明度显著高于敞水航道区,这是因为东太湖以围网养殖河蟹为主,而围网具有阻流作用,不利于污染物及颗粒物的扩散[14-15]。
pH总体上呈中性偏弱碱性,且具有较明显的季节性变化趋势,即冬季和春季的pH值高于夏季和秋季;各月样点间pH差异不大。
TN,TP季节变化明显,春季明显高于其他季节。春季水生植物生物量较小,对水体中的营养盐利用率较低。随温度升高,水生植物生物量明显增加,增加了水体中营养物质的吸收,从而使水体中营养盐降低。而10和12月水体中TN,TP含量明显升高,这可能与水生植物腐败,体内的营养盐再次释放到水体中有关。
2.3水生植物与水环境相关性分析
温度是影响水生植物的一个重要环境因子。水生植物在适宜的温度范围内,会随着温度的升高而快速生长。伊乐藻在水温5~30℃时能正常生长,最适温度在25℃左右[16]。合适的水体透明度能保证足够的光照投射至湖底,为水生植物的生长提供充足的光源。同时,水生植物的生长通过吸附、截留等作用使水体悬浮固体颗粒沉降,提高水体的透明度[13]。野菱和金银莲花与TN呈显著负相关,说明水生植物的生长需要吸收大量的氮营养盐[17]。与氮相比,水体中磷的情况有所不同。相比水生植物对N的消耗作用,水生植物对水体中的磷具有两面性,可将磷从沉积物中转移到水中[18],使水中的磷沉降回沉积物中,故东太湖水生植物的生长与磷元素相关性较小。
由表3可知,温度与水生植物呈显著正相关,穗花狐尾藻、马来眼子菜和伊乐藻等沉水植物与透明度呈显著正相关,浮叶植物与溶解氧呈显著正相关,野菱和金银莲花与TN呈显著负相关,水生植物与TP无显著相关关系。
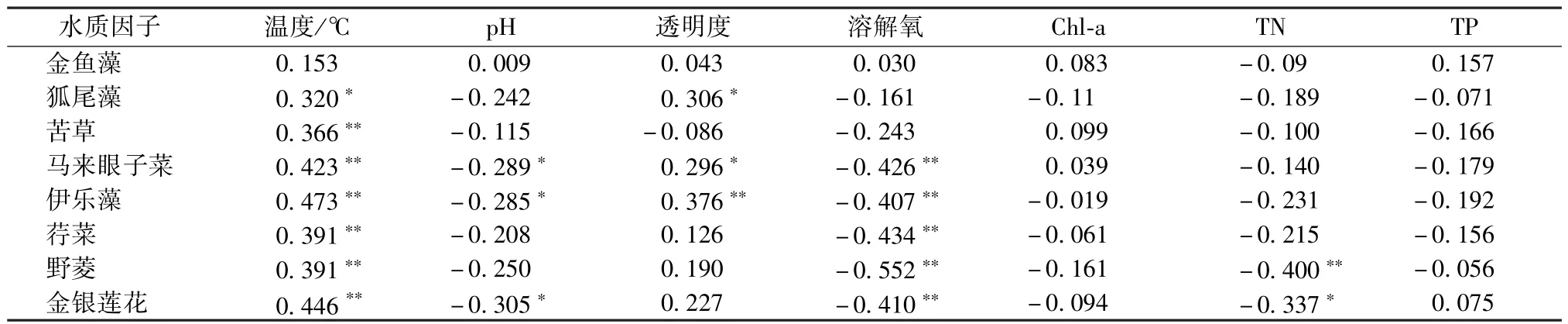
表3 东太湖主要水生植物与水质因子的相关关系
2.4水生植物抑藻效应初探
水生植物作为水域生态系统的重要组成部分,其抑藻机理主要是水生植物对水体中营养盐、光照的吸收明显优于藻类,为食藻型鱼类提供避难场所,增加水体中藻类的去除效率,以及部分水生植物能够分泌化感物质,抑制水体中藻类的生长[19-21]。研究表明,水生植物生物量与藻类生长密切相关。4月,东太湖水体中水生植物生物量较低,而Chl-a浓度较高(图4)。随着水体中水生植物生物量的增加,水体中Chl-a的浓度逐渐降低; 当8月水生植物生物量达到全年最高,水体中Chla的浓度也达到全年最低,后随水生植物生物量的减少,Chl-a的浓度逐渐增加。在12月,虽然水生植物生物量达到全年最低,但水体中的Chl-a浓度并未明显回升。这可能是因为12月东太湖水温较低,温度成为抑制藻类生长的主要因素。
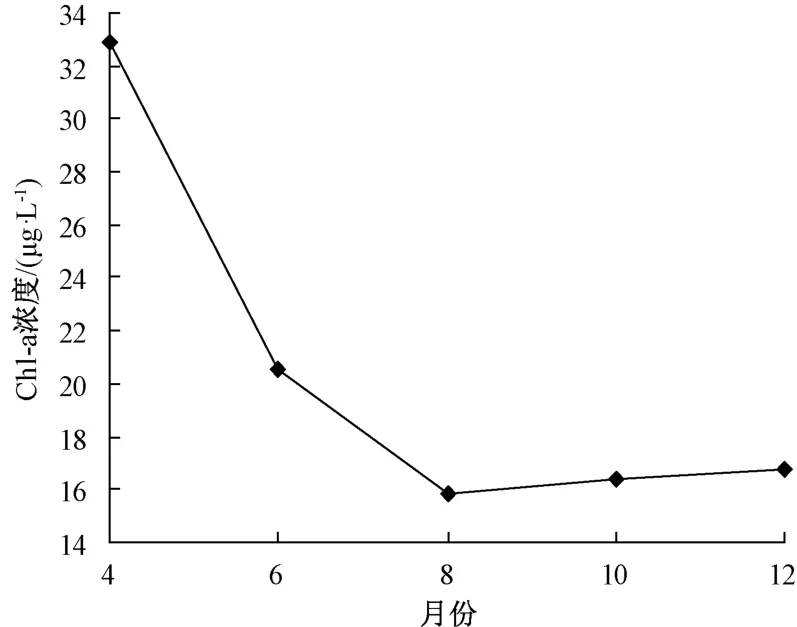
图4 Chl-a浓度季节分布
3 小结
调查发现,东太湖主要水生植物共14种,其中沉水植物6种,浮叶植物6种,挺水植物2种。水生植物不同季节优势种不尽相同,并表现出明显的区域特征。在围网养殖区(C5,C8,C10,C11,C12,C13)水生植物生长茂盛,主要优势种为伊乐藻、金鱼藻等沉水植物;而敞水航道区(C3,C4,C7,C9)水生植物生物量较小,植物种类单一,优势种主要为浮叶植物如金银莲花、荇菜、野菱等,亦有沉水植物伊乐藻、穗花狐尾藻等作为优势种。
水体环境不同水环境因子表现出明显的时空变化趋势,TN,TP浓度的季节变化为春季明显高于其他季节,且水生植物的生长受水体环境如温度、透明度影响较大,受TP的影响不显著。
东太湖水生植物抑藻效果明显,水生植物生物量与水体中Chl-a浓度呈明显负相关。因此,东太湖可通过控制水体中水生植物的生物量,从而控制水体中藻类的过度生长,防止水华爆发。
参考文献:
[1]摇叶春.洱海湖滨带生态恢复工程模式研究[M].北京:中国环境科学研究院,1999.
[2]GOPAL B,GOEL U.Competition and allelopathy in aquatic plant communities[J].Botanical Review,1993,59 (3):155-210.
[3]章宗涉.水生高等植物:浮游植物关系和湖泊营养状态[J].湖泊科学,1998,10 (4):83-86.
[4]HAVENS K E,FUKUSHIMA T,XIE P,et al.Nutrient dynamics and the eutrophication of shallow lakes Kasumigaura (Japan),Donghu (PR China),and Okeechobee (USA)[J].Environmental Pollution,2001,111 (2):263-272 (10) .
[5]吴振斌,邱东茹,贺锋,等.水生植物对富营养水体水质净化作用研究[J].武汉植物学研究,2001,19 (4):299-303.
[6]MIRANDA L E,HODGES K B.Role of aquatic vegetation coverage on hypoxia and sunfish abundance in bays of a eutrophic reservoir[J].Hydrobiologia,2000,427 (1 ):51-57.
[7]FRODGE J D,THOMAS G L,PAULEY G B.Effects of canopy formation by floating and submergent aquatic macrophytes on the water quality of two shallow Pacific Northwest lakes[J].Aquatic Botany,1990,38 (90):231-248.
[8]ROMAN C T,Barrett N E,Portnoy J W.Aquatic vegetation and trophic condition of Cape Cod (Massachusetts,USA) kettle ponds[J].Hydrobiologia,2001,443 (1/3):31-42.
[9]陈开宁,范成新.东太湖水环境质量调查及保护对策[J].海洋湖沼通报,1996 (1):9-17.
[10]白秀玲,谷孝鸿,杨龙元,东太湖水环境现状及保护对策[J].湖泊科学,2006,18 (1):91-96.
[11]WU G,XU Z.Prediction of algal blooming using EFDC model:Case study in the Daoxiang Lake[J].Ecological Modelling,2011,222 (6):1245-1252.
[12]STEFFEN K,LEUSCHNER C,MüLLER U,et al.Relationships between macrophyte vegetation and physical and chemical conditions in northwest German running waters[J].Aquatic Botany,2014,113 (3):46-55.
[13]何俊,谷孝鸿,刘国锋,东太湖水生植物及其与环境的相互作用[J].湖泊科学,2008,20 (6):790-795.
[14]马剑敏,严国安.武汉东湖受控生态系统中水生植被恢复结构优化及水质动态[J].湖泊科学,1997,9 (4):359-363.
[15]王瑁,顾宇飞,朱增银.不同营养状态下金鱼藻的生理响应[J].应用生态学报,2005,16 (2):337-340.
[16]朱松泉,刘正文.伊乐藻-草鱼圈养人工复合生态系统建设的研究[J].湖泊科学,1996 (8):46-61.
[17]OZIMEK T,DONK E V,GULATI R D.Growth and nutrient uptake of two species of Elodea in experimental conditions and their role in nutrient accumulation in a macrophyte dominated lake[J].Hydrobiologia,1993,251 (1/3):13-18.
[18]CARPENTER S R.Submersed vegetation:an internal factor in lake ecosystem succession[J].American Naturalist,1981,118 (3):372-383.
[19]王立新,吴国荣,王建安,等.黑藻(Hydrilla verticillata)对铜绿微囊藻(Microcystis aeruginosa)抑制作用[J].湖泊科学,2005,16 (4):337-342.
[20]雷泽湘,徐德兰,黄沛生,等.太湖沉水和浮叶植被及其水环境效应研究[J].生态环境,2006,15 (2 ):239-243.
[21]RUGGIERO A,SOLIMINI A G,CARCHINI G.Nutrient and chlorophyll a temporal patterns in eutrophic mountain ponds with contrasting macrophyte coverage[J].Hydrobiologia,2003,506/509 (1/3):657-663.
(责任编辑:张瑞麟)
通信作者:李建华,E-mail:leejianhua@tongji.edu.cn。
作者简介:李灵慧(1991—),女,在读硕士,研究方向为水域生态学,E-mail:lilinghui789@163.com。
基金项目:上海市科学技术委员会科研计划项目(13231203703,13DJ1400104)
收稿日期:2015-10-14
中图分类号:S682.32
文献标志码:A
文章编号:0528-9017(2016)02-0288-04
文献著录格式:李灵慧,吴建斌,陆雪林,等.东太湖水生植物季节演替及抑藻效应初探[J].浙江农业科学,2016,57 (2):288-291,293.
