葡萄光合作用光响应曲线拟合模型比较研究
2016-03-23白云岗刘洪波张江辉
白云岗,刘洪波,张江辉,冯 杰,丁 平
(新疆水利水电科学研究院,乌鲁木齐 830049)
0 引 言
光合作用是植物生长发育的基础,其效率与自身因素如叶绿素含量、叶片厚度和叶片成熟程度密切相关,又受外界环境因子如光强度、气温、空气相对湿度和土壤含水量等影响[1-3]。光响应曲线反映出净光合速率随光强改变而变化的规律[4-7],其光合速率、光饱和点、光补偿点、表观量子效率等可由植物光响应曲线估算的光合参数是植物重要的生态生理参数[8]。因此,研究光合作用的光响应曲线对了解植物光合效率及其生长规律非常重要。目前,常用的光响应模型有二次多项式模型[9,10]、指数函数模型[11-13]、直角双曲线模型[14]、非直角双曲线模型[15]和修正的直角双曲线模型[16-21],同时也对甘肃、陕西等地的红地球、巨峰等葡萄品种的光合特性进行了研究[25],而对于极端干旱的吐鲁番地区的主要特色林果葡萄的光响应曲线的研究鲜见。为此,通过分析不同模型下光合参数模拟值与实测值的接近度,选出适合该地区葡萄光响应曲线的模型,为吐鲁番地区的葡萄管理以及节水灌溉等相关研究提供基本的理论依据。
1 材料与方法
1.1 研究区概况
试验点位于火焰山以南,吐鲁番市东南部的葡萄乡铁提尔村,距吐鲁番市区12 km,地处北纬42°56′,东经89°13′,海拔-68.8 m。年均降雨量为16.6 mm,年均蒸发量为3 300 mm,地下水位为30 m,年均气温为14.4 ℃,多年最高气温、最低气温分别为48.3 ℃,-28.8 ℃,10 ℃以上活动积温为5 455 ℃,全年年均日照时数为3 095 h,无霜期达210 d。葡萄品种为无核白(Thompson Seedless),中晚熟品种,所需大于10 ℃的活动积温在3 300 ℃左右。大部分葡萄从1998年开始定植,栽培沟为东西走向,沟长为60 m,沟宽为1.0~1.2 m,沟深约为0.5 m;株距为1.2~1.5 m,行距3.5~4.5 m,栽培方式为小棚架栽培,棚架前段高2.0 m,后端高 0.8 m,平均高1.2 m。土壤为黏壤土,质地较均一。
试验采用3管布设,栽培沟内葡萄主干旁布设1条滴灌管,栽培沟内葡萄主干旁靠近垄一侧布设2条滴灌管,滴头流量为3.2 L/ h,滴头间距均为40 cm。葡萄生育期共灌水17次,灌水定额450 m3/hm2,灌溉定额9 300 m3/hm2。
1.2 光响应曲线测定
在葡萄果粒膨大期(7月)晴朗天气,随机选取生长一致的健壮植株,采用CIRAS-3型便携式光合仪(美国PP-systems公司)通过控光、控CO2测定光合响应曲线,2次重复,温度设置为(25±1)℃,空气相对湿度(RH )为60%~70%,设定CO2摩尔分数为400 μmol/mol,光照强度设置为0、50、100、150、200、300、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol/(m2·s)。
应用SPSS 22软件对研究对象的光响应曲线和CO2摩尔分数响应曲线进行拟合,用Excel 2010绘图。
2 结果与分析
2.1 葡萄光合作用的光响应曲线
光强响应曲线反映了植物光合速率随光照强度改变而变化的规律,光强响应曲线如图1所示。由图1可看出,光强在0~600 μmol/(m2·s)间增强时,净光合速率迅速上升。当光照强度在600~1 500 μmol/(m2·s)时,净光合速率升幅随光强的增加而减弱。当光照强度在1 500~2 000 μmol/(m2·s)时,净光合速率随着光照强度的增加而出现下降,表现出光抑制现象,即植物叶片接受的光能超过了其所能利用的光量时,长时间的强光照射可以引起叶片光合活性的降低,光抑制是植物光合作用非气孔限制因素的主要形式。
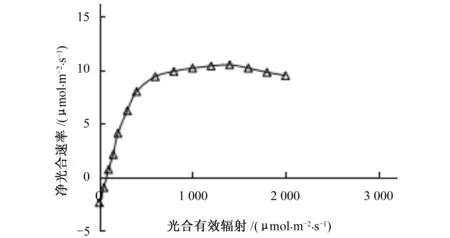
图1 葡萄光强响应曲线
图2为葡萄低光强响应曲线。从图2可看出,光合有效辐射在200 μmol/(m2·s)以下的线性方程拟合效果很好,决定系数(R2)为0.996。
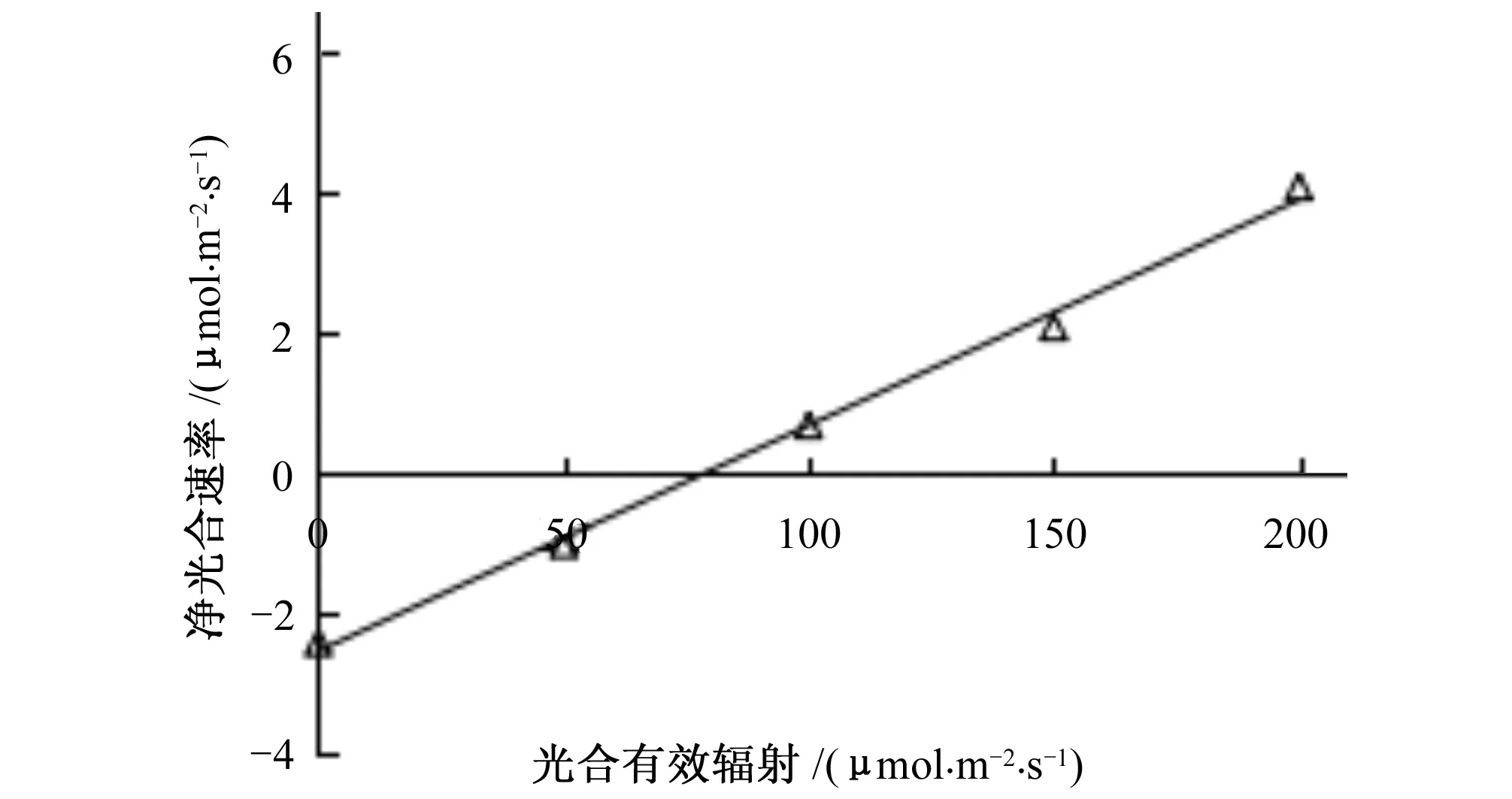
图2 葡萄低光强响应曲线
2.2 葡萄光响应曲线模型拟合值对比
从表1可看出,二次多项式模型的拟合值与实测值相差最大,其余4种模型在各光合有效辐射下的净光合速率值均比较接近,其中非直角双曲线模型拟合值最为接近实测值,其次是修正的直角双曲线模型。
同时,指数函数模型和直角双曲线模型均不能较好地解释在光饱和区域的净光合速率与光合有效辐射的相关性问题,而二次多项式模型、非直角双曲线模型和修正的直角双曲线模型的拟合曲线则体现出葡萄在高光强下光合作用的光抑制现象,即净光合速率达到光饱和点后,随着光合有效辐射的增加,净光合速率反而降低。且由于非直角双曲线模型拟合得到的净光合速率相比其他4种模型的拟合值更接近实测值,可见,非直角双曲线模型在葡萄光响应曲线中对净光合速率的拟合效果优于二次多项式模型、指数函数模型、直角双曲线模型和修正的直角双曲线模型。
2.3 葡萄光响应曲线模型拟合参数
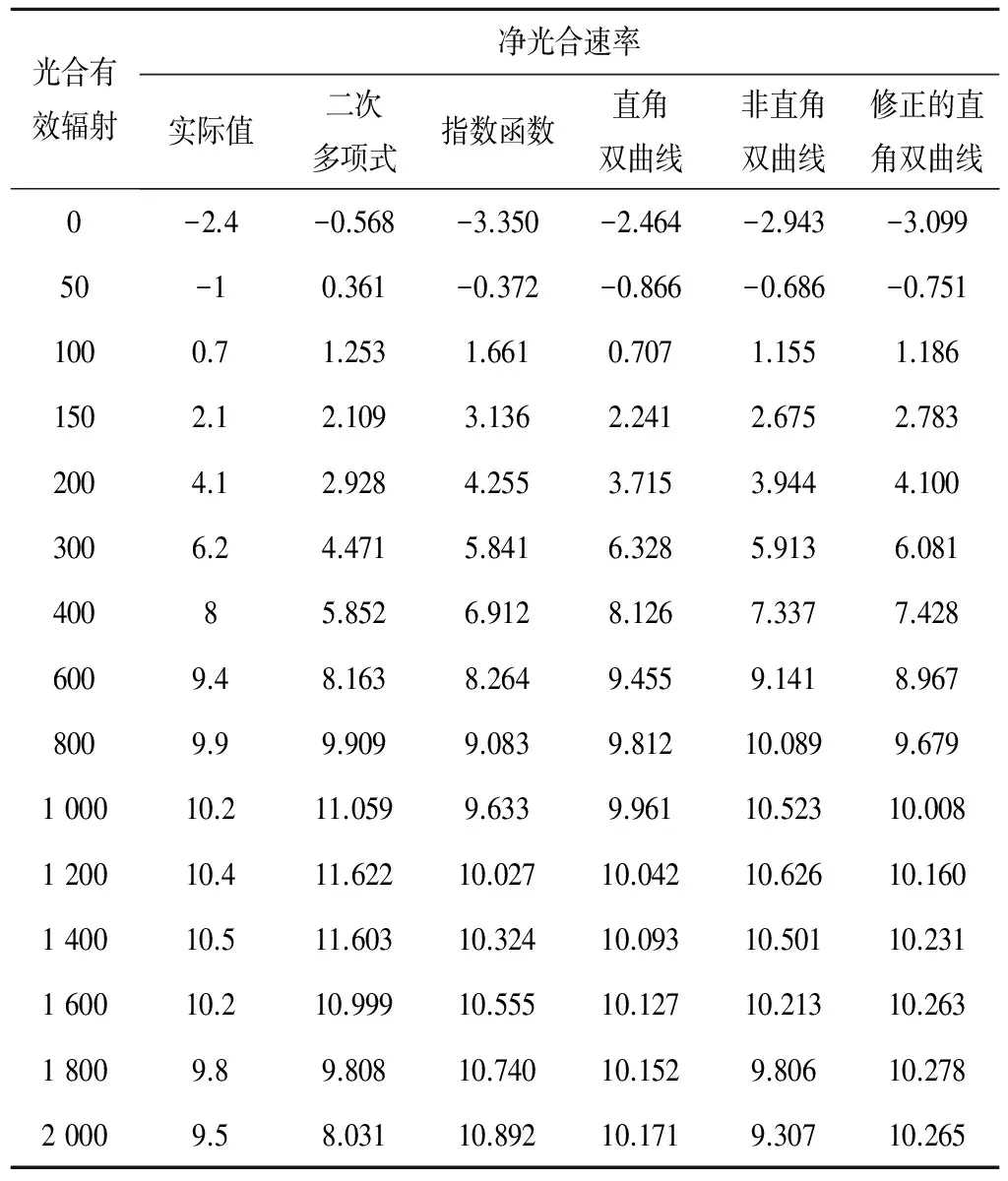
表1 葡萄光合作用光响应曲线实测值及其拟合值 μmol/(m2·s)
表2为葡萄光响应曲线在5种模型拟合下的光合参数,由相关性检验可知,5种光响应模型的拟合值与葡萄光响应曲线的相关关系均达到极显著水平,其中,非直角双曲线模型和修正的直角双曲线模型的决定系数最高,为0.996和0.994。就模型模拟得到的最大净光合速率而言,二次多项式模型在光响应曲线研究中只作为一个纯数学模型被应用,可以直接求出光饱和点、光补偿点和暗呼吸速率等参数,虽然二次多项式模型拟合得到的饱和光强与实测值最为接近,但其拟合得到的其他光合参数中有违背生物学常识的地方,其暗呼吸速率为负值(-0.568)。直角双曲线模型和非直角双曲线模型由于自身的非收敛性,不能得出光饱和点,只有通过光合有效辐射200 μmol/(m2·s)以下的线性回归方程和最大净光合速率(Amax)来估算,2种模型得到的光饱和点分别为568.043和474.907 μmol/(m2·s),与实测值1 400 μmol/(m2·s)差别较大。指数函数模型由于假设光合速率为0.9Amax所对应的光强为饱和光强,但拟合值397.857 μmol/(m2·s)远小于实测值,而假设光合速率为0.99Amax所对应的光强为饱和光强,则无法求得。修正的直角双曲线模型可模拟得到光饱和点,其值为1 178.362 μmol/(m2·s),且能较好地处理葡萄光合速率在高光强部分下降的问题,除和一般规律不符的二次多项式模型外,其与实测值最为接近。5种模型的光补偿点、表观量子效率和暗呼吸速率的模拟值均未达到显著差异水平,其中非直角双曲线模型拟合得到的光补偿点比其他4种模型更接近实测值,其次是修正的直角双曲线模型;在最大光合速率上,5种模型中修正的直角双曲线模型拟合值10.627 μmol/(m2·s)最接近实测值10.5 μmol/(m2·s),其后依次是指数函数模型、二次多项式模型、非直角双曲线模型和直角双曲线模型,最大光合速率拟合值分别为10.291、11.746、12.772和15.771 μmol/(m2·s)。同时,尽管指数函数模型和非直角双曲线模型的决定系数(R2)均在0.99以上,但他们拟合得到的光合参数与实测值相差较大,故R2越大只能说明模型的拟合程度越高,而并不能说明拟合结果就一定与实测值相符。对比5种不同模型给出的各项光合参数,只有修正的直角双曲线模型求解得到的各项光合参数与实测值均比较接近。

表2 葡萄光响应曲线的光合参数
张保玉[15]对红色、白色酿酒葡萄和红色与白色鲜食葡萄分别采用二项式回归、直角双曲线模型和非直角双曲线模型对葡萄光响应曲线相关生理参数进行了系统的研究分析和比较,与张保玉的研究结果对比可知:葡萄表观量子效率为0.032之间,说明葡萄品种的弱光利用效率属于中下水平;光补偿点值在30.247~77.392 μmol(m2·s)之间,表明葡萄品种对弱光的适应能力较强,净光合速率最大值在10.627~15.771 1 μmol/(m2·s)之间,表明葡萄品种利用强光能力较强;光饱和点值在390~1 300 μmol/(m2·s)之间,该值与张保玉的研究中光饱和点值在400~600 μmol/(m2·s)之间相差较大,是因为前者应用的是非直角双曲线、直角双曲线模型和二次多项式,如对比直角和非直角双曲线模型,其值相差很小,同时由于试验地及葡萄品种不同,因此与张保玉的研究中二次多项式模型参数产生了差异,但都说明葡萄品种对强光的适应能力较强,是典型的阳生植物;暗呼吸值小于4 μmol/(m2·s),表明葡萄品种的暗呼吸速率较小且消耗较小。综合各参数结果表明,葡萄光响应曲线参数在吐鲁番地区和张保玉在杨凌的系统研究中的变化规律保持一致。
3 结 语
(1)5种模型拟合值的决定系数(R2)均达到极显著相关关系,但只根据R2判断模型的适合性具有片面性,还应综合考虑各参数模拟值与实测值间差异及植物自身的生长规律等。
(2)二次多项式的各项光合参数拟合值与实测值相差较大,且有违背常识错误。直角双曲线模型和非直角双曲线模型均无法直接估算光饱和点。指数函数拟合的最大净光合速率与实测值相差大且无法模拟葡萄光抑制部分的光响应数据。
(3)吐鲁番地区宜采用修正的直角双曲线模型,可以直接估算出光饱和点和光补偿点等参数,能很好地体现葡萄在不同光强下的光响应曲线,且求解得到的各项光合参数均接近实测值,其拟合得到的葡萄最大净光合速率、光饱和点、光补偿点、暗呼吸速率分别为10.627、1 178.362、67.487、2.943 μmol/(m2·s),初始量子效率为0.05。
[1] 叶子飘,于 强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1 356-1 361.
[2] Awada T, Radoglou K, Fotelli M N, et al. Ecophysiology of seedlings of three Mediterranean pine species in contrasting light regimes[J]. Tree Physiology, 2003,23(1):33-41.
[3] Thornley J H M. Mathematical Models in Plants Physiology[M].New York: Academic Press,1976.
[4] 张中峰,黄玉清,莫 凌,等.岩溶植物光合-光响应曲线的两种拟合模型比较[J].武汉植物学研究,2009,27(3):340-344.
[5] 陈银华,蒋建篇.光照强度对辣椒光合特性与生长发育的影响[J].上海农业学报, 1998,14(3):46-50.
[6] 蒋高明,韩兴国,林光辉.大气CO2浓度升高对植物的直接影响[J].植物生态学报,1997,21(6):489-502.
[7] 张 宁,孟庆伟,赵世杰,等.光胁迫下银杏光合作用的光抑制[J].西北植物学报, 1999,19(3):461-465.
[8] 刘宇锋,萧浪涛,童建华,等.非直角双曲线模型在光合响应曲线数据分析中的应用[J].中国农学通报, 2005,21(8):76-79.
[9] 郭水良,方 芳,黄 华,等.外来入侵植物北美车前繁殖及光合生理生态学研究[J].植物生态学报,2004,28(6):787-793.
[10] 刘锦春,钟章成,何跃军,等.重庆石灰岩地区十大功劳的光合响应研究[J].西南师范大学学报:自然科学版,2005,32(2):158-163.
[11] 叶子飘.光合作用对光和CO2响应模型的研究进展[J]. 植物生态学报,2010,34(6):727-740.
[12] 王满莲,冯玉龙,李 新.紫茎泽兰和飞机草的形态和光合特性对磷营养的响应[J].应用生态学报,2006,17(4),602-606.
[13] 许大全.光合作用效率[M].上海: 上海科学技术出版社,2002:2-52.
[14] Baly E C C. The kinetics of photosynthesis[J]. Proceedings of the Royal Society of London. Series B:Biological Sciences, 1935,117(804):218-239.
[15] Thornley J H M. Mathematical Models in Plant Physiology[M].London: Academic Press(Inc.),1976:86-110.
[16] Ye Z P, Yu Q. A coupled model of stomatal conductance and photosynthesis for winter wheat[J]. Photosynthetica, 2008,46(4):637-640.
[17] Ye Z P. A new model for relationship between light irradiance and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica, 2007,45(4):637-640.
[18] 叶子飘,李进省.光合作用对光响应的直角双曲线修正模型和非直角双曲线模型的对比研究[J].井冈山大学学报:自然科学版,2010,31(3):38-44.
[19] 廖小锋,刘济明,张东凯,等.野生小蓬竹的光合光响应曲线及其模型拟合[J].中南林业科技大学学报,2012,32(3):124-128.
[20] 叶子飘,王建林.植物光合-光响应模型的比较分析[J].井冈山学院学报:自然科学,2009,30(4):9-13.
[21] 佘诚棋,程 鹏,季琳琳,等.油茶光合作用光响应曲线的拟合[J].经济林研究,2012,302(1):118-122,137.
[22] 张保玉.葡萄光合作用光响应曲线的研究[D]. 陕西杨凌:西北农林科技大学, 2009.
[23] 房玉林,惠竹梅,陈 杰,等.水分胁迫对葡萄光合特性的影响[J].干旱地区农业研究,2006,24(2):135-138.
[24] 郭继英,严大义.巨峰葡萄光合特性的研究[J].北京农业科学, 1994,12(2):30-32.
[25] 严巧娣.不同土壤水分条件下葡萄叶片光合特性的比较[J].西北植物学报, 2005,25(8):1 601-1 606.
