蓝花丹结实率低的传粉生物学和繁育系统初探
2016-03-21吴佩纹高素萍赵志惠
吴佩纹,高素萍,张 硕,赵志惠
( 1.四川农业大学风景园林学院,成都611130; 2.四川农业大学园林研究所,成都611130 )
蓝花丹结实率低的传粉生物学和繁育系统初探
吴佩纹1,高素萍2*,张硕1,赵志惠1
( 1.四川农业大学风景园林学院,成都611130; 2.四川农业大学园林研究所,成都611130 )
摘要:为解释蓝花丹自然结实率低的原因,该研究从传粉生物学和繁育系统入手,采用TTC法与联苯胺-过氧化氢法分别测定花粉活力和柱头可授性的动态变化;用花粉-胚珠比( P/O比)、杂交指数( OCI)估算蓝花丹繁育系统类型并通过人工控制授粉试验验证。结果表明: ( 1)蓝花丹L型雌器官与S型雄器官、L型雄器官与L型雌器官成熟时间重叠区域较多,雌雄性器官在成熟时间上无显著差异;而S型雌器官与L型雄器官、S型雌器官与S型雄器官成熟时间重叠区域较少,但蓝花丹持续开花的模式缓解了雌雄性器官时间差异而引起的生殖隔离。( 2)蓝花丹L型花P/O为502.00±52.30,S型花P/O为482.70±87.91,OCI为4,综合判断蓝花丹的繁育系统属于专性异交型且具有异型自交不亲和的特性。综上可初步解释蓝花丹自然结实率低的原因为内外因素共同作用的结果,其中雌雄性器官同熟时间短不是主要原因,而其本身较强的自交不亲和性可能是自然结实率低下的关键性内因。由于蓝花丹的专性异交繁育系统使其成功授粉需要传粉媒介,但因引种地与原产地环境存在显著差异,同时开花模式不集中导致的缺乏传粉者完成异花授粉或许是引起异交成功率降低的主要外因。因此,提高蓝花丹的结实率应从克服其自身自交不亲和性以及适当引入安全传媒昆虫入手。
关键词:蓝花丹,繁育系统,传粉生物学,结实率,自交不亲和
吴佩纹,高素萍,张硕,等.蓝花丹结实率低的传粉生物学和繁育系统初探[J].广西植物,2016,36( 1) : 107-113
WU PW,GAO SP,ZHANG S,et al.Pollination biology and the breeding system of Plumbago auriculata low seed-set ratio[J].Guihaia,2016,36( 1) :107-113
异型花柱现象长久以来受到植物学家的关注,包含了二型花柱和三型花柱两种类型,其中二型花柱更为普遍,为最常见的一种雌雄蕊异位( Barrett et al,2000;陈明林等,2010)。异型花柱植物的自交不亲和性被认定为该类型植物的基本特征( Meeus et al,2011; Klein et al,2009),但也有研究显示其具有自交亲和性( Eckert et al,1994)。因异型花柱植物的型内亲和与否和其自然结实率关系密切,故有必要对植物繁育系统中的自交不亲和程度及交配系统作进一步研究(张仁波等,2006)。
蓝花丹( Plumbago auriculata)为多年生常绿灌木,因富含大量的白花丹素具有较高药用价值,且花朵颜色独特,开花周期长,被视为园林景观材料中的新宠。已有学者(张硕等,2014)对蓝花丹的花部形态特征进行了测定,确定其属于二型花柱植物,有长短花柱之分(分别记为L型和S型),初步判定其具有自交不亲和特性。花部形态有着典型的二态性,在花筒口直径,花筒长度,雌蕊高度,雄蕊高度等方面差异极显著,这些特征是蓝花丹对型间互补式雌雄异位的进化适应策略(张硕等,2014)。目前,蓝花丹自然结实率低致使该资源短缺,成为阻碍蓝花丹推广应用的瓶颈问题。植物结实与传粉有着密切联系,传粉是种子植物繁殖的必经阶段(何璐等,2010)。花粉的运动在较大水平上限制了植物个体间的基因流和群体交配方式(黄双全等,2000)。通常成功的传粉需要满足四个条件: ( 1)花粉和花柱都处于成熟状态; ( 2)成熟且有活性的花粉散出; ( 3)拥有足够的传粉媒介及适宜的访花频率保证花粉在柱头的滞落率; ( 4)柱头和花粉之间能成功地相互识别(陈家宽等,1994)。现阶段国内外关于蓝花丹以及所在科属的研究主要集中于观赏特性,生物学特性、药用成分的分析以及组织培养上( Gangopadhyay et al,2008; Iyyakkannu et al,2009; Seiichi et al,2012),Ferrero et al( 2009)初步研究了蓝花丹的传粉特征,但并未从传粉生物学的角度结合其对结实率的影响进行深入探讨,关于其繁育系统的研究也未见报道。因此,蓝花丹是否具有典型的自交不亲和系统,其传粉特征与繁育系统是否与自然结实率低有密切的关系都值得开展深刻的探索。
本实验通过对蓝花丹花粉生活力、柱头可授性、繁育系统及其亲和特性的观察,试图探索蓝花丹自然结实率低的主要原因,为扩大该物种资源、促进其应用给予理论指导和奠定应用基础。
1 材料与方法
1.1材料
为蓝花丹5年生实生苗,原产于南非,其柱头二型分为长花柱型花( L型)和短花柱型花( S型),现种植在四川农业大学成都校区苗圃内。实生苗种子购至泛美,播种以来长势强健。
1.2方法
1.2.1花粉活力的测定用TTC法(张志良,2003)进行花粉活力的测定。把长短柱花型( L型和S型)各10朵进行随机套袋标记,以蓝花丹第1枚花瓣伸展初始记为0 h,分别于0、2、4、6、8、24、36、48、72 h时用解剖针将花粉挑出,置于洁净凹槽载玻片的凹槽内,滴入1~2滴0.5%的TTC( 2,3,5-氯化三苯基四氮唑)溶液,使用镊子把花粉搅拌均匀,于35℃的环境条件下放置15 min,最后于显微镜下观测花粉被染色的情况(赵兴华,2009)。判断活力的标准:染上深红色被认为是活性很强的花粉,略浅的红色表示活性相对低的花粉,未被染上色则表明花粉不存在活性。每张片随机选择3个视野,统计染色花粉数,计算花粉染色率,最后取其平均值为蓝花丹花粉活力百分比,重复3次。花粉活力( %) =染红花粉数(粒) /花粉总数(粒)×100%(朱慧等,2009) 1.2.2柱头可授性的检测柱头可授性用联苯胺-过氧化氢法(刘林德等,2001)检测。随机套袋标记L型花和S型花各10朵,分别于花朵开放后0、2、4、6、8、24、36、48、72 h取下花柱,置于含有1~2滴联苯胺-过氧化氢反应液( 1%联苯胺∶3%过氧化氢∶水= 4∶11∶22,体积比)的凹槽载玻片内。若花柱具有活性,花柱周边的溶液出现蓝色,同时生成较多气泡,相反,没有气泡生成,并且溶液不会变化为蓝色的花柱不具备可授能力。
1.2.3花粉-胚珠比( P/O)的测定按Cruden( 1977)的方法对花粉-胚珠比( P/O)进行测定。把未开裂的长、短柱型花各10朵进行随机采集花药,从每一朵花上获取全部花药,加入1.0 mol·L-1HC1溶液去除花药壁,配成0.5 mL悬浮液(丙三醇∶冰醋酸=1∶3) (徐荣等,2011)。使用微量移液枪移取10 μL至血球计数板,在显微镜下观察花粉染色情况并统计数量,重复操作4次后算其平均值,最后换算出每朵花的花粉粒数(牟勇等,2007)。
取上述实验中花朵的雌蕊放入含0.5 mL 75%酒精的15 mL离心管中保存。用刀片在显微镜下将雌蕊剖开,挑出胚珠并统计数量。按Cruden( 1977)的标准计算每朵花的花粉-胚珠比( P/O),P/O =单花花粉数/单花胚珠数(陆婷等,2010)。
1.2.4杂交指数( OCI)的测定根据Dafni( 1993)的标准,OCI由三个花部特征(单花直径、花药开裂时间和柱头可授性时间是否一致、花药和柱头在空间上的相对位置关系)决定,计算方法如下: ( 1)单花或者花序直径<1 mm时记为0; 1~2 mm时记为1; 2 ~6 mm时记为2,>6 mm时记为3。( 2)雌雄蕊成熟时间的顺序,两者同时成熟或雌蕊先成熟记为0,雄蕊先成熟记为1; ( 3)花药与柱头之间的相对位置,处于相同高度记为0,空间存在分离无法接触记为1。以上三者分值相加为OCI值,判断标准如下:繁育系统在OCI = 0时为闭花受精型( Cleistogamy) ; OCI= 1时为专性自交型( Obligate autogamy) ; OCI = 2时为兼性自交型( Facultative autogamy),有发生异交的几率; OCI= 3时为兼性异交型( Facultative outcrossing ),可自花授粉,某些品种需媒介传粉; OCI= 4时为部分自交亲和,多数品种异交,传粉依赖传粉媒介(杨艳娟等,2012)。
1.2.5人工控制授粉实验将花药未开裂且即将绽放的蓝花丹花蕾进行去雄并标记,根据蓝花丹二型花柱的特征,共进行L×L(自然套袋),S×S(自然套袋),L×L(人工同型同株授粉),S×S(人工同型同株授粉),L×L’(人工同型异株授粉),S×S’(人工同型异株授粉) ( L’:长花柱异株,S’:短花柱异株),不套袋自然传粉,L×S(人工异型花授粉),S×L(人工异型花授粉) 10种处理方法,每种处理供试花朵数30朵,重复3次,定期观察每种处理的结实情况,计算结实率。结种率=成熟种子数/(成熟种子数+败育种子数)×100%(费世民等,2006)。
2 结果与分析
2.1蓝花丹花粉活性的测定结果
由表1可知,长、短型花粉活力在72 h观察期间均出现先上升后下降的趋势。两型花瓣伸展时的花粉生活力L型大于S型,前者为45.5%,后者28.02%。随着时间的延长,其生活力呈上升趋势,但两者达峰值时间不同(前者95.76%,花瓣伸展后8 h;后者94.67%,6 h),即L型要滞后S型2 h。
L、S型花柱的花粉活力在花瓣伸展初始存在一定差异。初始S型花粉的活力略低于L型花粉生活力;花瓣伸展后6~24 h内,两种类型的花粉活力都处于高值阶段,且这一时期是授粉的最佳时期。24 h后,虽长花柱花粉活力还保持20%~80%之间,但此时短花柱花粉的活力已下降明显,48 h后已低于12%,此时授粉已难于成功。
2.2蓝花丹柱头可授性的测定结果
由表1可知,蓝花丹L型柱头在花瓣伸展初始就具有可授性,其可授性随着时间先上升再下降,在花瓣伸展后6 h时达到最大可授性,一直持续至24 h,随后迅速下降,到48 h可授性已十分微弱,直至72 h可授能力完全消失。S型花柱同样在花瓣伸展初始就具有可授性,在花瓣伸展0~2 h内可授性最强,其后迅速下降至6 h时仍具有较弱可授性,但8 h后完全丧失可授能力。
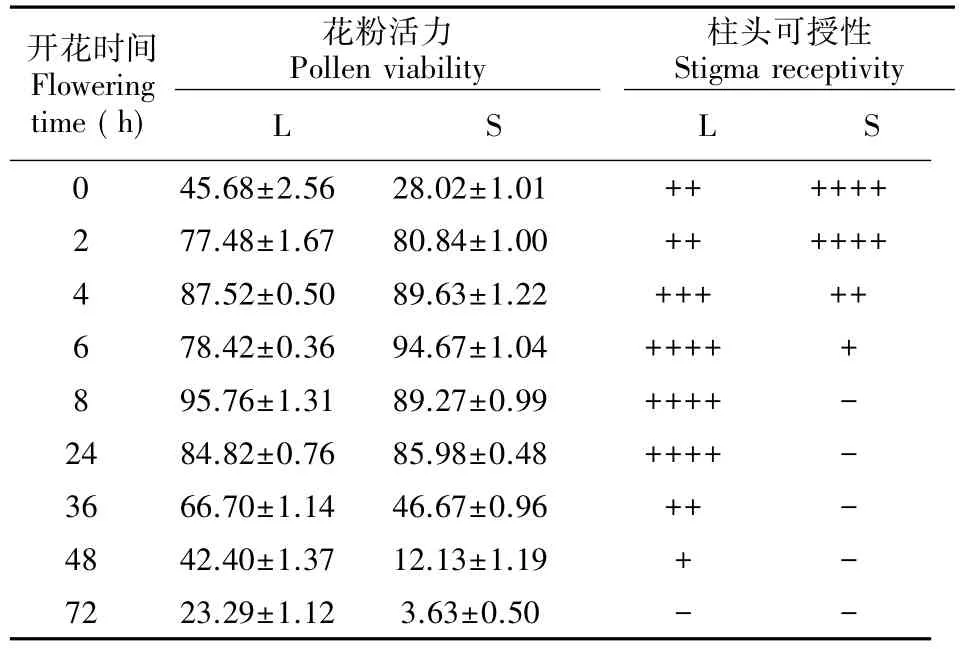
表1 蓝花丹花粉活性、柱头可授性一览表Table 1 Pollen viability and stigma receptivity of Plumbago auriculata

图1 蓝花丹长、短花柱花粉活力期及柱头可授期Fig.1 Long and short style pollen viability period and stigma receptivity period of Plumbago auriculata
尽管蓝花丹两型花粉在花瓣展开后0 h就有较强的可授性,对花柱和花粉活力的关注仍集中在散粉以后。根据观察,L型材料在花瓣展开后5 h左右开始散粉,S型材料在4 h左右开始散粉。由图1可知,由于散粉后,两型柱头上会同时落滞两型花粉,因此,花柱与花粉活力的比较分为四种情况。( 1) L柱头与S花粉活力的比较: S型散粉之后,L型柱头和S型花粉有44 h均具备可授能力; ( 2) S柱头与S花粉活力的比较: S型散粉之后,S型柱头和S型花粉仅有2 h均具备可授能力; ( 3) S柱头与L花粉活力的比较:当L型散粉之后,S型花柱与L型花粉仅有1 h均具备可授能力; ( 4) L柱头与L花粉活力的比较:当L型散粉之后,L型花柱与L型花粉有43 h均具备可授能力。
2.3花粉-胚珠比( P/O)的测定结果
由表2可知,蓝花丹L型花平均单花花粉粒为( 502.00±52.30)粒,单花胚珠数为1,因此可得花粉-胚珠比( P/O)为502.00±52.30; S型花平均单花花粉粒为( 482.70±87.91)粒,单花胚珠数为1,得其花粉-胚珠比( P/O)为482.70±87.91,按照标准,可判断蓝花丹属兼性异交类型。

表2 蓝花丹花粉胚珠比测定结果Table 2 Pollen/ovule ratio of Plumbago auriculata
2.4杂交指数( OCI)的测定结果
按照Dafni( 1993)的方法对蓝花丹进行杂交指数的测量。蓝花丹花冠直径约为26 mm,计为3;雌蕊先于雄蕊成熟,计为0;雌蕊明显高于或者低于雄蕊,计为1,统计得出蓝花丹的杂交指数( OCI)为4,其繁育系统为异交型。这样的结果与用P/O值测定出的繁育系统类型存在一定差异。
2.5人工控制授粉实验
通过表3所示的结果可知,在不去雄自然套袋、人工同型同株自交、人工同型异株自交的处理下,蓝花丹的长花柱型花和短花柱型花结实率都为0;不套袋的自然传粉处理结果显示:长花柱花的自然结实率为( 3.33±2.72) %,短花柱花的自然结实率为( 10.00±4.73) %。而人工辅助异型花杂交的处理中,结实率则分别高达( 90.00±4.71) %( S×L)和( 83.33±2.74) %( L×S)。此结果表明:蓝花丹具有自交不亲和性;人工辅助异花杂交能极大程度上提高该植物的结实率。
3 讨论
根据成功授粉并产生种子的四个条件,本研究从3个层面得出的结果,在一定程度上对自然条件下蓝花丹种子结实量低的原因给予了解释。
首先,在性成熟的时间上,本研究表明蓝花丹L型雌器官与S型雄器官、L型雄器官与L型雌器官成熟时间重叠区域较多,雌雄性器官在成熟时间上没有显著的差异;而S型雌器官与L型雄器官、S型雌器官与S型雄器官成熟时间重叠区域较少,存在一定的生殖隔离。赵志惠( 2012)研究表明蓝花丹群体的整个开花持续期为130 d,个体水平振幅为5,开花同步指数为0.98,呈现一种大量、分散的开放模式。开花不同步性使得整个花期植株周围总有异型的成熟花柱或花药,从而保证花期各个时间段都有充足的花粉供给柱头,在一定程度上缓解了雌雄器官成熟时间上差异带来的生殖隔离。因此,蓝花丹S型雌器官与L型雄器官、S型雌器官与S型雄器官成熟重叠时间短不会影响结实率。
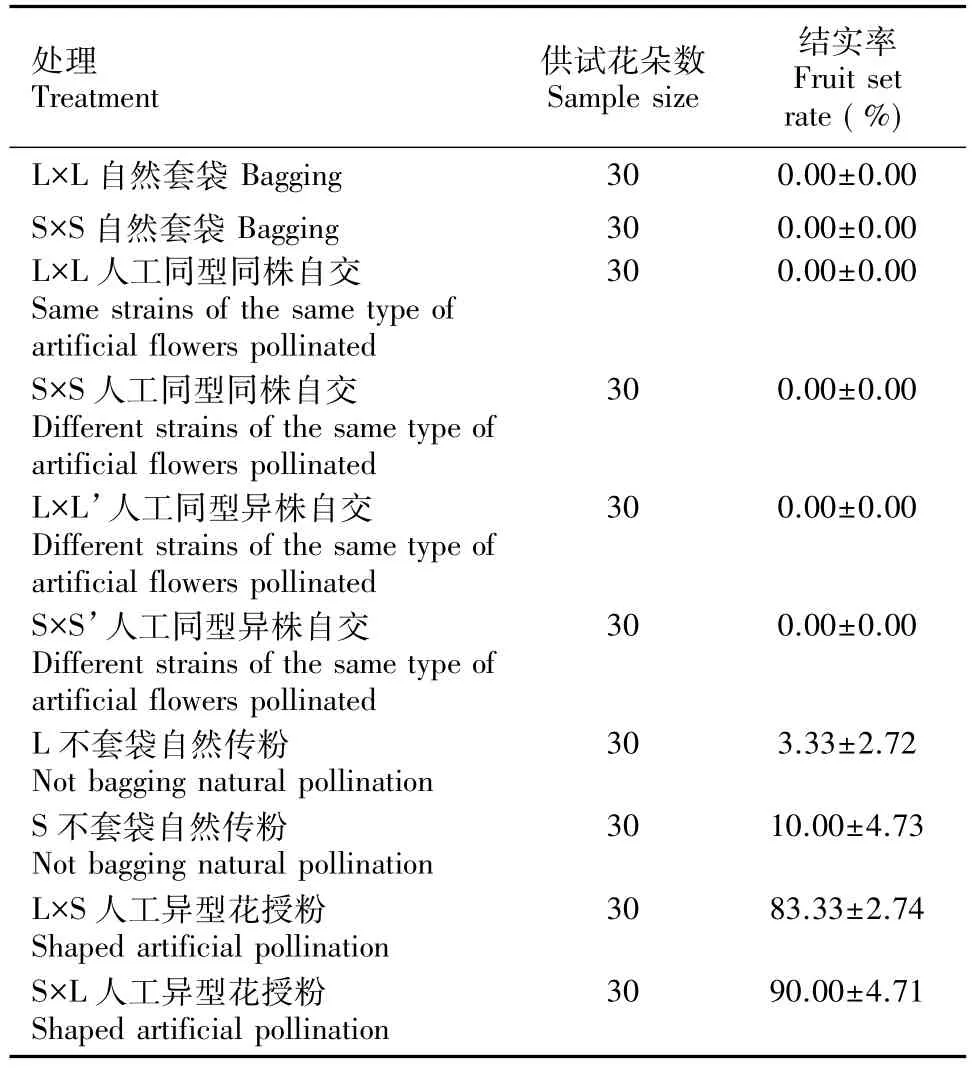
表3 蓝花丹人工辅助授粉的测定结果Table 3 Determination results of artificial pollination experiment on Plumbago auriculata
其次,传粉者作为花粉到柱头的媒介,其到访频率和传粉的有效性显著影响到植物的生殖成功。本研究结果显示,在一致的环境下,人工型间授粉后的结实率远高于自然传粉后的结实率,这说明自然条件下,蓝花丹存在花粉限制。通常情况,蓝花丹昆虫到访率很低,Ferrero et al( 2009)研究表明长喙虻( Philoliche aethiopica)是蓝花丹的重要花粉传播者,达摩蝴蝶( Papilio demodocus)为偶然到访者,且这类传粉者传粉效率不高,由于蓝花丹的原产地为南非,大多在中国被引种到华南、华东、西南等地为绿化树种,其地理位置和气候状况与原产地相比具有一定差异。陆宝麟等( 2003)研究表明长喙虻在中国主要分布于西藏,因此推测出蓝花丹的花粉限制是因为传粉媒介的缺乏。同时由于蓝花丹开花模式具有不集中,持续时间长,日均开花量少的特点,与同期的其他物种相比,开花时对传粉者没有足够的吸引力,也使得传粉者访问频率低,导致花粉落滞率低。由此推测缺乏合适的传粉者及适宜的访花频率使得自然条件下蓝花丹缺乏植株间的花粉交流,这是蓝花丹自然结实率低的重要原因之一。
花粉成功到达柱头后,柱头与花粉是否亲和成为限制植物结实的关键。植物繁育系统的测定中常用到P/O、OCI测定和人工控制授粉(何平,2005;周红军等,2003;张仁波等,2006;何亚平等,2003;赖家业等,2008;张文标等,2009,秦垦等,2009)。本研究中,P/O结果显示蓝花丹为兼性异交,而OCI及人工控制授粉结果显示其为专性异交。OCI检测是从植物形态适应虫媒传粉出发,相对于P/O正确性更高(张健等,2009)。因此两者结果出现差异时,通常取OCI值作为最后判定植物繁育系统的依据。人工控制授粉试验则排除了环境因素和授粉过程的差异,直接关注花柱和花粉的识别对结实率的影响,为三种方法中最为有效的检测方式。本研究综合三种测定的方法结果,判定蓝花丹为专性异交。
人工控制授粉中,长花柱型花和短花柱型花,不论自然套袋、人工同型同株自交、人工同型异株自交均不会结实。而型间人工授粉后结实率高达( 83.33 ±2.74) %和( 90.00±4.71) %。型间和型内结实率的巨大悬殊说明自交不亲和性在蓝花丹的生殖上起着决定性作用。一般认为,在二型花柱植物中,型间的交配是合法的,而型内交配则表现出不亲和性,如安徽羽叶报春等异型花授粉的结籽数均明显高于自花授粉和同型异花授粉结籽数(邵剑文等,2011)。艳丽耳草长/短花柱型花型间异交花粉管生长形态正常,而长/短花柱型花在自交及型内自交下均表现为不亲和(刘欣欣等,2012)。作为专性异交植物,因为强烈的自交不亲和特性而在自然条件下对花粉有着甄别作用,要想授粉成功要求蓝花丹种群有合适的型比,以及型间保持适宜的距离便于生殖。在缺乏某一型的种群中或种群中型间距离过长时,就会因为自交不亲和特性而导致较低的自然结实率。白花丹属的紫花丹也有相似现象(吴征镒,2004)。
综上所述,传粉者的缺乏和自交不亲和的特性共同影响蓝花丹的自然结实率。其中,雌雄性器官成熟期相遇时间短不是主要原因;缺乏有效的传粉者可能是重要的外因,而较强的自交不亲和性则有可能是其结实率低下的关键性内因。因此提高外来优良物种蓝花丹的结实率,要从引入安全的传粉昆虫和克服自交不亲和性入手。
参考文献:
BARRETT SCH,JESSON LK,BAKER AM,2000.The evolution and function of stylar polymorphisms in flowering plants[J].Ann Bot,85: 253-265.
CHEN JK,YANG J,1994.Plant evolutionary biology[M].Wuhan: Wuhan University Press: 232-280.[陈家宽,杨继,1994.植物进化生物学[M].武汉:武汉大学出版社: 232-280.]
CHEN ML,YOU YL,ZHANG XP,2010.Advances in the research of heterostyly[J].Acta Pratacul Sin,19( 1) : 226-239.[陈明林,游亚莉,张小平,2010.花柱异型研究进展[J].草业学报,19( 1) : 226-239.]
CRUDEN RW,1977.Pollen ovule ratios: A conservative indicator of breeding systems in flowering plants[J].Evolution,31( 1) : 32-46.
DAFNI,1993.Pollination ecology: a practical approach,Oxford University Press Oxford[J].J Evol Biol,6( 5) : 776.
ECKERT CG,BARRETT SCH,1994.Tristyly,self-compatibility and floral variation in Decodon verticillatus( Lythraceae)[J].Biol J Linn Soc,53( 1) : 1-30.
FEI SM,PENG ZH,YANG DS,et al,2006.Seed rain and seed bank of Castanopsis delavayi populations in mountainous area of Southwest Sichuan[J].Sci Silv Sin,42( 2) : 49-54.[费世民,彭镇华,杨冬生,等,2006.川西南山地高山栲种群种子雨和地表种子库研究[J].林业科学,42( 2) : 49-54.]
FERRERO V,DE VEGA V,STAFFORD GI,et al,2009.Heterostyly and pollinators in Plumbago auriculata ( Plumbaginaceae) [J].S Afr J Bot,75: 778-784.
FERRERO V,VEGA C,STAFFORD GI,et al,2009.Heterostyly and pollinators in Plumbago auriculata ( Plumbaginaceae)[J].S Afr J Bot,75( 4) : 778-784.
GANGOPADHYAY M,SIRCAR D,MITRA A,2008.Hairy root culture of Plumbago indica as a potential source for plumbagin [J].Biol Pant,52( 3) : 533-537.
HE L,YU H,FAN YH,et al,2010.Research progress in plant reproductive biology[J].J Mount Agric Biol,29( 5) : 456-460.[何璐,虞泓,范源洪,等,2010.植物繁殖生物学研究进展[J].山地农业生物学报,29( 5) : 456-460.]
HE P,2005.The rare and endangered plant protection biology [M].Chongqing: Southwest Normal University Press: 207-240.[何平,2005.珍稀濒危植物保护生物学[M].重庆:西南师范大学出版社: 207-240.]
HUANG SQ,GUO YH,2000.Advances in pollination biology[J].Chin Sci Bull,45( 3) : 225-237.[黄双全,郭友好,2000.传粉生物学的研究进展[J].科学通报,45( 3) : 225-237.]
HE YP,LIU JQ,2003.A review on recent advances in the studies of plant breeding system[J].J Plant Ecol,27( 2) : 151-163.[何亚平,刘建全,2003.植物繁育系统研究的最新进展和评述[J].植物生态学报,27( 2) : 151-163.]
IYYAKKANNU S,BYOUNG RJ,2009.Induction and establishment of adventitious and hairy root cultures of Plumbago zeylanica L.[J].Afr J Biotechnol,8( 20) : 5 294-5 300.
KLEIN DE,FREITAS L,2009.Self-incompatibility in a distylous of species of Rubiaceae: is there a single incompatibility respouse of the morphs[J].Sex Plant Reprod,22( 3) : 121-131.
LAI JY,SHI HM,PAN CL,et al,2008.Pollination biology of rare and endangered species Malania oleifera Chun et Lee[J].J Beijing For Univ,20( 2) : 59-63.[赖家业,石海明,潘春柳,等,2008.珍稀濒危植物蒜头果传粉生物学研究[J].北京林业大学学报,20( 2) : 59-63.]
LIU LD,ZHANG HJ,2001.Acanthopanax pollen viability and stigma receptivity study[J].Plant Res,21( 3) : 375-379.[刘林德,张洪军,2001.刺五加花粉活力和柱头可授性的研究[J].植物研究,21( 3) : 375-379.]
LIU XX,WU XQ,ZHANG DX,2012.Distyly and heteromorphic self-incompatibility of Hedyotis pulcherrima ( Rubiaceae)[J].Biodiversity,20( 3) : 337-347.[刘欣欣,吴小琴,张奠湘,2012.艳丽耳草的二型花柱及异型自交不亲和系统[J].生物多样性,20( 3) : 337-347.]
LU BL,WU HY,2003.China classification and identification of important medical insects[M].Zhengzhou: Henan Science Technology Press: 285-286.[陆宝麟,吴厚永,2003.中国重要医学昆虫分类与鉴别[M].郑州:河南科学技术出版社: 285-286.]
LU T,LUO SP,MENG M,et al,2010.Flowering biological characteristics of Zizyphus jujuba Mill.[J].Acta Bot Boreal-Occident Sin,30( 8) :1 589-1 594.[陆婷,罗淑萍,蒙敏,等,2010.灰枣开花生物学特性研究[J].西北植物学报,30( 8) :1 589-1 594.]
MEEUS S,JACAUEMYN H,HONNAY O,et al,2011.Self-incompatibility and pollen limitation in the rare tristylous endemic Hugonia serrata on La Reunion Island[J].Plant Syst&Evol,292( 34) : 143-151.
MOU Y,ZHANG YH,LOU AR,2007.A preliminary study on floral syndroma and breeding system of the rare plant Rhodiola dumulosa[J].J Plant Ecol,31( 3) 528-535.[牟勇,张云红,娄安如,2007.稀有植物小丛红景天花部综合特征与繁育系统[J].植物生态学报,31( 3) 528-535.]
QIN K,WANG B,JIAO EN,et al,2009.Preliminary study on the breeding system of Lycium barbarum[J].Guihaia,29( 5) : 587-591.[秦垦,王兵,焦恩宁,等,2009.宁夏枸杞繁育系统初步研究[J].广西植物,29( 5) : 587-591.]
SEIICHI S,WARAPORN P,BENYAKAN P,2012.Modulation of plumbagin production in Plumbago zeylanica using a singlechain variable fragment antibody against plumbagin[J].Plant Cell Rep,31: 103-110.
SHAO JW,ZHANG WJ,ZHANG XP,2011.Reproductive characteristics and adaptive evolution of pin and thrum flowers in endangered species Primula merrilliana[J].J Ecol,31( 21) : 6 410-6 419.[邵剑文,张文娟,张小平,2011.濒危植物安徽羽叶报春两种花型的繁育特性及其适应进化[J].生态学报,31( 21) : 6 410-6 419.]
WU ZY,2004.Flora of China[M].Beijing: Science Press,60 ( 1) : 5.[吴征镒,2004.中国植物志[M].北京:科学出版社,60( 1) : 5.]
XU R,WANG X,CHEN J,et al,2011.Study on breeding system and insect pollinator species of cultivated Cistanche deserticola [J].Chin Pharma J,46( 14) : 1 049-1 052.[徐荣,王霞,C陈君,等,2011.栽培肉苁蓉的繁育系统和传粉昆虫种类研究[J].中国药学杂志,46( 14) : 1 049-1 052.]
YANG YJ,XIE SQ,MENG ZG,2012.Pollination ecology of Coptis teeta Wall.an endangered medicinal plant[J].Acta Bot Boreal-Occident Sin,32( 7) : 1 372-1 375.[杨艳娟,谢世清,孟珍贵,2012.濒危药用植物云南黄连传粉生态学研究[J].西北植物学报,32( 7) : 1 372-1 375.]
ZHANG J,YANG FJ,LIU S,et al,2009.Breeding system study in a stragalusm embranaceus( Ficsh) Bunge[J].Ginseng Res,( 3) : 17-20.[张健,杨发君,刘顺,等,2009.膜荚黄芪繁殖系统的研究[J].人参研究,( 3) : 17-20.]
ZHANG RB,DOU QL,HE P,et al,2006.Study on the breeding system of the endangered plant Euonymus chloranthoides Yang [J].Guihaia,26( 3) : 308-312.[张仁波,窦全丽,何平,等,2006.濒危植物缙云卫矛繁育系统研究[J].广西植物,26 ( 3) : 308-312.]
ZHAO S(张硕),GAO SP(高素萍),2014.Distyly and selfing incompatibility of Plumbago auriculata[J].Guihaia,34( 6) : 747-753.[张硕,高素萍,2014.蓝花丹的花部形态二态性及自交不亲和特性[J].广西植物,34( 6) : 747-753.]
ZHANG WB,JIN ZX,2009.Floral syndrome and breeding system of endangered plantsinocalycanthus chinensis[J].J Zhejiang Univ Med Sci,36( 2) : 204-210.[张文标,金则新,2009.濒危植物夏腊梅花部综合特征与繁育系统[J].浙江大学学报,36( 2) : 204-210.]
ZHANG XH,ZHANG DX,YIN DS,et al,2009.Study on vitality measurement and storing methods of Asiatic Lily pollen[J].Northern Hortic,( 2) : 172-175.[赵兴华,张道旭,印东生,等,2009.百合花粉生活力测定和贮藏方法研究[J].北方园艺,( 2) : 172-175.]
ZHANG ZL,2003.Plant physiology experimental guidance[M].Beijing: Higher Education Press: 224-225.[张志良,2003.植物生理学实验导[M].北京:高等教育出版社: 224-225.]
ZHAO ZH,2012.A preliminary study of Plumbago auriculata biological characteristics[D].Ya’an: Sichuan Agric Univ.[赵志惠,2012.蓝花丹生物学特性初步研究[D].雅安:四川农业大学.]
ZHU H,MA RJ,2009.Preliminary studies on breeding system of invasive weed Ipomoea cairica L.[J].J NW A&F Univ,37 ( 5) : 211-216.[朱慧,马瑞君,2009.入侵杂草五爪金龙有性繁育系统的初步研究[J].西北农林科技大学学报,37( 5) : 211-216.]
ZHOU HJ,TANG L,MA X,et al,2003.A preliminary study of soil Radix pollination biology and mating systems[J].J Beijing For Univ,39( 5) : 669-673.[周红军,唐亮,马香,等,2003.土麦冬传粉生物和交配系统的初步研究[J].北京师范大学学报,39( 5) : 669-673.]
(上接第60页 Continue from page 60 )
WEBER A,1976.Beiträge zur morphologie und systematik der Klugieae und Loxonieae ( Gesneriaceae).III: Whytockia als morphologische und phylogenetische Ausgangsform von Monophyllaea[J].Beitr Biol Pflanzen,52: 183-205.
WEBER A,1982.Contributions to the morphology and systematics of Klugieae and Loxonieae ( Gesneriaceae).IX.The genus Whytockia[J].Notes Roy Bot Gard Edinburgh,40: 113-121.
WEBER A,BURTT BL,1998.Remodelling of Didymocarpus and associated genera ( Gesneriaceae)[M].Beitr Biol Pflanzen,70: 293-363.
WEBER A,2004.Gesneriaceae[M]/ / KUBITZKI K,KADEREIT JW ( eds).The families and genera of vascular plants.Vol.7.Flowering plants.Dicotyledons.Lamiales ( except Acanthaceae,including Avicenniaceae).Berlin/Heidelberg: Springer: 63-158.
WEBER A,Middleton DJ,Forrest A,et al,2011a.Molecular systematics and remodelling of Chirita and associated genera ( Gesneriaceae)[J].Taxon,60: 767-790.
WEBER A,WEI YG,PUGLISI C,et al,2011b.A new definition of the genus Petrocodon ( Gesneriaceae)[J].Phytotaxa,23: 49-67.WEBER A,WEI YG,SONTAG S,et al,2011c.Inclusion of Metabriggsia into Hemiboea ( Gesneriaceae)[J].Phytotaxa,23: 37-48.
WEBER A,CLARK JL,MÖLLER M,2013.A new formal classification of Gesneriaceae[J].Selbyana,31( 2) 68-94.
WEI YG,2004.Paralagarosolen Y.G.Wei,a new genus of the Gesneriaceae from Guangxi,China[J].Acta Phytotax Sin,42: 528-532.
WEI YG,WEN F,CHEN WH,et al,2010.Litostigma,a new genus from China: a morphological link between basal and derived Didymocarpoid Gesneriaceae[J].Edinburgh J Bot,67( 1) : 161-184.
WEN F,LIANG GY,WEI YG,2012.Petrocodon lancifolius ( Gesneriaceae),a new species endemic to a central subtropical zone of Guizhou Province,China[J].Phytotaxa,49: 45-49.
WEN F,MACIEJEWSKI S,HE XQ,et al,2015a.Briggsia leiophylla,a new species of Gesneriaceae from southern Guizhou,China[J].Phytotaxa,202 ( 1) : 51-56.
WENF,WEI YG,MÖLLER M,2015b.Glabrella leiophylla ( Gesneriaceae),a new combination for a former Briggsia species from Guizhou,China[J].Phytotaxa,218 ( 2) : 193-194.
Wood D,1974.A revision of Chirita ( Gesneriaceae)[J].Notes Roy Bot Gard Edinburgh,33: 123-205.
XU WB,MENG T,ZHANG Q,et al,2014.Petrocodon ( Gesneriaceae) in the limestone karsts of Guangxi,China: Three new species and a new combination based on morphological and molecular evidence[J].Syst Bot,39( 3) : 965-974.
XU WB,ZHANG Q,WEN F,et al,2012.Nine new combinations and one new name of Primulina ( Gesneriaceae) from South China[J].Phytotaxa,64: 1-8.
XU ZR,BURTT BL,SKOG LE,et al,2008.A revision of Paraboea ( Gesneriaceae)[J].Edinburgh J Bot,65( 2) : 161-347.
YANG CZ,CAI DL,WEN F,2015.Oreocharis striata ( Gesneriaceae),a new species from Fujian,China[J].Ann Bot Fenn,52: 369-372.
YAO TL,2012.A taxonomic revision of Loxocarpus ( Gesneriaceae) [D].MSc thesis,Faculty of Science,Universiti Malaya,Kuala Lumpur.
YU,XL,LI,M,ZHOU JJ,et al,2015.Petrocodon hunanensis ( Gesneriaceae),a new species identified by both morphological and molecular evidence from limestone area in Hunan,China [J].Phytotaxa,195( 1) : 65-72.
Pollination biology and the breeding system of Plumbago auriculata low seed-setratio
WU Pei-Wen1,GAO Su-Ping2*,ZHANG Shuo1,ZHAO Zhi-Hui1
( 1.College of Landscape Architecture,Sichuan Agricultural Uniersity,Chengdu 611130,China; 2.Landscape Research Institute of Sichuan Agricultural University,Chengdu 611130,China )
Abstract:In order to explain the main reasons for the low natural seed-set ratio of Plumbago auriculata,which can contribute to the large-scale production of P.auriculata and also help to get rid of the situation that the seeds over-rely on the import from abroad at the same time.We attempted to measured the dynamic changes of the pollen viability by the method of TTC and the stigma receptivity and the method of Benzidine-H2O2in two ways,which respectively focused more on the pollination biology and the breeding system.And we also estimated the type of the breeding system pollenovule ratio ( P/O),and verified the out-crossing index ( OCI) by the pollination experiments.The results of the experiment were as follows: ( 1) About P.auriculata,the maturity time of the L-type female organs and the S-type male organs,and also that of the L-type female organs and the L-type male organs overlaped a lot,there was no obvious difference in the maturity time between the female and the male organs.On the other hand,the maturity time of the S-type female organs and the L-type male organs,and also that of the S-type female organs and the S-type male organs overlapedbook=108,ebook=113a little,but the continuous blossoming pattern of P.auriculata reduced reproductive isolation caused by the different maturity time of the male and the female sexual organs.( 2) The P/O of L-type was 502±52.30,the P/O of S-type 482.7± 87.91,both the out-crossing index of the L-type and that of the S-type are 4.The result of the manual-controlled pollination experiments suggested that the breeding system of P.auriculata was obligate outcrossing and it had the feature of the heteromorphic incompatibility.To sum up,the reasons for the low natural seed-set ratio of P.auriculata were the combination of both internal and external factors.Among these factors,the short same maturity time of the male and the female sexual organs was not the main reason,while the strong self-incompatibility of P.auriculata itself might be the key internal factor to lead to the low natural seed-set ratio.Due to the breeding system of the obligate outcrossing,it was quite necessary for P.auriculata to had the pollination medium to lead to the successful pollination.However,the introduction environment and the original environment were distinctly different,in addition,the mode of the scattered blooming led to the cross-pollinator,which might be the key external factor of low success rate of outcrossing.In a conclusion,the improvement of the natural seed-set ratio of P.auriculata should mainly focus on overcoming the incompatibility of P.auriculata itself,and also properly bringing in some insects as the safe medium.This would provide the theoretical guidance and the application foundation for the expansion of this kind of species resource and also the acceleration of the promotion and the application.
Key words:Plumbago auriculata,breeding system,pollination biology,seed-set ratio,self-incompatibility
*通讯作者:高素萍,博士,教授,主要从事观赏植物生理生态研究,( E-mail) supingg@ hotmail.com。
作者简介:吴佩纹( 1991-)女,四川广安人,在读硕士生,主要从事蓝花丹繁殖研究,( E-mail) : 76794231@ qq.com。
基金项目:四川省科技支撑计划项目( 2012FZ0083)[Supported by Science and Technology Support Program of Sichuan( 2012FZ0083)]。
收稿日期:2014-09-29修回日期: 2014-12-27
DOI:10.11931/guihaia.gxzw201409056
中图分类号:Q945.5
文献标识码:A
文章编号:1000-3142( 2016) 01-0107-07
