如皋黄鸡开产性状加性遗传及母体遗传效应分析
2016-03-18沈曼曼窦套存胡玉萍王克华
郭 军, 曲 亮, 沈曼曼, 窦套存, 胡玉萍, 王克华
(中国农业科学院 家禽研究所, 江苏 扬州 225125)
如皋黄鸡开产性状加性遗传及母体遗传效应分析
郭军, 曲亮, 沈曼曼, 窦套存, 胡玉萍, 王克华
(中国农业科学院 家禽研究所, 江苏 扬州 225125)
摘要:【目的】以动物模型分析加性遗传、母体遗传对如皋黄鸡开产性状的影响,并计算开产性状遗传参数。【方法】收集如皋黄鸡2009—2012年生产数据,包括10 454个动物个体、31 362条生产记录。依据是否考量母体效应,设计4种统计模型。利用WOMBAT软件以平均信息约束最大似然法(AIREML)剖分表型方差,以似然比统计值检验模型适用性。以单性状模型分析遗传力,以多性状模型分析遗传相关系数和表型相关系数。【结果】母体环境对开产日龄有显著的影响,模型II为开产日龄最适模型,开产日龄遗传力为0.28±0.02;母体遗传对平均开产蛋质量影响显著,模型III适合计算开产蛋质量遗传力,开产蛋质量加性遗传力为0.23±0.02、母体遗传力为0.03±0.01;母体遗传和母体环境对开产体质量有显著影响,开产体质量加性遗传力为0.34±0.03、母体遗传力为0.04±0.01,模型IV为开产体质量最适模型。如皋黄鸡开产蛋质量、开产体质量加性遗传与母体遗传存在负相关。开产体质量、开产蛋质量与开产日龄之间的遗传相关系数分别为0.36±0.07、0.33±0.07,开产蛋质量与开产体质量之间的遗传相关系数为0.65±0.05。【结论】如皋黄鸡开产日龄、开产体质量、开产蛋质量等开产性状均受母体效应影响,且遗传力较低。
关键词:如皋黄鸡; 遗传力; 繁殖性状; 母体效应; 动物模型
遗传力是预测群体选择反应、估计个体育种值的关键参数,当前遗传力研究热点之一是分析母体效应对加性遗传的影响。开产性状是蛋鸡育种研究热点之一,不仅反映性成熟状况,还与早期产蛋性能密切相关。如皋黄鸡主要分布于长江下游流域,属于肉蛋兼用型品种,具有开产早、蛋黄比例高、肉质优良的特性。因此,系统研究如皋黄鸡繁殖性能遗传力,对于如皋黄鸡开发利用具有重要意义。近年来,生态学、进化生物学以及基因组学研究表明鸟类生长、生殖性状受母体效应影响。据Mousseau等[1]研究,雌鸟可依据环境状况改变后代数量和初生体质量。雌鸟可以改变禽蛋中非DNA物质含量,从而影响后代的生长发育。例如,蛋黄中生殖激素、类胡萝卜素以及维生素影响后代生长、免疫、行为、形态以及生活力,母体依据环境条件调节这些物质在禽蛋中的含量[2-3]。据报道鸡基因组存在母体遗传现象[4-5]。鸡1号染色体蛋质量、蛋品质QTL只是母系来源的等位基因表达。鸡3号染色体也存在基因组印记,屠宰性状以及蛋质量QTL是单个等位基因表达。研究表明,蛋鸡[6]、肉鸡[7]、火鸡[8]、 鸭子[9]、鹅[10]、 鹌鹑[11]体质量性状遗传力受母体效应影响。综上所述,多个研究团队应用不同的研究手段证明禽蛋在介导鸟类母体效应中发挥重要作用。然而,目前的家禽开产性状遗传力计算中很少考虑母体效应,因而可能导致遗传力估值过高。本研究以如皋黄鸡选育群体为材料,收集2009—2012年生产数据,检验加性遗传、母体效应对如皋黄鸡繁殖性状的影响并估计开产日龄、开产蛋质量及开产体质量遗传力。研究结果将为如皋黄鸡选育提供支持,为其他地方鸡种遗传参数研究提供参考。
1材料与方法
1.1试验动物
试验数据集来自扬州翔龙禽业有限公司。如皋黄鸡产蛋期饲养于指定鸡舍,育成期光照条件为8 h光照∶16 h黑暗,产蛋期光照条件为16 h光照∶8 h黑暗。全程笼养,常规蛋鸡饲料。试验群体测定开产日龄、开产蛋质量和开产体质量数据,其中开产蛋质量是最初3枚蛋平均值。选取2009—2012年连续4代有系谱记录和生产数据的资料,原始数据集由来自10 454只鸡的31 362条生产记录组成,经分析整理,去除离群值,其中,开产日龄、开产蛋质量、开产体质量的记录数分别为10 352、10 335、10 378条,父系数为242个,开产日龄、开产蛋质量数据来自1 494个母系,开产体质量数据来自1 493个母系。数据统计信息详见表1。
表1如皋黄鸡开产性状数据描述
Tab.1Date description statistics of reproduction traits in Rugao yellow chickens

项目开产日龄/d开产蛋质量/g开产体质量/g最大值18144.001948.00最小值12322.00857.00平均值±标准差150.69±9.3432.99±3.19 1350.93±161.38CV/%6.209.6711.95
1.2统计模型
首先,应用SAS GLM分析表型方差,以生产记录包含的年代、批次、笼位信息对表型值进行方差分析,确定进入固定效应的因素。其次,应用WOMBAT软件,采用平均信息最大约束似然法,分析各性状(协)方差组分以及遗传力[12]。以是否包含母体遗传、母体环境效应为区分标准,建立4个动物模型:
模型I(model I):Y=Xb+Zaa+e,
模型II(modelII):Y=Xb+Zaa+Zcc+e,
模型III(modelIII):Y=Xb+Zaa+Zmm+e,
模型IV(modelIV):Y=Xb+Zaa+Zcc+Zmm+e,
式中:Y是所有个体性状表型值向量;a、b、c、m、e分别是加性遗传效应、固定效应、母体环境效应、母体遗传效应以及残差效应的向量。X、Za、Zm、Zc分别是固定效应、加性遗传效应、母体遗传效应以及母体环境效应的指定矩阵。最后,以似然比比较各性状适用模型[13-14]。似然比检验统计量为:
-2/log(e)×[logLmax|modelⅠ-logLmax|modelⅡ]=
-4.605 2×[logLmax|modelⅠ-logLmax|modelⅡ],
2结果与分析
2.1影响开产性状表型方差的非遗传因素
表型值具有一定的弹性空间,其中环境是影响性状表现的重要因素。如表2所示,如皋黄鸡开产性状按环境因素分组后平均值和标准差存在显著差异。批次指定依照时间顺序,距离当下最近的为第1批,最远的是第10批。批次之间,饲养管理、气温、湿度条件存在差异,因此各批次开产性状的平均值表现显著差异。开产早的批次,平均开产蛋质量较小。1~3批是2012年7月13日—2012年8月13日出雏。4~6批是2011年8月6日—2011年9月8日出雏,批次之间时间间隔较短,育雏、育成管理较好,因而开产体质量差异不显著,且平均值较大。7~9批是2010年6月30日—2010年8月16日出雏,批次之间时间间隔长,所以批次间开产体质量差异显著。 综合考虑,(协)方差组分分析时将批次因素列入固定效应。
表2不同批次如皋黄鸡开产性状平均值和标准差1)
Tab.2Means and standard deviations of reproduction traits of Rugao yellow chickens from different batches
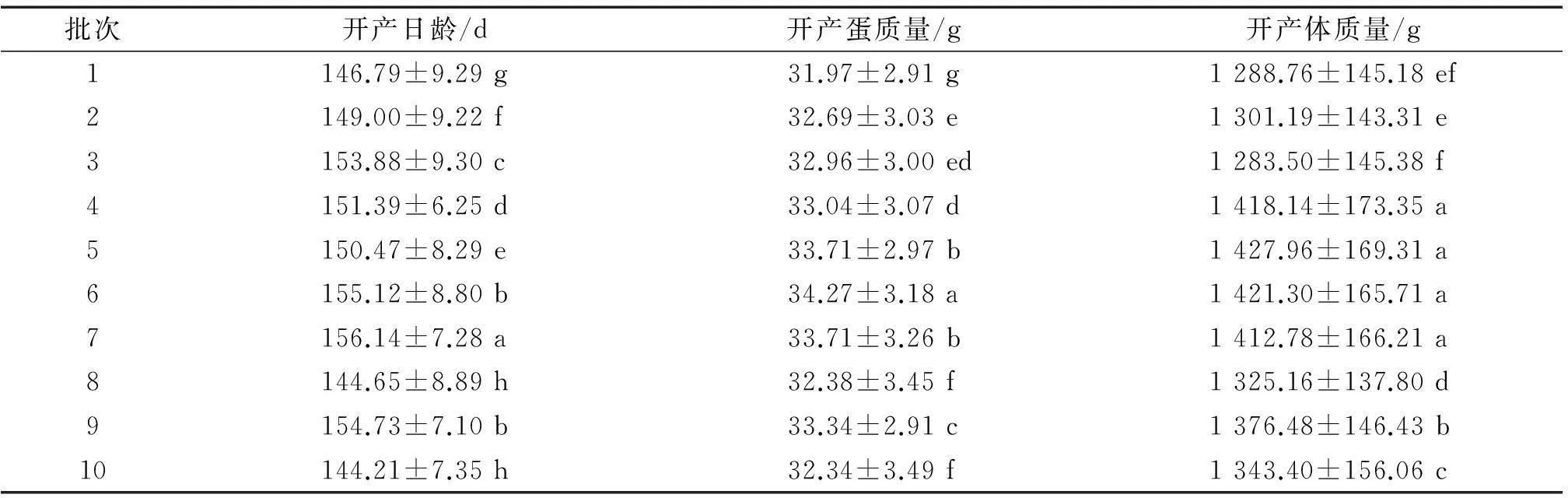
批次开产日龄/d开产蛋质量/g开产体质量/g1146.79±9.29g31.97±2.91g1288.76±145.18ef2149.00±9.22f32.69±3.03e1301.19±143.31e3153.88±9.30c32.96±3.00ed1283.50±145.38f4151.39±6.25d33.04±3.07d1418.14±173.35a5150.47±8.29e33.71±2.97b1427.96±169.31a6155.12±8.80b34.27±3.18a1421.30±165.71a7156.14±7.28a33.71±3.26b1412.78±166.21a8144.65±8.89h32.38±3.45f1325.16±137.80d9154.73±7.10b33.34±2.91c1376.48±146.43b10144.21±7.35h32.34±3.49f1343.40±156.06c
1)同列数据后凡是有一个相同小写字母者,表示在0.05水平上差异不显著(单因素方差分析SNK法)。
2.2开产性状遗传参数
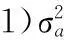
表3如皋黄鸡开产性状不同模型的方差组分和遗传力1)
Tab.3Variance components and heritabilities of reproduction traits of Rugao yellow chickens estimated with different models
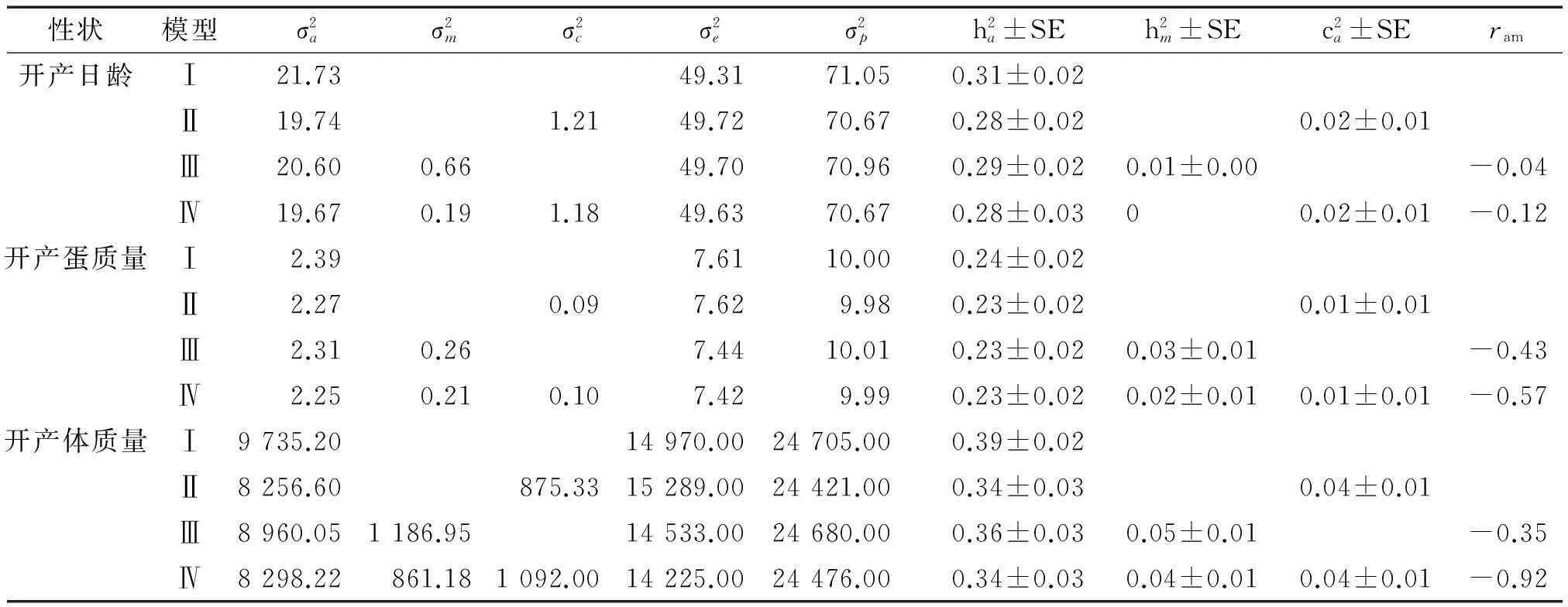
性状模型σ2aσ2mσ2cσ2eσ2ph2a±SEh2m±SEc2a±SEram开产日龄Ⅰ21.7349.3171.050.31±0.02Ⅱ19.741.2149.7270.670.28±0.020.02±0.01Ⅲ20.600.6649.7070.960.29±0.020.01±0.00-0.04Ⅳ19.670.191.1849.6370.670.28±0.0300.02±0.01-0.12开产蛋质量Ⅰ2.397.6110.000.24±0.02Ⅱ2.270.097.629.980.23±0.020.01±0.01Ⅲ2.310.267.4410.010.23±0.020.03±0.01-0.43Ⅳ2.250.210.107.429.990.23±0.020.02±0.010.01±0.01-0.57开产体质量Ⅰ9735.2014970.0024705.000.39±0.02Ⅱ8256.60875.3315289.0024421.000.34±0.030.04±0.01Ⅲ8960.051186.9514533.0024680.000.36±0.030.05±0.01-0.35Ⅳ8298.22861.181092.0014225.0024476.000.34±0.030.04±0.010.04±0.01-0.92

如皋黄鸡开产日龄加性遗传效应与母体遗传效应之间存在较弱负相关(ram为-0.04~-0.12,见表3),表明选育提高开产日龄加性遗传效应对母体遗传效应几乎没有影响。开产蛋质量的ram为-0.43~-0.57,表明选育提高开产蛋质量加性遗传将降低母体遗传效应。对于开产体质量而言,加性遗传效应与母体遗传效应相关系数为-0.35~-0.92。
以多性状模型计算的遗传力与表3列出的相差不大。由表4可知,表型相关系数小于遗传相关系数,表明受环境相关影响。
表4开产性状之间遗传相关(左下)和表型相关(右上)1)
Tab.4Estimated values of genetic (lower left) and phenotypic (upper right) correlations between traits at the first egg

性状开产日龄开产体质量开产蛋质量开产日龄0.28±0.031)0.19±0.010.23±0.01开产体质量0.36±0.070.35±0.030.31±0.01开产蛋质量0.33±0.070.65±0.050.23±0.02
1) 对角线上标示下划线数值为遗传力。
2.3动物模型比较
表5遗传评估模型最大似然值对数
Tab.5The maximum logarithmic likelihood value for each model
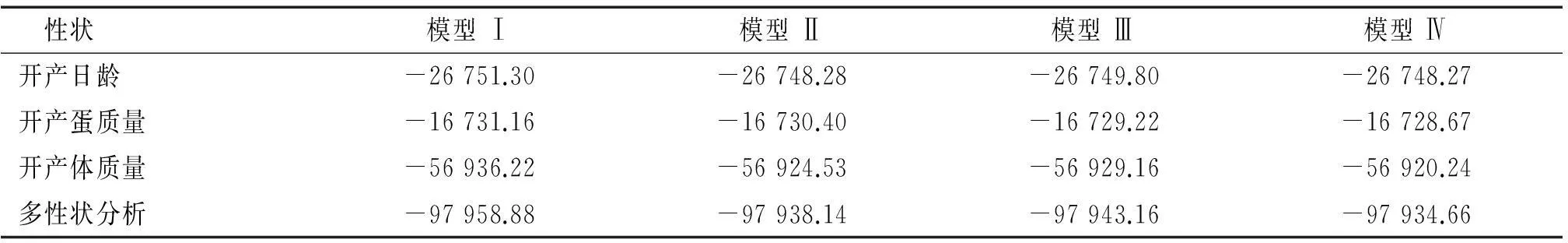
性状模型Ⅰ模型Ⅱ模型Ⅲ模型Ⅳ开产日龄-26751.30-26748.28-26749.80-26748.27开产蛋质量-16731.16-16730.40-16729.22-16728.67开产体质量-56936.22-56924.53-56929.16-56920.24多性状分析-97958.88-97938.14-97943.16-97934.66
表6模型之间似然比值及2检验结果1)
Tab.6Likelihood ratios and2 test results among different models
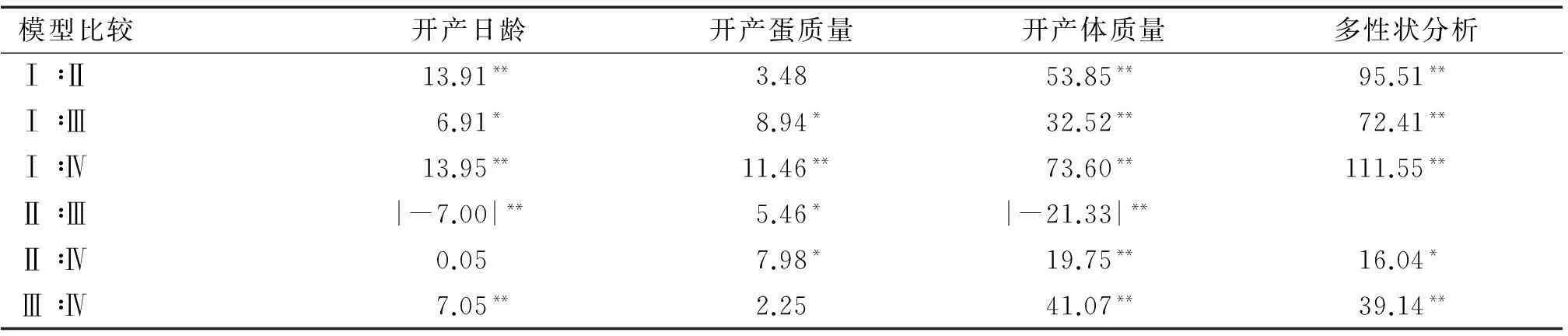
模型比较开产日龄开产蛋质量开产体质量多性状分析Ⅰ∶Ⅱ13.91**3.4853.85**95.51**Ⅰ∶Ⅲ6.91*8.94*32.52**72.41**Ⅰ∶Ⅳ13.95**11.46**73.60**111.55**Ⅱ∶Ⅲ|-7.00|**5.46*|-21.33|**Ⅱ∶Ⅳ0.057.98*19.75**16.04*Ⅲ∶Ⅳ7.05**2.2541.07**39.14**
1)*表示0.05水平差异显著;**表示0.01水平差异显著。
3讨论与结论
3.1开产性状遗传力
对于开产日龄,2000年之前的研究认为其遗传力较高,范围从0.31~0.48[15-17]。近年,以亚洲地方鸡种为试材遗传分析表明开产日龄属于低遗传力性状,其值从0.12~0.32[18-20]。本研究中,如皋黄鸡开产日龄遗传力估值除与东亚地方鸡种的近似外,低于其他研究估值。其原因除了本研究考虑了母体效应对遗传力影响外,遗传背景、统计方法及模型等因素也可能会造成遗传力估值上的不同。最近,全基因组关联研究解析出的原因性变异较少,预示以往开产日龄遗传力可能被过高估算[21- 22]。
对于如皋黄鸡,开产蛋质量遗传力为0.23左右。本研究结果略高于Wolc等[23]、Sang等[19]、Hosseini 等[24]以及Niknafs等[25]的研究结果,略低于吴桂琴等[26]的研究结果。与产蛋高峰期相比,开产阶段蛋质量遗传力较低,随着周龄增加,蛋质量遗传力逐渐升高,并趋于稳定。产蛋初期,卵巢、下丘脑、垂体以及能量供给器官仍以性成熟为主题发挥自身功能,在30周龄左右,产蛋鸡完成性成熟过程,一些与产蛋相关基因开始表达,加性遗传力逐步增加[27]。
生长性状遗传力一般高于生殖性状遗传力。如皋黄鸡开产体质量遗传力为0.34±0.03,高于开产日龄和开产蛋质量遗传力。如皋黄鸡开产体质量遗传力低于伊朗地方品种Far鸡(h2=0.57)、Mazandaran鸡(h2=0.48),但3个亚洲地方品种均检测到母体遗传的影响[20, 28]。如皋黄鸡开产体质量遗传力低于峪口褐壳蛋鸡父系(h2=0.60)[26],高于新杨白壳蛋鸡(h2=0.20)[29],与韩国地方鸡种的相近(h2=0.39)[19]。
3.2动物模型比较
以模型I作为子模时,似然比值与其他3个模型存在极显著差异,表明模型I不适于如皋黄鸡开产性状遗传力估计。对于开产日龄,模型III和IV之间存在显著差异,表明母体环境效应有着显著影响;模型II与III之间为负值,如果调换子模位置,则模型之间存在显著差异,支持母体环境效应影响下显著。两者综合,可选择模型II为开产日龄适用动物模型。
对于开产蛋质量,除去模型I之外,模型II与模型III、IV之间存在显著差异,表明模型II并非适用模型;模型III、IV之间似然比值差异不显著,残差比值以及母体遗传效应相差不大,参数少的模型III是开产蛋质量适用模型。
对于开产体质量,模型II与III互换子模位置,则2个模型之间存在显著差异,模型II好于模型III;除模型I之外,3种模型两两间存在显著差异,所以选择模型IV为最适模型。
对肉蛋兼用型品种如皋黄鸡来说,开产性状选育具有重要经济意义。研究结果表明如皋黄鸡开产性状属于低遗传力性状,受环境因素影响较大,因而应用传统表型选择很难获得良好遗传进展。母体效应影响如皋黄鸡开产性状,计算育种值、选择反应时应考虑母体效应对遗传力的影响。如皋黄鸡开产日龄、开产蛋质量、开产体质量遗传力分别为0.28、0.23、0.34。
参考文献:
[1]MOUSSEAU T A, FOX C W. The adaptive significance of maternal effects[J]. Trends Ecol Evol, 1998, 13(10): 403-407.
[2]BIARD C, GIL D, KARADAF, et al. Maternal effects mediated by antioxidants and the evolution of carotenoid-based signals in birds[J]. Am Nat, 2009, 174(5): 696-708.
[3]NEWBREY J L, REED W L. Growth of yellow-headed blackbirdXanthocephalusxanthocephalusnestlings in relation to maternal body condition, egg mass, and yolk carotenoids concentrations[J]. J Avian Biol, 2009, 40(4): 419-429.
[4]TUISKULA-HAAVISTO M, VILKKI J. Parent-of-origin specific QTL--a possibility towards understanding reciprocal effects in chicken and the origin of imprinting[J]. Cytogenet Genome Res, 2007, 117(1/2/3/4): 305-312.
[5]MCELROY J P, KIM J J, HARRY D E, et al. Identification of trait loci affecting white meat percentage and other growth and carcass traits in commercial broiler chickens[J]. Poult Sci, 2006, 85(4): 593-605.
[6]HARTMANN C, JOHANSSON K, STRANDBERG E, et al. Genetic correlations between the maternal genetic effect on chick weight and the direct genetic effects on egg composition traits in a White Leghorn line[J]. Poult Sci, 2003, 82(1): 1-8.
[7]MANIATIS G, DEMIRIS N, KRANIS A, et al. Model comparison and estimation of genetic parameters for body weight in commercial broilers[J]. Can J Anim Sci, 2013, 93(1): 67-77.
[8]ASLAM M L, BASTIAANSEN J W M, CROOIJMANS R P M A, et al. Genetic variances, heritabilities and maternal effects on body weight, breast meat yield, meat quality traits and the shape of the growth curve in turkey birds[J]. BMC Genet, 2011,[2015-06-06].http://www.biomedcentral.com/1471-2156/12/14. doi:10.1186/1471-2156-12-14.
[9]SZWACZKOWSKI T, GRZECH M, BOROWSKA A, et al. Maternal genetic effects on body weight and breast morphological traits in duck population under selection[J]. Arch Tierz-Arch Anim Breed, 2010, 53(5): 600-608.
[10]SZWACZKOWSKI T, WEZYK S, STANISLAWSKA-BARCZAK E, et al. Genetic variability of body weight in two goose strains under long-term selection[J]. J Appl Genet, 2007, 48(3): 253-260.
[11]LOTFI E, ZEREHDARAN S, AZARI M A. Direct and maternal genetic effects of body weight traits in Japanese quail (Coturnixcoturnixjaponica)[J]. Arch Geflugelkd, 2012, 76(3): 150-154.
[12]MEYER K. WOMBAT:A tool for mixed model analyses in quantitative genetics by restricted maximum likelihood (REML)[J]. J Zhejiang Univ Sci B, 2007, 8(11): 815-821.
[13]白俊艳,李金泉,贾小平,等. 母体遗传效应对绒山羊生产性状遗传参数估计的影响[J]. 遗传, 2006, 28(9): 1083-1086.
[14]MEYER K. Likelihood calculations to evaluate experimental designs to estimate genetic variances[J]. Heredity, 2008, 101(3): 212-221.
[15]POGGENPOEL D G, DUCKITT J S. Genetic basis of the increase in egg weight with pullet age in a White Leghorn flock[J]. Brit Poultry Sci, 1988, 29(4): 863-867.
[16]KOERHUIS A N M, MCKAY J C. Restricted maximum likelihood estimation of genetic parameters for egg production traits in relation to juvenile body weight in broiler chickens[J]. Livest Prod Sci, 1996, 46(2): 117-127.
[17]JEYARUBAN M, GIBSON J. Estimation of additive genetic variance in commercial layer poultry and simulated populations under selection[J]. Theor Appl Genet, 1996, 92(3/4): 483-491.
[18]张龙,刘益平,赵小玲,等. 二郎山山地鸡两品系主要繁殖性状遗传参数的估计及分析[J]. 中国家禽, 2012, 34(14): 69-71.
[19]SANG B D, KONG H S, KIM H K, et al. Estimation of genetic parameters for economic traits in Korean native chickens[J]. Asian Australas J Anim Sci, 2006, 19(3): 319-323.
[20]GHORBANI S, KAMALI M A, ABBASI M A, et al. Estimation of maternal effects on some economic traits of north iranian native fowls using different models[J]. J Agric Sci Technol, 2012, 14(1): 95-103.
[21]SHEN X, ZENG H, XIE L, et al. The GTPase activating Rap/RanGAP domain-like 1 gene is associated with chicken reproductive traits[J]. PLoS One, 2012, 7(4): e33851.
[22]LIU W, LI D, LIU J, et al. A genome-wide SNP scan reveals novel loci for egg production and quality traits in white leghorn and brown-egg dwarf layers[J]. PLoS One, 2011, 6(12): e28600.
[23]WOLC A, ARANGO J, SETTAR P, et al. Genetic parameters of egg defects and egg quality in layer chickens[J]. Poult Sci, 2012, 91(6): 1292-1298.
[24]HOSSEINI S, TAHMOORESPUR M. Estimation of genetic parameters of various economic traits in west Iranian native chickens[J]. Indian J Anim Res, 2013, 47(3): 205-211.
[25]NIKNAFS S, NEJATI-JAVAREMI A, MEHRABANI-YEGANEH H, et al. Estimation of genetic parameters for body weight and egg production traits in Mazandaran native chicken[J]. Trop Anim Health Prod, 2012, 44(7): 1437-1443.
[26]吴桂琴, 李花妮, 刘会英, 等. 峪口褐壳纯系蛋鸡父系主要性状的遗传参数分析[J]. 中国家禽, 2015, 37(9): 49-51.
[27]WOLC A, SZWACZKOWSKI T. Estimation of genetic parameters for monthly egg production in laying hens based on random regression models[J]. J Appl Genet, 2009, 50(1): 41-46.
[28]GHORBANI S, TAHMOORESPUR M, MAGHSOUDI A, et al. Estimates of (co)variance components for production and reproduction traits with different models in Fars native fowls[J]. Livest Sci, 2013, 151(2/3): 115-123.
[29]田春雷, 严华祥, 杨长锁, 等. 新杨白壳纯系蛋鸡主要性状的遗传参数估计[J]. 上海畜牧兽医通讯, 2009(5): 9-11.
【责任编辑柴焰】
Direct and maternal genetic effects for traits at the first egg in Rugao yellow chickens
GUO Jun, QU Liang, SHEN Manman, DOU Taocun, HU Yuping, WANG Kehua
(Institute of Poultry Science, Chinese Academy of Agricultural Science, Yangzhou 225125, China)
Abstract:【Objective】 The aims of this study were to evaluate direct and maternal genetic effects for traits at the first egg of Rugao yellow chickens with animal models, and to estimate the genetic parameters for these traits. 【Method】Reproduction data, including data for 10 454 individual animals and 31 362 reproduction records, of Rugao yellow chickens from 2009 to 2012 were collected. Four animal models with and without maternal effects were used to estimate the genetic parameters. Phenotypic variance of reproduction traits was partitioned using average information restricted maximum likelihood (AIREML) method with WOMBAT software. The models were evaluated with the likelihood ratio test. Univariate models were used to estimate heritabilities, and multivariate models were used to analyze genetic and phenotypic correlations.【Result】Maternal environment had significant influence on age at the first egg (AFE), for which the optimum model was model II and the heritability was 0.28±0.02. Maternal genetic effects had significant influence on egg mass at the first egg (EM1), for which the optimum model was model III, the direct heritability was 0.23±0.02 and the maternal genetic heritability was 0.03±0.01. Both maternal environment and maternal genetic effects had significant influence on body mass at the first egg (BM1), for which the optimum model was model IV, the heritability was 0.34±0.03 and the maternal genetic heritability was 0.04±0.01. There were negative correlations between the direct and maternal genetic effects for EM1 and BM1. The genetic correlation coefficients between AFE and BM1, AFE and EM1, and EM1 and BM1 were 0.36±0.07, 0.33±0.07, and 0.65±0.05, respectively.【Conclusion】The traits at the first egg of Rugao yellow chickens are influenced by maternal effects, and are lowly heritable.
Key words:Rugao yellow chicken; heritability; reproduction trait; maternal effect; animal model
中图分类号:S831
文献标志码:A
文章编号:1001- 411X(2016)02- 0001- 06
基金项目:现代农业产业技术体系建设专项资金(CARS-41-K02);江苏省自然科学基金(BK20131237);江苏省科技成果转化专项资金(BA2014132)
作者简介:郭军(1971—),男,副研究员,博士,E-mail:guojun.yz@gmail.com;通信作者:王克华(1962—),男,研究员,博士,E-mail:sqbreeding@126.com
收稿日期:2015- 06- 27优先出版时间:2016- 01- 18
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20160118.1657.034.html
郭军, 曲亮, 沈曼曼,等.如皋黄鸡开产性状加性遗传及母体遗传效应分析[J].华南农业大学学报,2016,37(2):1- 6.
