苯系物联合暴露仿刺参管足转录组差异表达基因分析
2016-03-17潘泳嘉周遵春王摆高杉孙红娟关晓燕董颖闫喜武
潘泳嘉,周遵春,王摆,高杉,孙红娟,关晓燕,董颖,闫喜武
1. 大连海洋大学,大连 116023 2. 辽宁省海洋水产科学研究院,大连 116023
苯系物联合暴露仿刺参管足转录组差异表达基因分析
潘泳嘉1,2,周遵春2,*,王摆2,#,高杉2,孙红娟2,关晓燕2,董颖2,闫喜武1
1. 大连海洋大学,大连 116023 2. 辽宁省海洋水产科学研究院,大连 116023
为了分析苯系物(BTEXs)联合暴露后仿刺参(Apostichopus japonicus)管足转录组差异表达基因,采用Illumina HiSeqTM2000测序技术,对1.0 mg·L-1苯系物(B)联合暴露12 h后和对照组(C)的仿刺参管足组织分别进行转录组测序。经Trinity软件进行de novo组装,获得了145 675条unigenes。利用公共数据库进行同源比对,共注释了35 330条unigenes。对比分析苯系物联合暴露组和对照组的转录组测序结果,获得了2 418个差异表达基因(DEGs) (|Log2Fold changes| ≥ 1且FDR ≤ 0.001),其中,上调表达和下调表达基因分别为1 049和1 369个。GOseq分析结果显示,158个DEGs显著富集在149个GO terms中,包括103个生物学过程、17个细胞组分和29个分子功能(P < 0.05);KEGG代谢通路分析结果显示994个差异基因映射到268条代谢通路,这些差异表达基因参与的生理过程与其他生物的同源基因参与的信号传导、癌症、外源性化合物的生物降解代谢等过程相类似。上述结果为在转录组水平筛选苯系物的生物标志物,解析苯系物对仿刺参毒性作用的分子机制提供了科学参考。
苯系物;仿刺参;管足;差异表达基因;转录组测序
苯系物(BTEXs)包括苯(Benzene)、甲苯(Toluene)、乙基苯(Ethylbenzene)和二甲苯(Xylene)[1],具有较强的生物毒性作用。苯系物中的苯是致癌物,可导致白血病和淋巴瘤[2],引起染色体断裂和阻碍染色体分离[3];甲苯具有神经毒性和致畸作用[4-5];乙基苯可以诱导细胞色素P450(Cytochrome P450, CYP)和细胞色素-c-还原酶系统,也是线粒体解偶联剂,长期的乙基苯暴露,则会造成呼吸系统和肾脏损伤[6-7];二甲苯对人的神经系统、肝脏和肾脏具有损伤作用[8];此外,苯系物作为全球生产和使用最为广泛的50种有机化合物之一,也是水环境中优先控制污染物之一。苯系物随着海洋溢油、生活污水和石化废水进入近岸海洋环境,表层海水中苯系物因挥发、光解、微生物的降解,通常维系在ng·L-1,港口表层海水苯系物含量可以达到μg·L-1,海水中部分苯系物随着中颗粒物质沉降,在沉积物中累积,可以达到μg·kg-1,甚至达到mg·kg-1[9-10],对近岸海洋底栖生态环境安全构成潜在危害。
仿刺参(A. japonicus)属于棘皮动物门,仿刺参属,是我国北方海域重要的底栖生物类群,通过管足行动并感知环境变化[11-12]。前期研究发现仿刺参对苯系物暴露较为敏感,低剂量苯系物暴露导致仿刺参的脂质过氧化损伤及相关抗氧化基因和酶活性的显著变化[13-15]。为进一步了解苯系物对仿刺参基因组转录水平的影响,本研究利用Illumina HiSeqTM2000测序技术,通过空白对照组和苯系物联合暴露组的仿刺参管足组织的转录组测序分析,筛选出差异表达基因,并进行差异表达基因的GO富集和KEGG通路分析,为在转录组水平上筛选苯系物的生物标志物,解析苯系物对仿刺参毒性作用的分子机制提供参考。
1 材料与方法(Materials and methods)
1.1 实验材料与试剂
仿刺参(A. japonicus)为辽宁省海洋水产科学研究院引育种中心培育的幼参,体重(10±2) g,经清洁海水驯养2周后,选取健康幼参用于后续试验。
苯、甲苯、乙基苯、二甲苯和二甲基亚砜(DMSO)均为分析纯(中国国药有限公司)。
1.2 苯系物联合毒性实验
根据苯系物对仿刺参96 h-LC50,计算出苯、甲苯、乙基苯、二甲苯对仿刺参急性毒性的比值为4:2:2:3。设置苯系物联合处理浓度1.0 mg·L-1(其中,苯0.36 mg·L-1,甲苯0.18 mg·L-1,乙基苯0.18 mg·L-1,二甲苯0.28 mg·L-1),同时设置空白对照组,各处理组设置2个平行。在玻璃缸中配制15 L实验溶液,每个缸中加10只幼参,试验条件:水温15 ℃,盐度30,pH 8.0,间断性充氧,确保溶解氧大于4.5 mg·L-1,避光。
1.3 样品采集、RNA提取
苯系物处理12 h后,分别取空白对照组C_1、C_2和苯系物联合处理组B_1、B_2的仿刺参9只,冰上采集管足样品,放于液氮中速冻,-80 ℃保存。Trizol法提取上述样品的总RNA,DNase I进行DNA消化处理,电泳检测总RNA的质量和纯度,Agilent 2100检测RNA完整性。
1.4 转录组测序
将检测合格的RNA样品,用带有Oligo(dT)的磁珠进行mRNA的富集与纯化,将mRNA打断成短片段,以mRNA为模板,进行测序文库构建和Illumina HiSeqTM2000测序,获得空白对照组C_1、C_2和苯系物联合暴露组B_1、B_2测序数据。
1.5 测序数据的拼接和组装
测序得到的原始reads,去除带接头的reads,N(无法确定碱基信息)比例大于10%的不确定reads和低质量reads,得到clean reads,采用Trinity软件(trinityseqrnaseq_r2013_08_04)对clean reads进行de novo混合拼接。过滤和组装以后得到高质量的unigenes。
1.6 Unigenes功能注释
利用Transdecoder软件对unigenes序列中编码区进行翻译,在翻译过程中调用Rfam数据库。将翻译后的氨基酸序列分别与NCBI蛋白数据库(NCBI non-redundant protein sequence, Nr)、注释的蛋白质序列数据库(SWISS-PROT)、蛋白质直系同源簇数据库(Cluster of Orthologous Groups, COG)、基因本体论(Gene Ontology, GO)数据库、KEGG直系同源(Kyoto Encyclopedia of Genes and Genomes Orthology, KO)数据库做BLASTP序列相似性比对,并使用Blast2GO[16]对Nr注释的结果文件进行GO term映射,获得GO注释。
1.7 差异表达基因的筛选及分析
利用RSEM软件(v1.2.6)进行基因表达定量分析,采用FPKM(fragments per kb million reads)进行表达量计算[17],使用DESeq算法进行差异表达分析[18],以P ≤ 0.05、假阳性率(false discovery rate, FDR) ≤ 0.001和|Log2Ratio| ≥ 1为标准,筛选苯系物联合暴露后仿刺参管足的差异表达基因。
采用GOSeq软件对差异表达基因进行GO富集分析[19],并对富集结果进行GO level 3水平上的DEG数量统计,获得GO level 3水平上差异表达基因的数量。根据unigenes的KO注释结果,统计差异表达基因在各个通路上的分布数量,利用超几何分布计算方式,以P < 0.05作为阀值,确定富集通路并统计差异表达基因的数量。
2 结果(Results)
2.1 仿刺参管足转录组的测序与组装
仿刺参管足4个转录组测序的总产出为217 144 662
条raw reads,去除低质量和含有接头的reads以后,得到212 804 626条clean reads(见表1),(G+C)%含量平均值为41.89%,碱基Q20均值为97.585%,说明转录组测序质量和数据量相对较高。
利用Trinity软件对clean reads进行组装(表2,图1),通过序列之间的overlap信息组装得到238 603重叠群(Contig),平均长度为827 bp,N50为1 493 bp,其中,长度在200 ~ 500 bp的Contig有155 653个,占总体的56.85%;500~1 000 bp的Contig有46 430个,占总体的19.45%;长度大于1 000 nt的Contig有56 520条,占全部Contig的23.68%。从中挑选145 675个转录本作为单基因簇(unigene),占总体的60.80%。
2.2 仿刺参管足unigenes功能注释
通过BLASP将unigenes序列比对到Nr、Swiss-PROT、COG、GO、KO数据库(E-value<1e-5),得到与unigenes具有高度序列相似性的蛋白,从而得到该unigenes的蛋白功能注释信息。其中匹配到Nr数据库中的有18 361条,占全部获得注释unigenes的51.97% (表3)。

图1 仿刺参管足序列长度分布图Fig. 1 Length distribution of sequences of the A. japonicus tube feet

表1 仿刺参管足Illumina HiSeqTM 2000测序产出质量统计Table 1 Data quality obtained by Illumina HiSeqTM 2000 sequencing of the A. japonicus tube feet
注:C_1、C_2为对照组,B_1、B_2为苯系物联合处理组,处理时间为12 h,苯系物处理浓度为1.0 mg·L-1。
Note: C_1, C_2 are control groups, B_1, B_2 are BTEXs treatment groups (exposed to 1.0 mg·L-1BTEXs for 12 h).

表2 仿刺参管足转录组测序组装质量统计Table 2 Assembly quality of A. japonicus tube feet transcriptome
unigenes注释到Nr数据库中的E值分布图显示,E值小于1E-100的unigene有18.03%,E值介于1E-60到1E-100之间的unigenes有17.63%,E值介于1E-5到1E-60之间的unigenes有64.34%(图2A)。相似度分布图显示,序列比对相似度在40%以上的unigenes占72.09%,相似度高于60%的unigenes占26.33%(图2B)。功能注释的同源序列的物种分布情况中见图2C,注释到紫海胆(Strongylocentrotus purpuratus)的序列有67.86%,注释到柱头虫(Saccoglossus kowalevskii)的序列有2.37%。
将仿刺参管足转录组获得的unigenes与COG数据库进行比对,对其做了功能分类和统计,注释到COG的8 409个unigenes分布于24个功能分类(图3),如:翻译,核糖体结构和生物合成,翻译后修饰,折叠和分子伴侣类基因,核酸复制、重组和修复,氨基酸、脂肪和糖的转运和代谢等。

表3 unigenes在Nr、SWISS-PROT、COG、GO、KO数据库中的注释Table 3 Annotation of unigenes in Nr, SWISS-PROT,COG, GO and KO databases
注:Nr、SWISS-PROT、COG、GO、KO分别表示NCBI蛋白数据库、注释的蛋白质序列数据库、蛋白质直系同源簇数据库、基因本体论数据库、KEGG直系同源数据库;Percentage表示在该数据库中得到匹配蛋白的unigenes占全部获得注释的unigenes的百分比。
Note: Nr, SWISS-PROT, COG, GO, KO stand for NCBI non-redundant protein sequences Database, Swiss-Prot Protein Sequence Database, Cluster of Orthologous Groups Database, Gene Ontology Database, Kyoto Encyclopedia of Genes and Genomes Orthology Database, respectively; Percentage means the percentage of unigenes matched in the database in all the annotated unigenes.
对仿刺参管足转录组unigenes进行GO分析发现,有10 532个unigenes注释到GO数据库,有6 722、8 018、8 503个unigenes被归属到细胞组分、分子功能和生物学过程中(图4),可细分为51个亚类,如生物学过程中,细胞过程和代谢过程所占比例较高,细胞和细胞器部分在细胞组分所占比例较高,连接和催化活性在分子功能中占有较高比例。
仿刺参管足转录组unigenes的KO注释结果显示(图5),11 756个unigenes获得9 873个同源蛋白注释,其中7 305个unigenes得到的5 422个同源蛋白参与344个已知代谢通路。代谢通路分为人类疾病、组织系统、基础代谢、环境信息进程、细胞进程、遗传信息进程等6大类。
2.3 苯系物联合暴露仿刺参管足差异表达基因筛选
采用DESeq算法计算unigene在对照组和苯系物联合暴露实验组中基因表达量进行差异分析,以|Log2Fold changes| ≥ 1、P < 0.05、FDR ≤ 0.001为筛选标准,共得到2 418个差异表达基因,其中,上调基因有1 049个,下调基因有1 369个。图6为差异表达基因的火山图。
通过差异表达基因的相关性分析发现,空白对照组C_1和C_2相关系数为0.95,苯系物联合暴露实验组B_1和B_2的相关系数为0.96,而对照组和苯系物联合暴露实验组的相关性为-0.98(图7),说明平行样测序分析获得的差异表达基因高度相似,苯系物暴露组与对照组的差异显著。
2.4 差异表达基因的GO富集分析
GOseq分析结果显示,158个DEGs显著富集在149个GO terms中,包括103个生物学过程,17个细胞组分和29个分子功能。将富集的GO terms归类到level GO 3分类中,并统计词条中DEGs的数量(图8)。
2.5 差异表达基因的KEEG富集分析
差异表达基因的KEEG富集分析结果显示(图9):994个差异基因分布在268个通路中,涉及相关人类疾病、生物系统、基础代谢、环境信息进程、细胞进程、遗传信息进程的差异基因数量分别为284、186、183、141、111和89个。差异基因集中在以下通路较多:PI3K/Akt信号通路(ko04151)、吞噬(ko04144)、MAPK信号通路(ko04010)、Rap1信号通路(ko04015)、剪接体(ko03040)、溶酶体(ko04142)等。
统计通路内基因分布情况,使用超几何分布法进行统计,P < 0.05阀值筛选得到6个次级富集通路,分别为癌症(cancers)、感染性疾病(infectious diseases)、细胞生长与死亡(cell growth and death)、转录(transcription)、复制与修复(replication and repair)、糖的生物合成与代谢(glycan biosynthesis and metabolism)等(图10)。

图2 unigene与Nr数据库比对结果统计Fig. 2 Comparison results of unigene to Nr databases

图3 仿刺参管足unigenes的COG功能分类Fig. 3 COG function classification of unigenes from A. japonicus tube feet

图4 Unigenes的GO分类Fig. 4 GO functional categories of unigenes

图5 unigenes的KO功能分类Fig. 5 KO functional categories of unigenes

图6 差异表达基因的火山图Fig. 6 Volcano map of differential expression genes (DEGs)
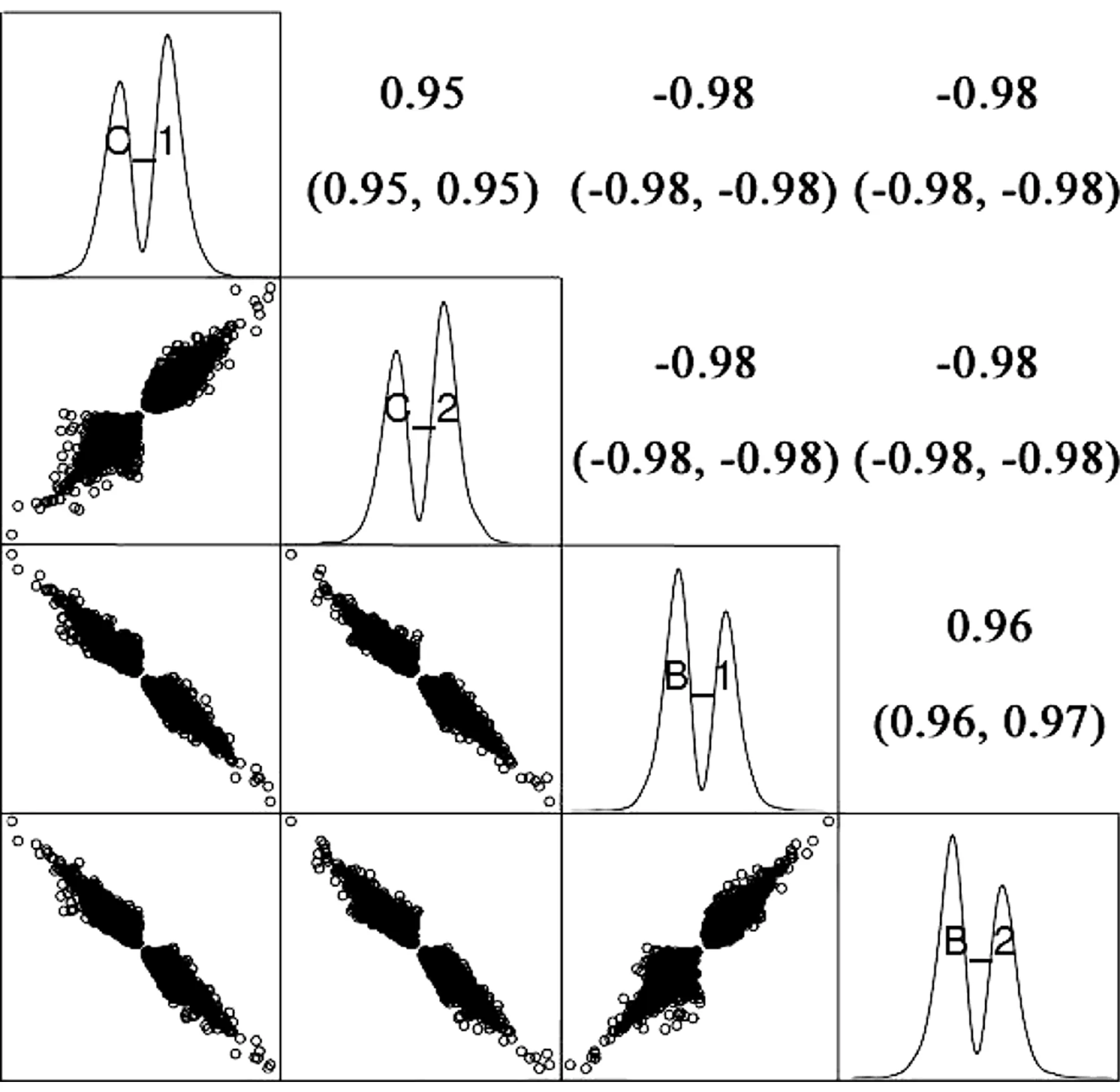
图7 对照组和苯系物处理组样本的相关性分析Fig. 7 Correlation between control and BTEXs treatment samples

图8 GO Term富集的DEGs数量统计Fig. 8 The enrichment GO Term of DEGs

图9 差异基因在通路中的分布情况Fig. 9 The number of differential expression genes in pathway

图10 pathway通路富集图Fig. 10 Statistics of pathway enrichment
3 讨论(Discussion)
转录组测序技术的广泛应用,为研究环境污染物对海洋生物基因组转录水平的影响,分析其分子作用机制提供了重要的技术手段。利用转录组测序技术,Hook等[20]分析了氨、铜、原油和除草剂西玛津对海洋硅藻(Ceratoneis closterium)转录组的影响;Huang等[21]报道了全氟辛烷磺酰基化合物(PFOS)对海水青鳉(Oryzias melastigma)转录组的影响;Ron等[22]研究了重金属汞、铜、锌、镉对海葵(Nematostella vectensis)转录组的影响。目前,有关苯系物对仿刺参基因组转录水平影响的研究尚未见报道。本研究中转录组测序、组装及注释的结果与仿刺参相关的转录组研究结果较为相近[23-25]。注释的unigenes中有23 975条(67.86%)与紫海胆(S. purpuratus)的匹配,紫海胆(S. purpuratus)作为唯一完成全基因组测序的棘皮动物,具有丰富的基因组信息[26],为仿刺参管足转录组的unigenes注释提供了参考序列。
苯系物作为单环芳烃化合物,其正辛醇-水分配系数logKow值在2.13~3.20之间,具有亲脂性和一定的生物富集性。Roose和Brinkman[27]研究发现海洋鱼类鳕鱼(Merlangius merlangus)和比目鱼(Limanda limanda)肌肉、肝脏组织中的苯系物含量在ng·g-1,是环境中苯系物含量的100倍以上。苯系物进入生物体后,可以被CYP同工酶(如:CYP1A1)代谢为可溶性产物排出体外[5]。差异表达基因的KEEG富集分析发现,41.1%的差异表达基因富集到286个代谢通路。部分差异表达基因富集到外源化合物生物降解代谢通路:CYP参与的外源化合物代谢(ko00980)和药物代谢(ko00982)以及其他酶参与的药物代谢(ko00983)通路,包括CYP A46、谷胱甘肽硫转移酶(glutathione S-transferase, GST)、微粒体环氧化物酶(microsomal epoxide hydrolase, EPHX1)、羧酸酯酶(carboxylesterase 2, CES2)、黄嘌呤脱氢酶/氧化酶(xanthine dehydrogenase/oxidase, XDH)等。这表明苯系物进入仿刺参管足组织后,可能被CYP氧化代谢。由芳香烃受体(aryl hydrocarbon receptor, AhR)介导的芳烃化合物CYP氧化代谢是耗氧过程,在产生氧自由基的同时,造成细胞内缺氧,进而激活缺氧诱导因子(hypoxia-inducible factor 1, HIF-1)调控的缺氧应答机制[28-29]。KEEG富集分析发现苯系物暴露后,仿刺参管足差异表达的基因在HIF-1代谢通路(ko04066)富集,如:缺氧诱导因子脯氨酰羟化酶(hypoxia-inducible factor prolyl hydroxylase, HPH)、己糖激酶(hexokinase, HK)等。
苯系物联合暴露导致仿刺参管足细胞内大量与人类癌症相关基因同源性较高的基因差异表达(上调基因44个,下调基因70个),包括:肿瘤坏死因子联合受体3(TNF receptor-associated factor 3, TRAF3)、细胞粘合素(tenascin, TN)、蛋白水解酶caspase 9等。这些差异表达基因富集的代谢通路包括:ko05200、ko05202、ko05203、ko05230、ko05231等。TRAFs为TNF转接上游分子信号激活下游基因,在适应性和先天性免疫中发挥重要作用[30]。TRAF3基因缺失导致新生小鼠(Mus musculus)的死亡[31]。TRAF3存在于仿刺参的各种组织中,可能在抗菌的过程通过调节ROS产生发挥重要的作用[32]。
此外,差异表达基因富集较多的信号通路有磷脂酰肌醇3-激酶/Akt信号通路(PI3K/Akt)、信号通路(ko04151)、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)信号通路(ko04010)等。其中,PI3Ks蛋白家族参与细胞增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节,PI3K活性的增加常与多种癌症相关[33]。MAPK是一组能被不同的细胞外刺激(如细胞因子、神经递质、激素、细胞应激及细胞黏附等激活的丝氨酸-苏氨酸蛋白激酶)调节细胞的生长、分化、对环境的应激适应、炎症反应等多种重要的细胞生理/病理过程[34]。
通过对差异基因的GO富集分析,可以定位差异基因最可能相关的GO term,有助于了解差异表达基因的功能[35]。苯系物联合暴露后,GOseq分析发现,158个DEGs显著富集在149个GO terms中,包括103个生物学过程,17个细胞组分和29个分子功能。为深入研究这些差异表达基因的生物学功能和了解苯系物对仿刺参毒性作用的分子机制提供了参考。
[1] 李学峰, 周启星. BTEX的环境质量标准研究进展[J]. 生态学杂志, 2011, 30(2): 369-375
Li X F, Zhou Q X. Environmental quality standard of BTEX: A review [J]. Chinese Journal of Ecology, 2011, 30(2): 369-375 (in Chinese)
[2] Recio L, Bauer A, Faiola B. Use of genetically modified mouse models to assess pathways of benzene-induced bone marrow cytotoxicity and genotoxicity [J]. Chemico-Biological Interactions, 2005, 153-154(6): 159-164
[3] Bird M G, Greim H, Snyder R, et al. International symposium: Recent advances in benzene toxicity [J]. Chemico-Biological Interactions, 2005, 153-154(6): 1-5
[4] Chen C S, You C H, Liang S H, et al. Assessment of genotoxicity of methyl-tert-butyl ether, benzene, toluene, ethylbenzene, and xylene to human lymphocytes using comet assay [J]. Journal of Hazardous Materials, 2008, 153(1-2): 351-356
[5] Rosa J C F D, Fiegenbaum M, Soledar A L, et al. Cytogenetic evaluation and the association with polymorphisms of the CPY1A1 and NR1I3 genes in individuals exposed to BTEX [J]. Environmental Monitoring & Assessment, 2012, 185(7): 5883-5890
[6] Hammer K D. Metabolite ratio of toluene-exposed rotogravure printing plant workers reflects individual mutagenic risk by sister chromatid exchanges [J]. Mutation Research, 2002, 519(1-2): 171-177
[7] National Toxicology Program. NTP toxicology and carcinogenesis studies of ethylbenzene (CAS No. 100-41-4) in F344/N rats and B6C3F1 mice (inhalation studies) [R]. Public Health Service, National Institutes of Health, US Department of Health and Human Services, 1999
[8] US EPA. Toxicological review of xylenes [R]. Washington DC: US Environmental Protection Agency, 2003
[9] Cavalcante R M, de Andrade M V F, Marins R V, et al. Development of a headspace-gas chromatography (HS-GC-PID-FID) method for the determination of VOCs in environmental aqueous matrices: Optimization, verification and elimination of matrix effect and VOC distribution on the Fortaleza Coast, Brazil [J]. Microchemical Journal, 2010, 96(2): 337-343
[10] 朱四喜, 周唯, 杨红丽, 等. 浙江舟山潮间带沉积物中二甲苯的检测[J]. 广东化工, 2010, 37(2): 125-127
Zhu S X, Zhou W, Yang H L, et al. Determination of xylene in interdial sediments in Zhoushan, Zhejiang Province [J]. Guangdong Chemical Industry, 2010, 37(2): 125-127 (in Chinese)
[11] 张宝琳, 孙道元, 吴耀泉. 灵山岛浅海岩礁区刺参(Apostichopus japonicus)食性初步分析[J]. 海洋科学, 1995, 3: 11-13
Zhang B L, Sun D Y, Wu Y Q. Preliminary analysis on the feeding habit of Apostichopus japonicus in the rocky coast water off Lingshan Island [J]. Marine Science, 1995, 3: 11-13 (in Chinese)
[12] 刘晓威, 姜森颢, 周一兵, 等. 大连地区仿刺参养殖池塘底栖硅藻生产状况的周年变化研究[J]. 水产科学, 2013, 31(11): 679-682
Liu X W, Jiang S H, Zhou Y B, et al. Annual changes in production of benthic diatoms in sea cucumber Apostichopus japonicus culture ponds in Dalian [J]. Fisheries Science, 2013, 31(11): 679-682 (in Chinese)
[13] 姜北, 刘薇, 周遵春, 等. 苯系物对仿刺参体内脂质过氧化程度的影响[J]. 水产科学, 2014, 33(1): 15-21
Jiang B, Liu W, Zhou Z C, et al. Effects of benzene, toluene, ethylbenzene, and xylene (BTEX) on lipid peroxidation in sea cucumber Apostichopus japonicus [J]. Fisheries Science, 2014, 33(1): 15-21 (in Chinese)
[14] 高士博, 王摆, 董颖, 等. 苯系物对仿刺参catalase基因表达及酶活性的影响[J]. 生态毒理学报, 2015, 10(2): 297-305
Gao S B, Wang B, Dong Y, et al. The effects of BTEXs on the catalase gene expression and catalase activities in respiratory tree and intestine of sea cucumber Apostichopus japonicus [J]. Asian Journal of Ecotoxicology, 2015, 10(2): 297-305 (in Chinese)
[15] 董颖, 王摆, 崔程, 等. 苯系物对仿刺参肠、呼吸树谷胱甘肽过氧化物酶基因表达的影响[J]. 水产科学, 2015, 34(5): 311-315
Dong Y, Wang B, Cui C, et al. The effects of BTEXs on the glutathione peroxidase gene expression in intestine and respiratory tree of sea cucumber Apostichopus japonicus [J]. Fisheries Science, 2015, 34(5): 311-315 (in Chinese)
[16] Conesa A, Götz S, Garcíagómez J M, et al. Blast2GO:A universal tool for annotation, visualization and analysis in functional genomics research [J]. Bioinformatics, 2005, 21(18): 3674-3676
[17] Li B, Dewey C N. RSEM:Accurate transcript quantification from RNA-Seq data with or without a reference genome [J]. BMC Bioinformatics, 2011, 12(31): 93-99
[18] Anders S, Huber W. Differential expression of RNA-Seq data at the gene level- the DESeq package [R]. The European Molecular Biology Laboratory, 2013
[19] Young M D, Wakefield M J, Smyth G K, et al. Goseq: Gene Ontology testing for RNA-seq datasets [OL]. (2013-10-15) [2016-03-07]. http://159.226.251.229/videoplayer/goseq.pdf?ich_u_r_i=f68ba4c61b48099258830ed3d156c05e&ich_s_t_a_r_t=0&ich_e_n_d=0&ich_k_e_y=1645128906750963352452&ich_t_y_p_e=1&ich_d_i_s_k_i_d=4&ich_u_n_i_t=1
[20] Hook S E, Osborn H L, Gissi F, et al. RNA-Seq analysis of the toxicant-induced transcriptome of the marine diatom, Ceratoneis closterium [J]. Marine Genomics, 2014, 16(1): 45-53
[21] Huang Q, Dong S, Fang C, et al. Deep sequencing-based transcriptome profiling analysis of Oryzias melastigma exposed to PFOS [J]. Aquatic Toxicology, 2012, 120-121(3): 54-58
[22] Ron E, Maayan R, Roey K, et al. Early and late response of Nematostella vectensis transcriptome to heavy metals [J]. Molecular Ecology, 2014, 23(19): 4722-4736
[23] Ye Z, Yang H, Storey K B, et al. RNA-Seq dependent transcriptional analysis unveils gene expression profile in the intestine of sea cucumber Apostichopus japonicus during aestivation [J]. Comparative Biochemistry & Physiology Part D Genomics & Proteomics, 2014, 10(6): 30-43
[24] Zhou Z C, Dong Y, Sun H J, et al. Transcriptome sequencing of sea cucumber (Apostichopus japonicus) and the identification of gene-associated markers [J]. Molecular Ecology Resources, 2013, 14(1): 127-138
[25] Du H, Bao Z, Hou R, et al. Transcriptome sequencing and characterization for the sea cucumber Apostichopus japonicus (Selenka, 1867) [J]. Plos One, 2012, 7(3): e33311
[26] Sodergren E, Weinstock G M, Davidson E H, et al. The genome of the sea urchin Strongylocentrotus purpuratus [J]. Science, 2006, 314(5801): 941-952
[27] Roose P, Brinkman U A T. Determination of volatile organic compounds in marine biota [J]. Journal of Chromatography A, 1998, 799(s1-2): 233-248
[28] Fleming C R, Billiard S M, Di Giulio R T. Hypoxia inhibits induction of aryl hydrocarbon receptor activity in topminnow hepatocarcinoma cells in an ARNT-dependent manner [J]. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2009, 150(3): 383-389
[29] Vorrink S U, Severson P L, Kulak M V, et al. Hypoxia perturbs aryl hydrocarbon receptor signaling and CYP1A1 expression induced by PCB 126 in human skin and liver-derived cell lines [J]. Toxicology and Applied Pharmacology, 2014, 274(3): 408-416
[30] Chung J Y, Park Y C, Ye H, et al. All TRAFs are not created equal: Common and distinct molecular mechanisms of TRAF-mediated signal transduction [J]. Journal of Cell Science, 2002, 115(Pt 4): 679-688
[31] Xu Y, Cheng G, Baltimore D.Targeted disruption of TRAF3 leads to postnatal lethality and defective T-dependent immune responses [J]. Immunity, 1996, 5(5): 407-415
[32] Yang L, Chang Y, Wang Y, et al. Identification and functional characterization of TNF receptor associated factor 3 in the sea cucumber Apostichopus japonicus [J]. Developmental & Comparative Immunology, 2016, 59(6): 128-135
[33] Balachandran C, Emi N, Arun Y, et al. In vitro antiproliferative activity of 2, 3-dihydroxy-9, 10-anthraquinone induced apoptosis against COLO320 cells through cytochrome C release caspase mediated pathway with PI3K/AKT and COX-2 inhibition [J]. Chemico-Biological Interactions, 2016, 249(2): 23-35
[34] Cao C, Cui N, Wang P, et al. Sulfated polysaccharide isolated from the sea cucumber Apostichopus japonicus against PC12 hypoxia/reoxygenation injury by inhibition of the MAPK signaling pathway [J]. Cellular & Molecular Neurobiology, 2015, 35(8): 1-12
[35] Zhong S, Xie D. Geneontology analysis in multiple gene clusters under multiple hypothesis testing framework [J]. Artificial Intelligence in Medicine, 2007, 41(2): 105-115
◆
Analysis of Differentially Expressed Genes in the Tube Feet of Sea Cucumber (Apostichopusjaponicus) Co-exposed to BTEXs by Transcriptome Sequencing
Pan Yongjia1,2, Zhou Zunchun2,*, Wang Bai2,#, Gao Shan2, Sun Hongjuan2, Guan Xiaoyan2, Dong Ying2, Yan Xiwu1
1. Dalian Ocean University, Dalian 116023, China 2. Liaoning Ocean and Fisheries Science Research Institute, Dalian 116023, China
Received 07 March 2016 accepted 04 May 2016
In order to identify and analyze the differentially expressed genes (DEGs), transcriptome sequencing in the tube feet of sea cucumber Apostichopus japonicus exposed to 0.0 (C), 1.0 mg·L-1BTEXs (B) for 12 h, was performed respectively using the Illumina HiSeqTM2000 platform. The clean reads were then de novo assembled into 145 675 unigenes, and 35 330 unigenes were annotated by a similarity search against the public databases. By comparing B and C using the criteria |Log2Fold changes| ≥ 1 and false discovery rate (FDR) ≤ 0.001, 2 418 DEGs were identified, among which 1 049 were up-regulated and 1 369 were down-regulated. GOseq analysis revealed that for B vs. C, 158 DEGs were highly enriched in 149 GO terms, including 103 biological processes, 17 cellular components and 29 molecular function terms (P value < 0.05). Pathways associated with BTEXs challenge were also mined. The result indicated that 994 DEGs were enriched in 268 pathways, including signal transduction, cancer, xenobiotics biodegradation and metabolism pathways. The results obtained in this study could be used for the screening of BTEXs biomarkers on the transcriptome level, and provide the reference to understand the molecular mechanism of toxic effects of BTEXs on sea cucumber A. japonicus.
BTEXs; Apostichopus japonicus; tube feet; differentially expressed gene; transcriptome sequencing
海洋公益性行业科研专项(201205012-7);大连市科学技术基金(2012J21DW029)
潘泳嘉(1988-),男,硕士研究生,研究方向为动物遗传育种与繁殖,E-mail: 820092256@qq.com;
*通讯作者(Corresponding author), E-mail: zunchunz@hotmail.com
10.7524/AJE.1673-5897.20160307002
2016-03-07 录用日期:2016-05-04
1673-5897(2016)6-082-11
X171.5
A
周遵春(1967—),男,海洋生物学博士,研究员,主要研究方向海洋生物学,发表学术论文80余篇。
王摆(1981—),男,海洋生物学博士,副研究员,主要研究方向海洋生态毒理学,发表论文20余篇。
# 共同通讯作者(Co-corresponding author), E-mail:wangbai1980@hotmail.com
潘泳嘉, 周遵春, 王摆, 等. 苯系物联合暴露仿刺参管足转录组差异表达基因分析[J]. 生态毒理学报,2016, 11(6): 82-92
Pan Y J, Zhou Z C, Wang B, et al. Analysis of differentially expressed genes in the tube feet of sea cucumber (Apostichopus japonicus) co-exposed to BTEXs by transcriptome sequencing [J]. Asian Journal of Ecotoxicology, 2016, 11(6): 82-92 (in Chinese)
